 2017, Vol. 37
2017, Vol. 37文章信息
- 王恒方, 吕光辉, 周耀治, 曹靖.
- WANG Hengfang, LÜ Guanghui, ZHOU Yaozhi, CAO Jing.
- 不同水盐梯度下功能多样性和功能冗余对荒漠植物群落稳定性的影响
- Effects of functional diversity and functional redundancy on the stability of desert plant communities under different water and salt gradients
- 生态学报. 2017, 37(23): 7928-7937
- Acta Ecologica Sinica. 2017, 37(23): 7928-7937
- http://dx.doi.org/10.5846/stxb201610192139
-
文章历史
- 收稿日期: 2016-10-19
- 网络出版日期: 2017-08-14





2. 绿洲生态教育部重点实验室, 乌鲁木齐 830046;
3. 新疆大学干旱生态环境研究所, 乌鲁木齐 830046
2. Key Laboratory of Oasis Ecology, Ministry of Education, Urumqi 830046, China;
3. Institute of Arid Ecology and Environment, Xinjiang University, Urumqi 830046, China
随着全球生物多样性的迅速丧失, 物种多样性和生态系统稳定性的关系以及他们的维持机制已经成为人类面临的重要科学问题[1-4]。影响生态系统稳定性的因子众多, 生态学家关注最多的是物种多样性对其的影响[4]。人们提出很多关于物种多样性和生态系统功能的假说, 包括多样性-稳定性假说[2], 铆钉假说[5], 物种冗余假说[6], 不确定假说[7]和关键种假说[8]等。研究表明物种多样性和稳定性间可能存在正相关[1-3, 9]、负相关[10]和非线性的关系[11], 并没有一致的结论。Bello等认为物种多样性包括功能多样性和功能冗余两个部分[12], 功能多样性和功能冗余中任何一个因子的变化都可能导致生态系统稳定性的变化。
功能多样性是联系物种多样性和生态系统功能的关键性因素[13], 是表示种间功能属性相对差异性和生态互补性的指标, 通常认为在决定生态系统功能上比物种多样性有更直接的作用。植物的功能多样性在阐明环境变化、群落组成及与生态系统的关系中起到了重要的作用, 功能多样性包括功能丰富度、功能均匀度和功能分离度。常用的方法是以Rao′s功能多样性指数代表功能多样性, Rao′s二次方程整合了物种丰富度和物种之间的功能特征差异。可以很好的解释物种间趋异性和物种在群落中的比例的表征[14]。通常认为当功能多样性随物种多样性增加表现为饱和增加时, 就预示着存在功能冗余。Walker[6]在冗余假说中提出在一个包含许多物种的功能群中, 物种通常表现出功能冗余。也就是说, 一些物种呈现出类似的或相同的功能, 并且对环境的变化有异步响应, 或显示出时间生态位分化的现象[6, 15], 这些物种在群落受到干扰后有助于保持群落的稳定性[16-18]。Bello等[12]提出了一种计算功能冗余的方法, 并认为功能冗余是物种多样性除去功能多样性, 即物种多样性与功能多样性的差异。
本文主要研究了不同水盐梯度下荒漠区植被群落功能多样性和功能冗余的关系。在干旱区的空间资源再分配中的研究中, 水盐是影响多样性和生态系统功能的重要的环境因素, 分析了研究区植被群落的多样性和群落稳定性的变化, 以及他们在不同水盐梯度下的相互关系。试图回答以下问题:(1)冗余假说(冗余可以提高稳定性)在该研究区是否成立。(2)功能多样性和功能冗余哪个对稳定性的影响更大。(3)水分和盐分哪个因素对群落的功能多样性、功能冗余及群落稳定性影响更大。该研究除了可以加深人们对多样性和稳定性关系的了解外, 也可以为荒漠区植被的保护和恢复提供科学依据。
1 研究方法 1.1 研究区概况研究区位于艾比湖国家级湿地自然保护区(79°53′—85°02′ E, 43°38′—45°52′ N)。该区域属北温带大陆性干旱气候, 年均降水量为107 mm, 年均蒸发量1600 mm以上, 日照时数约2800 h, 地处温带又位于阿拉山口大风的主风道上, 年平均大风(大于17 m/s)日数多达164 d。土壤类型丰富多样, 典型的土壤为灰漠土、灰棕漠土和风沙土[19], 多样化的土壤类型孕育了丰富的植物群落, 荒漠旱生植物物种尤其丰富, 主要植物种类有胡杨(Populus euphratica)、梭梭(Sacsaoul)、芦苇(Reed)、柽柳(Tamarix chinensis)、盐穗木(Halostachys caspica)、盐爪爪(Kalidium foliatum)、琵琶柴(Reaumuria songonica)和骆驼刺(Alhagi sparsifolia)等。
