 2017, Vol. 37
2017, Vol. 37文章信息
- 刘玉槐, 严员英, 张艳杰, 严月, 赵玉皓, 徐燕, 陈伏生, 葛体达, 鲁顺保.
- LIU Yuhuai, YAN Yuanying, ZHANG Yanjie, YAN Yue, ZHAO Yuhao, XU Yan, CHEN Fusheng, GE Tida, LU Shunbao.
- 不同温度条件下亚热带森林土壤碳矿化对氮磷添加的响应
- Effects of nitrogen and phosphorus addition under different temperatures on the soil carbon mineralization in a Cunninghamia lanceolata plantation in the subtropics
- 生态学报. 2017, 37(23): 7994-8004
- Acta Ecologica Sinica. 2017, 37(23): 7994-8004
- http://dx.doi.org/10.5846/stxb201610272195
-
文章历史
- 收稿日期: 2016-10-27
- 网络出版日期: 2017-08-14





2. 江西省林业科学院湿地中心, 南昌 330032;
3. 江西农业大学林学院, 南昌 330045;
4. 中国科学院亚热带农业生态研究所区域农业生态研究中心, 长沙 410125
2. Centre for Wetland Jiangxi Forestry Academy, Nanchang 330032, China;
3. College of Forestry, Jiangxi Agricultural University, Nanchang 330045, China;
4. Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China
土壤呼吸是陆地生态系统土壤和大气之间CO2交换的重要组成部分。全球气候变化引起气温升高加速了土壤有机碳在微生物和动物的矿化作用, 其产物通过土壤呼吸作用以CO2形式释放反馈回大气圈, 进一步加剧全球变暖, 因此, 土壤呼吸对环境变化反应最为敏感[1-2], 这也是目前全球变化研究的热点问题, 各国学者对此研究异常活跃, 他们正尝试通过量化各种生态系统碳通量与环境因子之间的关系模型来预测未来全球变化对陆地生态系统的影响[3]。
N、P是植物生长的必需元素, 也是植物从土壤中吸收最多的元素。在土壤中99%以上的氮素是通过微生物矿化作用将其转化为植物可直接吸收的有效态氮[4]。当前全球气温升高、降水格局改变以及大气氮沉降增加等因素正在强烈改变着陆地生态系统的碳汇强度及其分布格局[5]。这些动态的环境因子迫使植被改变其小气候进而影响土壤呼吸。大量的研究表明土壤水分和温度敏感性对土壤碳矿化具有重要影响[6-7], 施氮肥对土壤呼吸的影响也存在较大争议, 如Gallardo等[8]认为施氮肥能够增加土壤呼吸, Kane等[9]的研究表明施氮肥对土壤呼吸速率没有显著的影响, Haynes等[10]对针叶林地施肥结果却显著降低土壤的呼吸速率。
近年来, 大气氮沉降加重[11], 并成为全球化趋势[12], 我国现已是世界第三大氮沉降集中区[13]。全球化背景下大气氮沉降量的增加已成为许多森林的新生态因子[14], 长期的高氮输入可提高土壤碳矿化速率, 导致土壤酸化。土壤呼吸对气候变化(氮沉降、温度和水分)又非常敏感, 且氮沉降、温度及水分对其土壤碳矿化影响机制尚未得到一致结论, 氮沉降和温度变化是亚热带地区面临的重要生态环境问题, 它们对森林地土壤碳矿化的影响如何?土壤呼吸对二者间的响应是否存在差异?均具有重要科学意义。本文以杉木人工林为研究对象, 拟通过模拟气候变化试验, 研究该地区土壤碳矿化对氮沉降以及温度敏感性的响应, 研究结果将有利于了解氮沉降如何引起土壤碳排放的变化, 为降低土壤CO2排放提供参考。
1 研究地区与研究方法 1.1 试验地概况试验地位于江西省泰和县中国科学院千烟洲生态试验站(26°42′N, 115°04′E), 该地区属典型的中亚热带湿润季风气候, 夏季炎热干燥, 冬季寒冷潮湿。年均相对湿度89%, 年均温度在18℃, 6月最高温达29.7℃, 1月最低温可达6.5℃, 年均日照时数1400 h, 年总辐射量约为4349 MJ/m2, 年平均降水量约为1500 mm, 降雨集中在4—6月, 夏季降水量占全年降水量的50%以上。该区域为典型红壤丘陵地貌, 土壤母质是第四纪红粘土。研究区域大气沉降向林地输入的氮为82.8 kg hm-2 a-1), 干沉降为主, 占67%, 大气氨沉降是大气氮沉降主要贡献者, 占大气干沉降82.0%, 占大气氮沉降总量的56%[15]。
