 2017, Vol. 37
2017, Vol. 37文章信息
- 赵天启, 古琛, 王亚婷, 王玺, 贾丽欣, 杜宇凡, 陈万杰, 王舒新, 赵萌莉.
- ZHAO Tianqi, GU Chen, WANG Yating, WANG Xi, JIA Lixin, DU Yufan, CHEN Wanjie, WANG Shuxin, ZHAO Mengli.
- 不同利用方式下典型草原植物群落物种多度分布格局
- Species-abundance distribution of typical steppe grassland plant communities under different use patterns
- 生态学报. 2017, 37(23): 7894-7902
- Acta Ecologica Sinica. 2017, 37(23): 7894-7902
- http://dx.doi.org/10.5846/stxb201612212630
-
文章历史
- 收稿日期: 2016-12-21
- 网络出版日期: 2016-08-16

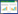




物种多度格局是群落结构的重要反映, 近年来受到生态学家的重视, 是当前生态学领域内的一个热点问题[1-2]。从定义上来讲它描述了群落内不同物种个体数量的分布情况[3-4]。不同群落具有不同的多度格局, 它是由同一群落的多个物种不断相互作用、相互影响的结果[5]。相比较传统的生物多样性指数, 物种多度格局能更好的体现出物种关系和作用机制[6]。根据物种的多度格局追溯物种的作用机制过程是揭示群落本质的重要方式[7]。对于这一重要的生态学概念, 研究者们开始从自然科学界的各个方面进行研究, 希望可以在理论基础上解释更多的生态学问题, 例如阔叶林多度格局的研究[8]、放牧强度和地形与物种多度分布的影响以及鸟类多度分布格局等。
从20世纪30年代开始, 数学模型结合生态学理论开始广泛的应用于物种多度分布的研究中[9-10]。于是Motomura[11]首先提出几何级数模型即生态位优先占领模型(niche preemption model, NPM)拟合物种多度分布曲线, 此后各种新的模型相继出现。MacArthur在1957年基于生态位分化理论提出了著名的断棍模型(broken stick model, BSM)也对一些数学模型进行了生态学解释[12], 与此同时, 他还提出了生态学研究中又一重要的模型:重叠生态位模型(overlapping niche model, ONM)。之后, 人们通过改变模型中的参数从而得到更多的生态位模型, 因而使其能够有更广的应用范围, 解释更多的生态学意义[13-14]。如Tokeshi[15]在1996年提出的随机分配模型(random assortment model, RAM)从数学理论上来说与K=0.5(NPM模型参数)时的NPM相一致, 但二者所述的生态学意义却是截然不同。还有类似优势优先模型(dominance preemption model, DPM)也是如此, 这些模型大多都应用在动物群落的研究中[16], 在植物群落尤其是在草原植物群落中的研究还比较少。近年来也有许多学者对各类物种多度模型进行了详细的介绍, 也对一些物种多度模型分布进行了实际分析应用。目前, 大多数的物种多度模型的拟合研究都集中于森林群落[17], 虽然也有少部分对于草地群落的研究, 但其主要是针对自然草地群落的研究[18], 而对人类活动干扰、动物行为干扰的草地生态系统的研究报道较少。
内蒙古典型草原是欧亚大陆草原的主要组成部分, 其物种组成和群落结构也是我国温带草原的典型代表, 在历史上一直被用作放牧场和割草场[19]。在草原生态系统中, 放牧和割草是两种主要的草地利用方式。其中放牧是草地生态系统中最普遍的利用方式[20], 家畜通过放牧行为会对草地生物多样性进行一定程度的影响[21]。割草是刈割方式的一种, 也是放牧的替代性行为, 适度刈割可以缓解放牧对草原生态系统的持续性压力[22]。本文以内蒙古典型草原为研究对象, 利用不同的数学模型对植物多度数据进行拟合研究, 旨在探求在不同利用方式下典型草原群落物种多度分布格局, 为研究典型草原植物群落多度格局提供思路和参考资料。