1.2 实验设计及样品采集试验样地起始时间为1990年, 1991年4月开始种植植物。施肥自2012年春季起, 每季度各施肥1次, 施肥方式是将氮肥和磷肥拌少量细沙进行均匀撒施。土壤样品采集时间为2015年1月, 在立地条件一致的杉木人工林中, 选取12个20 m×20 m的样方, 将这12个样方随机各选出3个样方作为重复, 每个重复分别进行CK、N1、N2和P处理, 4种处理分别为:CK为不施肥只施沙;N1:氮肥(NH4NO3)量为142.857 kg hm-2 a-1(含氮量为50 kg N hm-2 a-1);N2:285.714 kg hm-2 a-1(含氮量为100 kg N hm-2 a-1);P:磷肥(Na2HPO4)量为229.043 kg hm-2 a-1(含磷量为50 kg P hm-2 a-1), 相邻样方之间留出一个间距为10 m的缓冲带以减少不同施肥带来的影响。在每个样方中以S型选取5个点, 分别用直径7.5 cm的土钻在0—10 cm取土壤样品, 然后将从同一样方的土壤混合, 每份土壤样品约1000 g, 装入无菌自封袋带回实验室。将新鲜土壤样品过2 mm筛去除土壤动物、小石头、可见的植物残体和根系等杂质, 再将每份土壤样品平均分为2份, 一份用于测定土壤含水量、pH、全氮和全碳的含量, 另一份放入4℃冰箱保存, 用于测定土壤可溶性碳氮及生物活性。土壤基本理化性质见表 1。
| 处理 Treatments |
含水量 Moisture/% |
pH | 全氮 Total N/% |
全碳 Total C/% |
碳氮比 C/N |
全磷 Total P/‰ |
| CK | 20.64±1.35a | 4.45±0.12ac | 0.14±0.02a | 2.24±0.23a | 15.84±0.57a | 0.16±0.02b |
| N1 | 19.55±0.81a | 4.19±0.05b | 0.13±0.01a | 2.14±0.10a | 16.36±0.36 | 0.16±0.01b |
| N2 | 19.30±0.93a | 4.21±0.05c | 0.13±0.00a | 2.11±0.10a | 15.76±0.36a | 0.17±0.01b |
| P | 19.08±1.35a | 4.65±0.11a | 0.13±0.02a | 2.02±0.16a | 15.66±0.95a | 0.27±0.01a |
| CK为不施肥只施沙;N1:含氮量为50 kg N hm-2 a-1;N2:含氮量为100 kg N hm-2 a-1;P:含磷量为50 kg P hm-2 a-1。同列不同字母a、b、c表示不同处理间土壤理化性质差异显著(P<0.05) | ||||||
方法参照Chen等[16], 采用元素分析仪(Isoprime-EuroEA3000, 意大利)测定土壤全碳、全氮含量, SOC、SON含量采用2 mol/L KCl浸提, 然后用TOCN分析仪测定SOC和可溶性总氮(total soluble N, TSN), pH值用1:25水溶液、pH计测定, 土壤碳矿化测试方法:先称取新鲜土壤40.0 g, 将其含水量调到田间持水量的60%, 放入4个恒温培养箱(温度分别设为5、15、25、35℃)里进行42 d培养。土壤碳矿化测定方法采用碱液吸收法:2个70 mL的带盖塑料瓶置于1 L的容器内, 其中一个为CO2吸收装置, 其内加入用于吸收CO2的0.1 mol/L NaOH, 另一个装有40.0 g鲜土, 在22℃条件下进行培养, 同时做5个对照试验(仅放置碱液)。培养第1、3、7、14、21、28、35、42 d后将盛有0.1 mol/L NaOH吸收瓶取出后, 先加入1 mol/L BaCl2使得土壤排放的CO2以碳酸盐的形式沉淀, 加入酚酞指示剂后搅拌, 用0.05 mol/L HCl进行滴定剩余的NaOH。对土壤碳矿化进行模拟方程
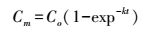
|
式中, Cm为瞬时碳排放量, Co是土壤碳排放量极大值, k为常数, t为培养时间。做潜在最高碳排放量[17], 结合温度敏感性Q10分析, 即温度每升高10℃土壤碳矿化速率所增加的倍数。
1.4 数据处理实验数据采用SPSS 17.0软件进行分析, 对不同处理间的差异显著性进行双因素方差分析, 并用LSD法进行差异显著性检验, 并用SigmaPlot 12.0软件进行数据分析和制图, 显著性差异水平α=0.05。
2 结果与分析 2.1 氮沉降对土壤碳矿化的影响 2.1.1 氮沉降对不同温度条件下土壤累积碳排放量的影响从图 1可知, 在5℃时, 土壤碳矿化累积排放量随氮浓度的升高呈下降趋势;在15℃时, 氮沉降对土壤碳矿化累积排放量具有一定的抑制作用, 但P肥对土壤累积碳排放量具有一定的促进作用;在25℃时, 氮沉降对土壤呼吸累积碳排放量均低于对照组, 且低氮处理抑制作用更强;在35℃条件下, 两者氮沉降处理对土壤呼吸累积碳排放量均表现为一定的抑制作用。在同一温度下, 各处理间均无显著性差异(P > 0.05)。随着温度的升高, 对照CK和N、P添加处理下土壤碳矿化累积碳排放量显著增加(P<0.05, 图 2)。

|
| 图 1 NP添加对杉木林在相同温度下土壤碳矿化量的影响 Fig. 1 Effects of substrate addition on CO2-C emission under Cunninghamia lanceolata in the same temperature CK为不施肥只施沙;N1:含氮量为50 kg N hm-2 a-1;N2:含氮量为100 kg N hm-2 a-1;ns:差异不显著(P > 0.05)。图中数据为平均值±标准误 |

|
| 图 2 底物添加在不同温度下对杉木林土壤碳矿化量的影响 Fig. 2 Effects of substrate addition on CO2-C evolved under Cunninghamia lanceolata in different temperature 不同小写字母表示同一处理不同温度间差异显著(P < 0.05), 图中数据为平均值±标准差 |
从图 2可以看出, 温度升高显著提升了林地土壤碳矿化量, 不同处理间, 不同温度对土壤碳排放量均有一定的影响。在CK处理中, 5℃和15℃间无显著性差异, 但均显著低于25℃和35℃对土壤碳矿化的影响(P<0.05), 35℃对土壤碳排放量影响显著高于25℃。N1处理中, 于35℃时CO2累计排放量显著高于其他温度土壤碳排放量(P < 0.05), 但5、15、25℃间无显著差异。N2处理中, 随温度升高CO2土壤累计排放量逐渐增加, 其中5℃与15℃, 15℃与25℃之间差异不显著, 但5℃显著低于25℃、后者显著低于35℃(P<0.05)。添加P处理中, 不同温度对CO2累计排放量与N2变化规律类似, 即35℃处理下碳排放量最大, 其次为25℃和15℃, 最小排放量为5℃处理。
2.1.2 氮沉降对不同温度条件下土壤碳矿化速率的影响由图 3可知, 在同一温度下, 不同处理土壤矿化速率大致趋势是先急速下降然后趋于平缓。在5℃时, 由于温度较低, 各处理土壤矿化率偏低, N2、P二者呈先缓慢上升再趋于平缓的单峰曲线, CK、N1培养3 d内呈急速下降, 并且N1在前3天土壤矿化速率高于CK, P处理最低, 在第7天后变化缓慢;在15℃时, 对照CK在第7天略有升高外, 其他处理都是先下降再趋于平缓;在25℃时, P处理后的第3天后土壤矿化速率开始下降, 其他处理都是先下降再趋于稳定, N1处理土壤矿化速率始终为最低;在35℃时, 各处理都是先下降后趋于平稳, NP添加对土壤矿化速率影响的规律为:P > N1 > N2。从图 3还可以看出, 温度对土壤碳矿化速率的影响显著。

|
| 图 3 不同底物添加对杉木林土壤矿化速率的影响 Fig. 3 Effects of different substrate addition on rate of Cunninghamia lanceolata soil respiration |
从图 4可以看出, 对土壤在不同温度条件下, 做土壤全碳和累积碳排放量进行分析得出, 在5℃和15℃条件下, 土壤全碳对土壤碳排放量无显著性差异, 在25℃条件下, 土壤全碳与土壤碳排放量呈类似开口朝上的抛物线的变化趋势, 即随土壤全碳含量的增加, 土壤碳排放量先减小后增加。在35℃条件下, CO2累积排放量随土壤全碳含显著增加(P<0.05), 对其全碳和碳排放量进行回归分析, 得回归方程为:y=-10.1595+143.201x, R2=0.38。

|
| 图 4 全碳对不同氮添加处理杉木林土壤碳矿化的影响 Fig. 4 Effects of total carbon for Cunninghamia lanceolata soil carbon minerlization under different nitrogen addition |
从图 5可以看出, 在5、15、35℃条件下, 土壤DOC对土壤碳排放量影响无显著性差异(P > 0.05)。在25℃条件下, 土壤碳排放量随DOC含量先增后减, 呈类似开口朝下的抛物线变化规律, 且差异达显著性(P<0.05)。在同一温度条件下, 土壤DON对土壤累积碳排放量均无显著性差异(P > 0.05)。

|
| 图 5 可溶性有机碳对不同氮添加处理杉木林土壤碳矿化的影响 Fig. 5 Effects of Dissolved Organic for Cunninghamia lanceolata soil carbon minerlization under different nitrogen addition |
从图 6可以看出, 在5℃时, 不同处理下杉木林土壤碳排放量随土壤含水量增加而增加, 且差异达显著性水平(P < 0.05)。在其他温度条件下并无显著性差异, 在同一含水量的条件下, 土壤碳排放量随温度的升高而增加, 说明土壤含水量并不是影响土壤碳排放量的调控因子。

|
| 图 6 土壤含水量对不同氮添加处理杉木林土壤碳矿化的影响 Fig. 6 Effect of moisture for Cunninghamia lanceolata soil carbon minerlization under different nitrogen addition |
在没有外界干扰的情况下长时间培养后土壤累积碳排放量将会达到一个峰值。对杉木林地土壤进行氮添加处理, 在42 d的室内培养实验中, 运用方程Cm=Co(1-exp-kt)对土壤潜在碳排放进行模拟, 得出在不同温度不同处理下杉木林土壤最大碳排放量(表 2), 相关系数R2基本在0.9以上, 说明拟合效果很好。在5℃条件下, N会抑制土壤碳矿化, 且随N浓度升高抑制作用越强, P处理能促进土壤碳排放;15℃和35℃条件下, NP处理对土壤碳排放量均有抑制作用;在25℃条件下, 不同处理对杉木林土壤潜在排放量影响不显著。
| 温度/℃ Temperature |
处理 Treatments |
潜在碳排放量 Potentially carbon mineralizable |
参数 Constant (K) |
相关系数 Relation coefficient (R2) |
| 5 | CK | 138.663±22.858ab | 0.032±0.005a | 0.941±0.018a |