1 材料与方法 1.1 研究地区概况本实验是在内蒙古锡林浩特市东部约36km的毛登牧场(116.03°—116.50°E, 44.80°—44.82°N)进行的。该区域海拔约1055m, 属于温带干旱大陆性气候, 年平均气温0—1℃, 最热月在7月为21.2℃, 最冷月在1月为-19.4℃, ≥0℃的积温1800℃, 年平均降水量300—360mm, 无霜期90—115d, 具有光、热、水同期特点。土壤以栗钙土为主, 含少量褐色土, 软栗钙土层15—80cm, 土壤有机质含量2%—3%, 土质比较肥沃。地带性植被为大针茅草原。天然草地以大针茅(Stipa grandis)和羊草(Leymus chinensis)为优势种, 常见的物种还有糙隐子草(Cleistogenes squarrosa)、冰草(Agropyron cristatum)、冷蒿(Artemisia frigida)等。选择放牧和割草利用方式下的草地, 以围封草地作为对照, 各类样地在气候、地貌、土壤、原生群落类型相同。其中放牧样地全年放牧, 放牧利用超过50a, 依据2011—2016年移动围笼数据, 放牧利用率为55.6%。割草样地从2011年开始, 每年8月底进行1次刈割, 留茬高度5cm;围封样地自2008年起围栏封育。
1.2 实验设计与取样方法实验于2016年8月份, 分别在上述3类样地内选取植物生长均匀、地形相对一致的地段进行。每块样地内设3条50m的样线, 每条样线每隔0.5m设置一个50cm×50cm的大样方, 共100个, 称为L型样方, 再将每个L型样方均等地分成4个25cm×25cm的小样方, 称为S型样方。
8月中旬在生长季最旺盛的时候记录S型样方里出现的所有植物种。如果群落内某物种在L型样方内的4个S型样方内均未出现, 则记作0, 只在1个S型样方中出现就记作1, 如果在任意2个S型样方中出现记作2, 以此类推, 如果某物种在4个S型样方内都出现就记作4。记录小型样方里出现的所有植物种。
1.3 模型选取国内外在物种多度格局研究中运用的模型基本可分为3类:1)基于统计学的模型, 先提出假设, 然后运用已知数学理论, 经过计算算出的物种多度。这类模型的缺点是不能体现生物与生态学意义, 对赋予实际数据意义比较麻烦, 但它的优点对群落是有较好的拟合结果。2)生态位模型, 是为解决实际物种分布的生态学意义, 有些学者提出了能相对容易理解生态学过程的模型, 这类模型的优点是可以较好的把群落的物种多度与生态学过程结合在一起。3)中性理论为核心理论的其他模型。其目的与以上的模型相似, 都是用来解释物种多样性和生物学原理。这一理论的提出, 对研究森林多度和种-面积关系起了关键作用, 在生态学研究中也有着重要意义。
上述众多模型中, 经前人研究, 运用生态位模型与草地群落物种进行拟合的效果是最佳的, 它解释了草地生态系统中植被真实分布的生态学意义。因此本实验选取了其中5种较为普遍认可和应用较广泛的生态位模型进行典型草原物种多度格局的研究。
1.3.1 生态位优先占领模型生态位优先占领模型:又称几何级数模型, 该模型是假设第一个种先占据群落总资源(生态位)份额的k份, 第二个种占用剩余的k份, 即k(1-k), 第三种再占用据再剩余的k份, 即k(1-k)2, 以此类推。因此, 如果只考虑常见种, 则第i个种的理论多度所对应的比例为:

|
式中, k是待定参数。k的最佳取值应使真实多度与理论多度的相对偏差(d)最小。把k的初始值设定为0, 以0.001为步长逐步增加, 并利用Microsoft Excel求出与之对应的d值, 直到d越过最小值并显露增加趋势时停止迭代。取与d最小值对应的k作为最佳k值。
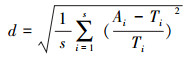
|
式中, Ai表示第i个物种的真实多度, Ti表示第i个物种的理论多度。S表示某群落的物种数, i表示第i个物种(下同)。
1.3.2 生态位重叠模型生态位重叠模型:是指把群落资源总量视为一根棒, 每个种的生态位等于棒上随机的两点间的距离, 各个种是彼此独立的。重叠生态位模型不是把棒分配给各个种, 而是每个种取其所需资源比例, 这样群落的总生态位或资源总量不再是1, 因为各个种之间有重叠。因此, 第i个物种的理论多度所对应的比例Pi为:
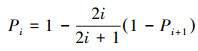
|
断棍模型:又称为分割线段模型、随机生态位假说。它是一种资源分配模型, 多度反映了资源在几个竞争种之间的随机分配是沿着一维梯度进行的。该模型假设一个群落中的总资源量是1的一根棒, 随机在棒上设置n-1个点, 把这根棒, 分为n段, 表示n个占有这个生态位。并且认为这n个物种分类地位接近, 竞争能力也相似, 而且在群落中出现时间相同。因此, 第i个物种的理论多度所对应的比例Pi为:

|
优势优先模型:该模型认为在群落中最优势的种类先占用总生态位的一半以上, 次优势种再占用剩余的一半以上, 以此类推。因此该模型实际上是几何级数模型的特殊形式, 即k>0.5, 该模型保证优势种类优先利用资源, 所有种在生态位占有上明显形成一个等级。Tokeshi认为该模型的k值应等于0.75。
1.3.5 随机分配模型随机分配模型:该模型假定一个群落中各个种的多度之间没有联系, 种间竞争不存在, 这有两种情况, 一是种的多度大小和生态位分配无关, 这可能在物种不饱和的群落中存在;二是由于环境不断变化, 生态位的分配是变化的, 非等级的。由于群落总生态位不断变化, 多数情况下, 物种不能全部占用其生态位, 这样每个种的生态位也总是在变化, 该模型认为物种对n个独立大小的生态位是随机的选取, 这些生态位若从大到小排列, 最大的生态位记为1, 次大的生态位为小于1的一个值, 第三大生态位是小于第二个生态位的一个值, 以此类推, 这一排列关系可以表示为:

|
Ni为第i个生态位的大小, ri为独立的随机变量介于(0, 1)之间, Ni的估计值为:
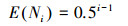
|
当由n个种组成的群落被考虑时, 第i个种的相对多度为:
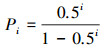
|
根据以上模型得到的所有种的Pi, 在同一群落中它们的和未必等于1。所以, 需要对Pi修正, 令修正以后的Pi等于修正以前的Pi除以所有种的Pi总和。也就是使修正后所有种的Pi的和等于1。用修正后的Pi乘以对应群落中所有物种的多度和, 即可算出各个物种的理论多度。5个模型对同一群落的理论多度各不相同, 用模型拟合的理论多度与真实多度相比较, 从而分析真实多度的分布特征。数据处理、模型拟合、统计检验和作图采用Excel 2013和R语言软件(R Development Core Team)[23]完成。
1.4.1 χ2检验χ2检验(Chi-square test, χ2)是对物种多度分布模型进行适合度检验的一种常用方法, χ2值越小, 模型的拟合效果就越好。总体检验时χ2大于1000、分功能群检验时χ2大于100时均未通过检验。
1.4.2 AIC准则和BIC准则赤池信息量准则(Akaike information criterion, AIC)由赤池弘次提出, 假设模型的误差服从正态分布, AIC可表示为:AIC=2k+nln(RSS/n)。式中, k表示模型中参数的数量, n表示观察数, RSS表示残差平方和。AIC值越小, 拟合效果更好。AIC准则也有不足之处:如果时间序列很长, 相关信息就越分散, 需要多自变量复杂拟合模型才能使拟合精度比较高。为了弥补AIC准则的不足, Akaike于1976年提出BIC准则。而Schwartz在1978年根据Bays理论也得出同样的判别准则, 称为BIC准则[24]。同样, BIC值越小, 模型拟合也越好。两种准则的模型检验都在R语言软件中完成。
2 结果与分析 2.1 不同利用方式下群落物种组成与变化不同利用方式下典型草原植物组成不同(表 1), 3个样地内共有35种植物分布。其中刈割区、放牧区和围封区的物种数分别为33种、16种和29种。相较于围封对照区, 刈割区的物种数增加了12%, 增加的物种有百里香和龙胆等, 而放牧区的物种数减少了44.8%, 减少的物种有冷蒿和乳白黄耆等。群落优势种大针茅、糙隐子草、知母、羊草和黄囊薹草在刈割区、放牧区及对照区的出现频度分别占整个群落物种总频度的88.33%、81.47%和75.6%。刈割区知母及羊草频度增加, 放牧区黄囊薹草、细叶韭的频度增加。
| 物种名称 Species |
频度Frequency/% | ||
| 刈割 Mowing |
放牧 Grazing |
对照 Enclose |
|
| 大针茅Stipa grandis | 28.67 | 32.48 | 31.05 |
| 糙隐子草Cleistogenes squarrosa | 21.03 | 10.1 | 22.75 |
| 知母Anemarrhena asphodeloides | 18.48 | 0.06 | 8.68 |
| 羊草Leymus chinesis | 15.1 | 8.15 | 12.62 |
| 黄囊薹草Carex korshinskyi | 5.05 | 30.68 | 0.5 |
| 刺藜Chenopodium aristatum | 2.35 | 1.08 | 1.05 |
| 冷蒿Artemisia frigida | 2.04 | 3.92 | |
| 猪毛菜Salsola collina | 1.65 | 0.41 | 2.3 |
| 黄花黄耆Astragalus luteolus | 0.84 | 0.03 | 1.35 |
| 银灰旋花Convolvulus ammannii | 0.8 | 0.14 | |
| 细叶韭Allium tenuissimum | 0.7 | 6.93 | 0.38 |
| 野韭Allium ramosum | 0.69 | 5.94 | 1.33 |