| N1 | 134.848±18.117ab | 0.026±0.007ab | 0.921±0.040a | |
| N2 | 77.133±1.532b | 0.037±0.020a | 0.960±0.014a | |
| P | 206.517±104.477a | 0.016±0.009b | 0.925±0.023a | |
| 15 | CK | 315.946±21.335a | 0.054±0.034b | 0.887±0.007a |
| N1 | 85.397±20.987bc | 0.102±0.079a | 0.954±0.015a | |
| N2 | 74.193±18.464c | 0.062±0.040ab | 0.840±0.040a | |
| P | 111.495±12.358b | 0.098±0.085a | 0.680±0.139b | |
| 25 | CK | 176.355±22.848a | 0.062±0.004a | 0.955±0.017a |
| N1 | 174.745±26.953a | 0.041±0.010b | 0.987±0.005a | |
| N2 | 187.938±21.846a | 0.041±0.001b | 0.987±0.002a | |
| P | 186.869±6.041a | 0.053±0.007a | 0.983±0.012a | |
| 35 | CK | 488.887±35.656a | 0.025±0.002b | 0.991±0.003a |
| N1 | 348.949±64.454ab | 0.033±0.002b | 0.972±0.011a | |
| N2 | 315.799±48.418b | 0.041±0.007a | 0.981±0.004a | |
| P | 477.196±130.299a | 0.031±0.009a | 0.974±0.021a | |
| 同一列不同小写字母表示同一温度不同处理间差异显著(P < 0.05), 图中数据为平均值±标准差 | ||||
土壤矿化与温度的关系还可以从温度敏感性角度来研究(Q10), 因为二氧化碳产生的温度敏感性可作为有机质降解温度敏感性的主要指标。本研究中Q10培养42d的结果见表 3, 其范围为1.5149—1.8354, 从表 3可以看出, N和P处理对土壤Q10值均有下降趋势, 其中N浓度升高, Q10值下降, 对不同处理的Q10值做方差分析得出它们无显著差异。
| 处理Treatment | Q10 value Q10值 |
| CK | 1.8354±0.1932 |
| N1 | 1.6454±0.1909 |
| N2 | 1.6189±0.1125 |
| P | 1.5149±0.0500 |
土壤矿化是一个非常复杂的生物学过程, 受土壤温度、土壤含水量、土壤C、N含量等非生物因子以及植被类型、根系等生物因子综合影响[18]。温度是影响土壤矿化一个重要的因子, 土壤矿化和温度之间具有显著的相关关系[19-20]。本研究结果表明, 温度升高能显著提升林地土壤碳矿化量。在不同氮沉降处理下, 温度对土壤碳排放量均存在不同程度的影响。在对照处理中, 25℃和35℃对土壤碳排放量影响显著, 且显著高于5℃和15℃间对土壤碳矿化的影响(P<0.05), 可能原因是随着温度的升高, 土壤中的微生物活性增加促进了土壤矿化, 增温在短时间内可刺激土壤矿化产生大量CO2[21-22]。在森林生态系统中, 土壤碳矿化受温度的影响较为复杂, 主要是温度对土壤生物活性的影响, 往往同时还受土壤外源N添加的影响[20]。在N1处理中, 35℃时CO2累计排放量显著高于其他温度对土壤碳排放量的影响(P < 0.05), 且其他温度间无显著差异。在N2处理中, 随温度升高CO2土壤累计排放量逐渐增加, 且5℃与25、35℃间的差异显著(P<0.05), 外源氮的输入可能导致根系氮浓度的增加, 进而影响土壤矿化作用[23]。在同一温度下, 与对照组相比, 土壤碳排放量均随氮浓度的升高而减少, 说明氮沉降对土壤碳呼吸具有抑制作用, 在25℃和35℃条件下, 氮抑制呼吸作用, 高温度条件下抑制作用更强[24-25], 主要原因是高氮处理降低土壤pH值, 导致土壤微生物活性降低, 分解有机碳的速率降低, 从而抑制了土壤矿化[26], 这种现象在针叶林和阔叶林中的影响更为明显[27]。也可能是由于氮素的增加降低土壤C/N进而改变土壤微生物的呼吸进而影响土壤矿化[28-30], 也有学者认为氮添加对土壤矿化的无影响或促进作用[31-32]。在不同温度条件下, 土壤有机碳矿化速率呈日趋下降, 培养21 d后矿化速率趋于平缓, 其原因可能是土壤可利用性碳、氮基质的量是影响其矿化的主要因素, 因此, 在矿化培养实验中, 前期土壤易矿化的有机碳数量较多, 已被微生物分解, 培养到中后期土壤碳矿化主要受到可利用碳含量的限制, 微生物开始难矿化有机碳, 使得分解速率下降, CO2释放量开始降低[33]。较高温度培养过的样品CO2释放速率显著低于较低温度, 很多长期培养试验中, 都观测到基质大量损耗后, 试验后期土壤矿化速率的降低[34], Yoshitake等发现添加氮在开始会刺激土壤矿化, 但不会持续下去[26], 这也与本研究中土壤矿化率在后期降低并趋于稳定相一致[35]。随着温度的升高, CK和N、P添加在短时间内对土壤矿化碳排放呈显著性增加趋势(P<0.05)。添加P处理中, 不同温度对CO2累计排放量与N2变化规律类似, 可能的原因是P对土壤微生物比较敏感抑制土壤矿化[36-37]。