| 乳白黄耆Astragalus galactites | 0.59 | 1.4 | |
| 砂韭Allium bidentatum | 0.57 | 0.14 | |
| 灰绿藜Chenopodium glaucum | 0.37 | 1.31 | 5.33 |
| 小叶锦鸡儿Caragana microphylla | 0.17 | 2.15 | 0.17 |
| 点地梅Androsace umbellata | 0.11 | 0.03 | |
| 黄芩Scutellaria baicalensis | 0.11 | 0.02 | |
| 草麻黄Ephedra sinica | 0.1 | 0.12 | 0.1 |
| 唐松草Thalictrum aquilegifolium | 0.09 | 1.54 | |
| 百里香Thymus mongolicus | 0.09 | ||
| 大籽蒿Artemisia sieversiana | 0.09 | 1.21 | |
| 黄花葱Allium condensatum | 0.08 | 0.06 | 0.19 |
| 木地肤Kochia prostrata | 0.07 | 0.44 | 0.57 |
| 阿尔泰狗娃花Heteropappus altaicus | 0.04 | 0.02 | |
| 冰草Agropyron cristatum | 0.03 | 0.1 | |
| 藜Chenopodium album | 0.02 | 0.09 | 0.76 |
| 龙胆Gentiana scabra | 0.02 | ||
| 萹蓄Polygonum aviculare | 0.02 | ||
| 地锦Euphorbia humifusa | 0.01 | 0.02 | |
| 虫实Trigonotis corispermoides | 0.01 | 0.45 | |
| 小花花旗竿Dontostemon micranthus | 0.01 | ||
| 少花米口袋Gueldenstaedtia verna | 0.01 | ||
| 鹤虱Carpesium abrotanoides | 1.88 | ||
| 空白处表示相应的物种未出现 | |||
在放牧区的物种组成拟合中:卡方检测中放牧利用方式下χ2值为92.99, 在5中模型中值最低, (AIC=18.86、BIC=1.18)检验中表现最优的是随机分配模型(RAM), 但其值和生态位优先占领模型(NPM)中的相差甚微。NPM中AIC=174.52、BIC=176.86。
刈割区群落物种与模型的拟合结果是:生态位优先占领模型NPM的卡方值虽然最小(χ2=90.51), 但该模型下所有卡方值在全部处理中是最大(表 2)。AIC和BIC值也相对较小(AIC=321.32、BIC=325.81), 同样在全部处理中这两个值也最大;说明相较对照, 刈割对模型的拟合效果较差。
| 草地群落类型 Community type |
物种数 Number of species |
检测方法 Testing method |
模型拟合结果Model fitting results | ||||
| NPM | ONM | BSM | DPM | RAM | |||
| 放牧Grazing | 16 | 卡方分布χ2 | 92.99 | 422.69 | 268.98 | 579.34 | 129.14 |
| AIC | 174.52 | 182.02 | 170.1 | 187.14 | 168.86 | ||
| BIC | 176.84 | 184.33 | 172.42 | 189.46 | 171.18 | ||
| 对照Enclose | 29 | 卡方分布χ2 | 22.29 | 458.88 | 325.85 | 552.95 | 180.14 |
| AIC | 246.17 | 306.91 | 279.02 | 296.01 | 248.69 | ||
| BIC | 250.27 | 311.01 | 283.13 | 300.11 | 252.79 | ||
| 刈割Mowing | 33 | 卡方分布χ2 | 90.51 | 770.05 | 557.59 | 680.67 | 191.4 |
| AIC | 321.32 | 357.11 | 332.69 | 356.11 | 323.73 | ||
| BIC | 325.81 | 361.6 | 337.18 | 360.6 | 328.22 | ||
| BSM:断棍模型Broken stick model;NPM:生态位优先占领模型Niche preemption model;DPM:优势优先模型Dominance preemption model;RAM:随机分配模型Random assortment model;ONM:生态位重叠模型Overlapping niche model;AIC:赤池信息量准则Akaike information criterion;BIC:贝叶斯信息准则Bayesian Information Criterion | |||||||
对照区物种组成的拟合为:3种不同检验中的处理结果相同, 均符合生态位优先占领模型。χ2、AIC和BIC检测拟合度从优到差的排列结果均为:NPM>RAM>BSM>ONM>DPM。其中模型预测中的生态位优先占领模型(NPM)的检验结果最优(χ2=22.29、AIC=246.17、BIC=250.27), 这个模型中的卡方结果在两种利用方式及对照和所有模型拟合中的值也最低。
2.3 不同利用方式下群落物种多度格局将群落中的物种从常见到少见依次排序为横坐标(表 1), 各个物种相对应的多度为纵坐标, 比较5种生态位模型对3种不同利用方式的拟合效果(图 1)。其中:放牧利用方式下, 生态位优先占领模型(NPM)和随机分配模型(RAM)拟合效果相对较好(图 1);围封对照内, 生态位优先占领模型(NPM)的群落物种多度分布曲线与实际多度分布曲线几乎重叠, 拟合效果极好, 并且在所有利用方式下的草地群落物种多度预测模型中拟合中吻合度也最高(图 1);刈割利用方式下的观测值曲线初期和NPM、RAM均十分接近, 直至曲线后期, 观测值曲线与RAM的偏差越来越大, 而与NPM拟合的就相对吻合。3种利用方式中RAM和DPM均小于观测值曲线, BSM和ONM总体趋势都在观测曲线之上, 只有NPM的曲线一直与观测曲线拟合度最高, 且5种模型均通过检验。对整个典型草原物种群落进行具体物种分析, 在真实多度分布中, 建群种大针茅, 优势种糙隐子草、知母、羊草和黄囊薹草在不同利用方式下多度均较高, 而其余物种均表现出多度逐渐减少的特征。这5种植物的真实多度在与各个模型的前期均吻合。

|
| 图 1 不同利用方式下典型草原物种多度 Fig. 1 Species abundance of pattern under different use types in typical steppe BSM:断棍模型Broken stick model;NPM:生态位优先占领模型Niche preemption model;DPM:优势优先模型Dominance preemption model;RAM:随机分配模型Random assortment model;ONM:生态位重叠模型Overlapping niche model |
物种群落多度分布在使用不同的生态学模型拟合后, 能更好的反映出其生态学意义[2]。放牧和刈割两种不同利用方式下的物种多度分布最优模型分别是RAM、NPM和NPM(图 1, 表 2)。随机分配模型(RAM)是指假定一个群落中各个种的多度之间没有联系, 种间竞争不存在, 多数情况下物种不能全部占用其生态位[25]。这样的群落体系的一个最大的特点是没有足够的时间提供给基于资源竞争的内部关系去发展进化, 因此, 在整个生态位空间中, 各个物种的生态位不可能达到比较好的分配[8]。本实验中群落优势种大针茅、糙隐子草、知母、羊草和黄囊薹草在刈割区、放牧区及对照区的出现频度均占整个群落物种总频度的75%以上, 说明群落优势种未发生变化。但放牧利用下草地物种数量减少, 但黄囊薹草频度增加(表 1), 多度分布最优拟合模型为随机分配模型(RAM), 在放牧干扰下, 由于家畜的选择性采食和践踏的胁迫, 会使一部分物种, 如冷蒿、冰草等没有足够的时间去发展进化, 无法形成种间竞争, 进而从草群中消失, 有些地区出现裸地状况致使植物无法全部占用其生态位[26-27]。因此, 随机分配模型(RAM)就很好的拟合在放牧利用下的观测值曲线(图 1)。放牧利用下RAM和NPM中的结果相差不大(图 1, 表 2)。RAM模型曲线与观测值曲线在前中期拟合十分好, 但是后期是RAM和预测值曲线的拟合程度就比较NPM的拟合效果差。形成此现象的原因可能是放牧过程中, 家畜的采食、践踏和排泄等一系列活动使植物群落结构产生了变化[28]。除此之外, 群落中其他物种也可能受到其他不同形式的干扰, 例如降水和温度的变化, 使其在整个空间的生存竞争受到影响, 导致群落物种多度发生变化, 也致使各个模型曲线与真实曲线在物种序列后期发生了的偏差。施建敏等[29]研究的石灰岩山地淡竹(Phyllostachys glauca)林演替序列的群落物种多度分布格局中发现从淡竹林向阔叶林的进展演替过程中, 淡竹纯林、竹阔混交林和阔叶林的物种多度分布最优模型分别是DPM、NPM和NPM, 群落物种多度分布格局由DPM逐渐向NPM转变。在对物种多度分布的拟合结果中表明, 无论是平地还是坡地系统, 物种多度分布基本都符合基于生态位分化机制的幂分割模型, 这与Harpole和Tilman[30]的研究结果较为一致。张姗等[31]在长白山阔叶红松(Pinus koraiensis)林草本层物种多度分布格局及其季节动态的研究中认为:虽然不同季节的草本层物种多度分布有差异, 但模型的拟合结果相似, 表明不同季节的草本层可能具有相似的维持机制。