有关土壤矿化速率与温度的关系研究比较多, 他们之间拟合的曲线方程较为复杂。本研究从温度敏感性角度来分析(Q10)土壤矿化速率受温度的影响, Q10培养范围为1.5149—1.8354, 变异比较小, 说明氮沉降的影响表明氮添加处理对杉木人工林土壤有机碳矿化对温度变化的敏感性不是特别明显, 这与王红等研究的结果不一致[38], 其原因可能是土壤矿化是一个十分复杂的生物学过程, Q10值除受温度的影响外, 还受土壤水分、根生物量及其他因子的综合结果[28], 因此, 氮添加对土壤矿化温度敏感性影响的机理需要进一步细化研究。总之, 未来气候变化对亚热带森林生态系统碳循环的影响及其机理会更为复杂, 这将是今后的主要研究方向之一。
在森林生态系统中, 土壤矿化是土壤碳库主要输出途径, 因此, 土壤碳库含量的高低对土壤碳矿化作用产生重要影响。从土壤全碳与土壤碳矿化的关系来看, 在5℃和15℃条件下, 土壤全碳对土壤碳排放量无显著性差异, 在25℃和35℃条件下, 土壤全碳对土壤碳排放量的影响均达差异显著(P<0.05), 说明土壤全碳对土壤碳矿化影响同时也受温度调控。土壤碳库以及土壤氮素作为土壤有机碳矿化的底物, 其含量必将直接对土壤有机碳矿化作用产生影响[39-40]。但生态系统中非生物因子和生物因子间相互关系的复杂性, 使得土壤有机碳矿化研究仍存在不确定性, 如Giardina等认为温度的升高不会对土壤有机碳矿化产生影响[41], 本研究结果表明在25℃条件下, 土壤碳排放量随DOC含量先增后减(P<0.05), 而其他温度条件下, 土壤DOC对土壤碳排放量并无显著性影响。且在同一温度条件下, 土壤DON对土壤累积碳排放量均无显著性差异。土壤矿化除受土壤碳库影响外, 可能还与土壤含水量密切相关, 由于土壤含水量对呼吸速度影响较为复杂, 往往同时取决于温度的配置状况。在一定温度条件下, 土壤含水量适度提高不仅可以增强微生物的活性, 也可以增加土壤中微生物的数量, 从而加快土壤碳的矿化速度。本研究结果表明, 在5℃时, 杉木林土壤碳矿化量随土壤含水量的增加而显著增加(P<0.05), 在其他温度条件下并无显著性差异。在同一含水量的条件下, 土壤碳排放量随温度的升高而增加, 这与结论与Hartley研究的结果一致[35]。温度对土壤有机碳矿化作用的影响可能会因土壤有机碳库以及土壤水分等因子的限制作用而减弱, 并且对于土壤水分以及土壤碳、氮含量等因子对土壤有机碳矿化的影响也存在较大的争议[40]。因此, 土壤碳矿化对温度的响应是土壤性质、生物活性以及可利用性碳、氮综合作用的结果[33]。
目前, 室内培养方法广泛应用于土壤潜在碳排放的研究, 由于土壤矿化在没有根系或气候因子等干扰的情况下, 土壤矿化在长时间培养后终将达到平衡。本研究采用单库模式一级动力学方程Cm=Co(1-exp-kt)对土壤矿化潜在碳排放进行模拟, 得出相关系数均在0.9以上。在25℃条件下, 不同处理对杉木林土壤潜在排放量影响不显著。在其他温度条件下, N添加均会抑制土壤潜在碳排放量, 且随N浓度升高抑制作用越强(P<0.05)。
4 结论氮沉降对土壤矿化碳的累积排放量具有抑制作用。随着温度的升高, CK、N和P添加对土壤矿化碳排放呈显著性增加趋势(P<0.05)。不同处理间, 不同温度对土壤碳排放量均有一定的影响。不同温度条件下添加P处理中对CO2累计排放量与N2变化规律类似。在5℃条件下, N会抑制土壤碳矿化, 且随N浓度升高抑制作用越强, P处理能促进土壤碳排放;15℃和35℃条件下, NP处理对土壤碳排放量均有抑制作用;25℃时不同处理对土壤潜在排放量影响不显著。
在较高温度(25℃和35℃)培养下, 土壤全碳对CO2累积排放量均有显著性影响, 较低的温度(5℃和15℃)培养下, 土壤全碳对土壤碳排放量无显著性差异。在25℃条件下, 土壤DOC含量对土壤碳排放量的影响显著(P<0.05), 其他温度无影响。在同一温度条件下, 土壤DON对土壤累积碳排放量均无显著性差异。在5℃时, 不同处理下土壤含水量对土壤碳排放量影响显著, 在其他温度条件下并无显著性差异。
氮沉降和P处理对土壤Q10值与对照相比均有下降趋势, 随N浓度升高Q10值下降, 对不同处理的Q10值做方差分析得出它们无显著差异。
| [1] | 姜艳, 王兵, 汪如玉, 杨清培. 亚热带林分土壤呼吸及其与土壤温湿度关系的模型模拟. 应用生态学报, 2010, 21(7): 1641–1648. |
| [2] | Zerva A, Mencuccini M. Short-term effects ofclearfelling on soil CO2, CH4, and N2O fluxes in a Sitka spruce plantation. Soil Biology and Biochemistry, 2005, 37(11): 2025–2036. DOI:10.1016/j.soilbio.2005.03.004 |
| [3] | Li S G, Asanuma J, Eugster W, Kotani A, Liu J J, Urano T, Oikawa T, Davaa G, Oyunbaatar D, Sugita M. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia. Global Change Biology, 2005, 11(11): 1941–1955. |