生态位优先占领模型(NPM)认为种的个体数量与它们所占有的资源数量成比例, 即优势度越高的物种应该占有的资源越多, 而且NPM的群落最优势种不占绝对优势, 其生态位小于总生态位的一半[12]。不同地区物种多度分布可能相同, 但造成这种相同格局的生态学过程可能不同。本研究中, 刈割和围封下物种多度分布最优拟合模型均为生态位优先占领模型(NPM), 但形成此最优模型的生态学过程完全不同。刈割利用会导致草原群落优势种植物的盖度降低, 减少优势种所占群落比例, 与此同时其他杂类草会获得繁殖的机会, 最后导致植物多样性和植物种类丰富度增加(表 1)。短期刈割处理没有改变群落优势种的优势地位, 群落优势种生态位占有率也未大于总生态位的一半, 这可能是因为刈割群落中的物种所占资源数量与其个体数量成比例的原因, 此现象符合生态位优先占领模型(NPM)是最优的生态学意义。但在围封对照中, 草地由于较长时间的不刈割、不放牧的处理, 积累了较厚的枯落物, 阻碍新种子与土壤接触, 导致新种子无法萌发, 进而致使一些植物种类不能及时更新繁衍, 最终导致物种数量下降[32]。但是围封可以积累足够的贮藏营养物质, 逐渐恢复草地生产力, 并促进草地群落自然更新;同时, 减少草地利用, 能使草地群落能更好的完成结籽或营养繁殖[33]。围封也没有使群落优势种占据绝对优势, 群落植物只按各自生长比例进行资源利用, 这同样符合生态位优先占领模型(NPM)的拟合结果。这说明在围封对照区植物群落物种分布更加均匀, 而放牧和刈割利用在一定程度上使群落优势种的生存优势进一步扩大, 从而使群落优势种得到整个群落中更大的生存空间。这是因为放牧区由于放牧的原因裸地的出现要多于其他两个区, 使其物种分布不是很均匀。许多研究现象与本研究的结果相一致, 如陈卓琳等[1]所研究的鸟类多度分布格局中表明:在不同的时间尺度和空间尺度干扰下, 北京西山静福寺地区鸟类群落多度格局模型与分割线段模型显著相关, 利用分割线段模型(断棍模型BSM)可以良好的拟合该地区鸟类群落多度格局。徐国瑞等[26]在海拔对表居土壤动物多度分布影响的研究中也体现出:拟合表居土壤动物群落整体的最优生态位分配模型不受海拔影响, 均为断棍模型(BSM)。但表居土壤动物各功能群多度分布受海拔影响, 杂食者随海拔升高, 最优模型由断棍模型转变为生态位优先占领模型(NPM), 而植食者与之相反。程佳佳等[34]在对亚热带常绿阔叶林群落物种多度分布格局对取样尺度的响应的实验中指出:中性理论模型除了边长为10m和20m尺度以外, 在其他尺度上均比其他5种模型的预测效果更好, 但是其他尺度中的最优拟合模型并不是中性理论模型。
本研究中用来研究典型草原物种群落多度格局的模型都属于生态位模型, 生态位模型也称之为资源分配模型, 运用这类模型可以把物种多度与生态位的占有或分配密切的联系起来。常用的生态位模型有11种[5], 本文选取了其中的5种。典型草原物种群落属于比较复杂的自然群落[33], 有的模型会对这些群落多度拟合较好, 同样也有一些模型的拟合效果较差。朱新玉等[18]研究的森林-草原交错带土壤节肢动物物种相对多度模型拟合研究种中指出:几何级数分布模型和分割线段分布模型则不适于描述该区土壤节肢动物群落物种多样性。所以, 在采用生态位模型对群落多度格局进行模拟时, 最好能够选用3种或3种以上模型, 并比较它们的结果, 然后确定最优模型[6]。模型的建立在很大程度上是数学过程, 其结果如不经过数理统计检验, 理论上难以被接受[35]。从本研究来看, 虽然在不同利用方式下的5种模型拟合结果都通过了检验, 但是检验结果却相差很大。不同利用方式下NPM和ONM模型间χ2值相差较大, 但AIC检验两种模型值的差异很小。因此, 仅用一个检验指标来检测多个多度模型就容易得到错误的结论, 只有综合多个指标才能做出正确的检验结果。
| [1] | 陈卓琳, 贾丽丽, 关文彬. 北京西山静福寺地区鸟类多度分布格局. 生态学报, 2017, 37(6): 1805–1815. |
| [2] | 刘梦雪, 刘佳佳, 杜晓光, 郑小刚. 亚高寒草甸不同生境植物群落物种多度分布格局的拟合. 生态学报, 2010, 30(24): 6935–6942. |