| [4] | 栾军伟, 向成华, 骆宗诗, 宫渊波. 森林土壤呼吸研究进展. 应用生态学报, 2006, 17(12): 2451–2456. DOI:10.3321/j.issn:1001-9332.2006.12.044 |
| [5] | Piao S L, Fang J Y, Ciais P, Peylin P, Huang Y, Sitch S, Wang T. The carbon balance of terrestrial ecosystems in China. Nature, 2009, 458(7241): 1009–1013. DOI:10.1038/nature07944 |
| [6] | Cox P M, Betts R A, Jones C D, Spall S A, Totterdell I J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 2000, 408(6809): 184–187. DOI:10.1038/35041539 |
| [7] | Xu M, Qi Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology, 2001, 7(6): 667–677. DOI:10.1046/j.1354-1013.2001.00435.x |
| [8] | Gallardo A, Schlesinger W H. Factors limiting microbial biomass in the mineral soil and forest floor of a warm-temperate forest. Soil Biology and Biochemistry, 1994, 26(10): 1409–1415. DOI:10.1016/0038-0717(94)90225-9 |
| [9] | Kane E S, Pregitzer K S, Burton A J. Soil respiration along environmental gradients in Olympic National Park. Ecosystems, 2003, 6(4): 326–335. DOI:10.1007/s10021-002-0115-7 |
| [10] | Haynes B E, Gower S T. Belowground carbon allocation in unfertilized and fertilized red pine plantations in northern Wisconsin. Tree Physiology, 1995, 15(5): 317–325. DOI:10.1093/treephys/15.5.317 |
| [11] | Vitousek P M, Aber J D, Howarth R W, Likens E L, Matson P A, Schindler D W, Schlesinger W H, Tilman D G. Human alteration of the global nitrogen cycle:sources and consequences. Ecological Applications, 1997, 7(3): 737–750. |
| [12] | Matson P, Lohse K A, Hall S J. The globalization of nitrogen deposition:consequences for terrestrial ecosystems. Ambio:A Journal of the Human Environment, 2001, 31(2): 113–119. |
| [13] | Mo J M, Brown S, Xue J H, Fang Y T, Li Z. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant and Soil, 2006, 282(1/2): 135–151. |
| [14] | Kazda M. Indications of unbalanced nitrogen nutrition of Norway spruce stands. Plant and Soil, 1990, 128(1): 97–101. DOI:10.1007/BF00009401 |
| [15] | 樊建凌, 胡正义, 庄舜尧, 周静, 王体健, 刘翠英. 林地大气氮沉降的观测研究. 中国环境科学, 2007, 27(1): 7–9. |
| [16] | Chen C R, Xu Z H, Mathers N J. Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Science Society of America Journal, 2004, 68(1): 282–291. DOI:10.2136/sssaj2004.2820 |
| [17] | Smith J L, Schobel R R, McNeal B L, Campbell G S. Potential errors in the first-order model for estimating soil nitrogen mineralization potentials. Soil Science Society of America Journal, 1980, 44(5): 996–1000. DOI:10.2136/sssaj1980.03615995004400050025x |
| [18] | 李玉宁, 王关玉, 李伟. 土壤呼吸作用和全球碳循环. 地学前缘, 2002, 9(2): 351–357. |
| [19] | 严俊霞, 汤亿, 李洪建. 城市绿地土壤呼吸与土壤温度土壤水分的关系研究. 干旱区地理, 2009, 32(4): 604–609. |
| [20] | Menyailo O V, Huwe B. Denitrification and C, N mineralization as function of temperature and moisture potential in organic and mineral horizons of an acid spruce forest soil. Journal of Plant Nutrition and Soil Science, 1999, 162(5): 527–531. DOI:10.1002/(ISSN)1522-2624 |
| [21] | 杨庆朋, 徐明, 刘洪升, 王劲松, 刘丽香, 迟永刚, 郑云普. 土壤呼吸温度敏感性的影响因素和不确定性. 生态学报, 2011, 31(8): 2301–2311. |
| [22] | Mikan C J, Schimel J P, Doyle A P. Temperature controls of microbial respiration in arctic tundra soils above and below freezing. Soil Biology and Biochemistry, 2002, 34(11): 1785–1795. DOI:10.1016/S0038-0717(02)00168-2 |
| [23] | Eissenstat D M, Wells C E, Yanai R D, Whitbeck J L. Building roots in a changing environment:implications for root longevity. New Phytologist, 2000, 147(1): 33–42. DOI:10.1046/j.1469-8137.2000.00686.x |
| [24] | 吴浩浩, 徐星凯, 李团胜, 段存涛. 碳氮添加对不同湿润程度的温带森林土壤氧化亚氮排放影响. 科学通报, 2014, 59(14): 1337–1347. |
| [25] | Zhao S C, Qiu S J, Cao C Y, Zheng C L, Zhou W, He P. Responses of soil properties, microbial community and crop yields to various rates of nitrogen fertilization in a wheat-maize cropping system in north-central China. Agriculture, Ecosystems & Environment, 2014, 194: 29–37. |
| [26] | Yoshitake S, Sasaki A, Uchida M, Funatsu Y, Nakatsubo T. Carbon and nitrogen limitation to microbial respiration and biomass in an acidicsolfatara field. European Journal of Soil Biology, 2007, 43(1): 1–13. DOI:10.1016/j.ejsobi.2006.01.003 |
| [27] | 王效科, 白艳莹, 欧阳志云, 苗鸿. 全球碳循环中的失汇及其形成原因. 生态学报, 2001, 22(1): 94–103. |
| [28] | Janssens I A, Dieleman W, Luyssaert S, Subke J A, Reichstein M, Ceulemans R, Ciais P, Dolman A J, Grace J, Matteucci G, Papale D, Piao S L, Schulze E D, Tang J, Law B E. Reduction of forest soil respiration in response to nitrogen deposition. Nature Geoscience, 2010, 3(5): 315–322. DOI:10.1038/ngeo844 |