| [3] | MacArthur R. On the relative abundance of species. American Naturalist, 1960, 94(874): 25–36. DOI:10.1086/282106 |
| [4] | McGill B J, Etienne R S, Gray J S, Alonso D, Anderson M J, Benecha H K, Dornelas M, Enquist B J, Green J L, He F L, Hurlbert A H, Magurran A E, Marquet P A, Maurer B A, Ostling A, Soykan C U, Ugland K I, White E P. Species abundance distributions:moving beyond single prediction theories to integration within an ecological framework. Ecology Letters, 2007, 10(10): 995–1015. |
| [5] | 彭少麟, 殷祚云, 任海, 郭勤峰. 多物种集合的种-多度关系模型研究进展. 生态学报, 2003, 23(8): 1590–1605. |
| [6] | 马克明. 物种多度格局研究进展. 植物生态学报, 2003, 27(3): 412–426. |
| [7] | 牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579–593. |
| [8] | 张金屯. 美国纽约州阔叶林物种多度格局的研究. 植物生态学报, 1999, 23(6): 481–489. |
| [9] | Fisher R A, Corbet A S, Williams C B. The relation between the number of species and the number of individuals in a random sample of an animal population. The Journal of Animal Ecology, 1943, 12(1): 42–58. DOI:10.2307/1411 |
| [10] | Preston F W. The commonness, and rarity, of species. Ecology, 1948, 29(3): 254–283. DOI:10.2307/1930989 |
| [11] | Motomura I. On the statistical treatment of communities. Zool Magazine, 1932, 44: 379–383. |
| [12] | MacArthur R H. On the relative abundance of bird species. Proceedings of the National Academy of Sciences of the United States of America, 1957, 43(3): 293–295. |
| [13] | Cohen J E. Alternate derivations of a species-abundance relation. The American Naturalist, 1968, 102(924): 165–172. DOI:10.1086/282533 |
| [14] | Webb D J. The statistics of relative abundance and diversity. Journal of Theoretical Biology, 1974, 43(2): 277–291. DOI:10.1016/S0022-5193(74)80060-3 |
| [15] | Tokeshi M. Power fraction:a new explanation of relative abundance patterns in species-rich assemblages. Oikos, 1996, 75(3): 543–550. DOI:10.2307/3545898 |
| [16] | 刘文萍, 邓合黎. 木里蝶类多样性的研究. 生态学报, 1997, 17(3): 266–271. |
| [17] | 高利霞, 毕润成, 闫明. 山西霍山油松林的物种多度分布格局. 植物生态学报, 2011, 35(12): 1256–1270. |
| [18] | 朱新玉, 王玉玲, 高宝嘉, 胡云川. 森林-草原交错带土壤节肢动物物种相对多度模型拟合研究. 干旱区资源与环境, 2012, 26(6): 79–84. |