| [29] | Traore S, Thiombiano L, Millogo J R, Guinko S. Carbon and nitrogen enhancement incambisols andvertisols by Acacia spp. in eastern Burkina Faso:relation to soil respiration and Microbial biomass. Applied Soil Ecology, 2007, 35(3): 660–669. DOI:10.1016/j.apsoil.2006.09.004 |
| [30] | Persson T, Wirén A. Microbial activity in forest soils in relation to acid/base and carbon/nitrogen status. Communications of the Norwegian Forest Research Institute, 1989, 81(suppl): S15–21. |
| [31] | Cheshire M V, Chapman S J. Influence of the N and P status of Plant material and of added N and P on the mineralization of C from 14C-labelled ryegrass in soil. Biology and Fertility of Soils, 1996, 21(3): 166–170. DOI:10.1007/BF00335929 |
| [32] | Bradford M A, Fierer N, Reynolds J F. Soil carbon stocks in experimentalmesocosms are dependent on the rate of labile carbon, nitrogen and Phosphorus inputs to soils. Functional Ecology, 2008, 22(6): 964–974. DOI:10.1111/fec.2008.22.issue-6 |
| [33] | 沈征涛, 施斌, 王宝军, 姜洪涛. 土壤有机质转化及CO2释放的温度效应研究进展. 生态学报, 2013, 33(10): 3011–3019. |
| [34] | Hartley I P, Hopkins D W, Sommerkorn M, Wookey P A. The response of organic mattermineralisation to nutrient and substrate additions in sub-arctic soils. Soil Biology and Biochemistry, 2010, 42(1): 92–100. DOI:10.1016/j.soilbio.2009.10.004 |
| [35] | Jarvis P, Linder S. Botany:constraints to growth of boreal forests. Nature, 2000, 405(6789): 904–905. DOI:10.1038/35016154 |
| [36] | Keith H, Jacobsen K L, Raison R J. Effects of soil phosphorus availability, temperature and moisture on soil respiration in Eucalyptuspauciflora forest. Plant and Soil, 1997, 190(1): 127–141. DOI:10.1023/A:1004279300622 |
| [37] | 王红, 范志平, 邓东周, 陈德龙, 孙学凯, 高俊刚, 曾德慧. 不同环境因子对樟子松人工林土壤有机碳矿化的影响. 生态学杂志, 2008, 27(9): 1469–1475. |
| [38] | Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165–173. DOI:10.1038/nature04514 |
| [39] | Hopkins D W, Sparrow A D, Elberling B, Gregorich E G, Novis P M, Greenfield L G, Tilston E L. Carbon, nitrogen and temperature controls on microbial activity in soils from an Antarctic dry valley. Soil Biology and Biochemistry, 2006, 38(10): 3130–3140. DOI:10.1016/j.soilbio.2006.01.012 |
| [40] | 杨继松, 刘景双, 孙丽娜. 温度、水分对湿地土壤有机碳矿化的影响. 生态学杂志, 2008, 27(1): 38–42. |
| [41] | Giardina C P, Ryan M G. Evidence that decomposition rates of organic carbon in mineral soil do not vary with temperature. Nature, 2000, 404(6780): 858–861. DOI:10.1038/35009076 |