| [19] | Bai Y F, Wu J G, Xing Q, Pan Q M, Huang J H, Yang D L, Han X G. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology, 2008, 89(8): 2140–2153. DOI:10.1890/07-0992.1 |
| [20] | 杨婧, 褚鹏飞, 陈迪马, 王明玖, 白永飞. 放牧对内蒙古典型草原α、β和γ多样性的影响机制. 植物生态学报, 2014, 38(2): 188–200. |
| [21] | Bai Y F, Wu J G, Clark C M, Pan Q M, Zhang L X, Chen S P, Wang Q B, Han X G. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204–1215. |
| [22] | 李静鹏, 郑志荣, 赵念席, 高玉葆. 刈割、围封、放牧三种利用方式下草原生态系统的多功能性与植物物种多样性之间的关系. 植物生态学报, 2016, 40(8): 735–747. |
| [23] | R Development Core Team. R:A Language and Environment for Statistical Computing. Vienna, Austria:R Foundation for Statistical Computing, 2014. |
| [24] | Gideon Schwarz. Estimating the Dimension of a Model. The annals of Statistics, 1978, 6(2): 461–464. DOI:10.1214/aos/1176344136 |
| [25] | 方晓峰, 杨庆松, 刘何铭, 马遵平, 董舒, 曹烨, 袁铭皎, 费希旸, 孙小颖, 王希华. 天童常绿阔叶林中常绿与落叶物种的物种多度分布格局. 生物多样性, 2016, 24(6): 629–638. |
| [26] | 徐国瑞, 张育新, 张霜, 马克明. 海拔对表居土壤动物不同取食功能群多度分布的影响. 土壤学报, 2017, 54(1): 237–245. |
| [27] | Walker T D, Valentine J W. Equilibrium models of evolutionary species diversity and the number of empty niches. The American Naturalist, 1984, 124(6): 887–899. DOI:10.1086/284322 |
| [28] | 李文怀, 郑淑霞, 白永飞. 放牧强度和地形对内蒙古典型草原物种多度分布的影响. 植物生态学报, 2014, 38(2): 178–187. |
| [29] | 施建敏, 范承芳, 刘扬, 杨清培, 方楷, 范方礼, 杨光耀. 石灰岩山地淡竹林演替序列的群落物种多度分布格局. 应用生态学报, 2015, 26(12): 3595–3601. |
| [30] | Harpole W S, Tilman D. Non-neutral patterns of species abundance in grassland communities. Ecology Letters, 2006, 9(1): 15–23. |
| [31] | 张姗, 蔺菲, 原作强, 匡旭, 贾仕宏, 王芸芸, 索炎炎, 房帅, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林草本层物种多度分布格局及其季节动态. 生物多样性, 2015, 23(5): 641–648. |
| [32] | 苏淑兰, 李洋, 王立亚, 郭丁, 康海军, 李旭东, 傅华. 围封与放牧对青藏高原草地生物量与功能群结构的影响. 西北植物学报, 2014, 34(8): 1652–1657. |
| [33] | 杨浩, 白永飞, 李永宏, 韩兴国. 内蒙古典型草原物种组成和群落结构对长期放牧的响应. 植物生态学报, 2009, 33(3): 499–507. |
| [34] | 程佳佳, 米湘成, 马克平, 张金屯. 亚热带常绿阔叶林群落物种多度分布格局对取样尺度的响应. 生物多样性, 2011, 19(2): 168–177. |
| [35] | Magurran A E. Ecological Diversity and Its Measurement. London:Croom Helm Limited, 1982. |