1.2 实验设计 1.2.1 样方设置在艾比湖国家级湿地自然保护区的荒漠区内, 按土壤水分和盐分可能沿着离河距离的变化梯度, 垂直于艾比湖补给河流之一的阿其克苏河, 沿河岸在保护区东大桥管护站以北布设样地, 样地东西相距2 km, 南北相距3 km, 共6 km2。将样地平均划分为30个400 m×500 m小样地, 在每个小样地的中心点设置一个30 m×30 m的样方, 沿从东到西方向共5条样带, 每条样带6个样方, 总计30个30 m×30 m的样方。

|
| 图 1 研究区及样方分布示意图 Fig. 1 The study area and location of the plots |
在每个30 m×30 m的样方中调查乔木, 沿对角线取2个5 m×5 m的灌木样方, 在每个乔木样方的对角线上各取2个1 m×1 m的草本样方。调查时主要记录乔木植物的种名、多度、胸径、株高、冠幅, 灌木和草本植物调查种名、丛数、丛幅、高度和盖度。同时记录各样方的经纬度。
在每个样方中, 选取每个物种的5片不同健康叶片, 利用游标卡尺测定叶片厚度, 同时用便携式叶绿素检测仪(SPAD-502)测定植物叶片的相对叶绿素含量。用叶面积仪测定叶面积, 梭梭、沙拐枣等形状近似圆柱体的叶片, 利用游标卡尺测定叶片直径和长度, 按圆柱体表面积计算公式求出叶片总面积。然后带回实验室烘干, 用万分之一天平(AL204, Mettler Toledo Group, 上海)称重, 计算平均叶面积(m2)、比叶重(kg/m2)和比叶面积(m2/kg)。
每个样方中, 采用“S”采样法, 选取3个土壤样点(包含一份乔木冠下土、一份灌木冠下土以及一份裸地土)采集0—15cm土样, 除去凋落物, 分别装入已称重的铝盒和密封袋。立即称量铝盒和土壤的质量, 带回实验室测定土壤含水量和土壤含盐量。
1.3 数据计算与统计分析 1.3.1利用SPSS 21.0, 采用系统聚类中的wards方法对样方进行系统聚类, 利用多重比较的Duncan检验方法, 对土壤的含水量及含盐量等理化性质进行差异性检验(表 2)。
| 样地 Plots |
样地编码 Plots Number |
土壤含水量/% Soil moisture content |
土壤含盐量/% Soil salt content |
| 高水高盐 High water and salt | H | 16.916±3.098a | 0.7884±0.2192a |
| 中水中盐 Moderate water and salt | M | 9.136±1.078b | 0.6179±0.1539b |
| 低水低盐 Low water and salt | L | 5.447±1.668c | 0.4232±0.0975c |
| 注:同一列相同字母表示不同水盐梯度下土壤理化性质无显著差异(P>0.05) | |||
通过群落调查, 以19个物种(表 1)的4个定性性状(科、生活周期、叶表面类型、生长型)和6个定量性状(冠幅、株高、叶厚度、比叶重、比叶面积和相对叶绿素含量)共10个功能特征类型(表 3), 用于计算功能多样性。在计算功能多样性时, 对物种×性状状态数据矩阵中每个物种定性性状进行赋值分类(表 3)。
| 序号 Id |
植物种名 Plant species name |
科 Family |
| 1 | 胡杨 Populus euphratica | 杨柳科 |
| 2 | 梭梭 Haloxylon ammodendron | 藜科 |
| 3 | 柽柳 Tamarix ramosissima | 柽柳科 |
| 4 | 铃铛刺 Halimodendron halodendron | 豆科 |
| 5 | 盐穗木 Halostachys caspica | 藜科 |
| 6 | 白刺 Nitraria schoberi | 蒺藜科 |
| 7 | 盐爪爪 Kalidium foliatum | 藜科 |
| 8 | 小叶碱蓬 Suaeda microphylla | 藜科 |
| 9 | 盐节木 Halocnemum strobilaceum | 藜科 |
| 10 | 骆驼刺 Alhagi sparsifolia | 豆科 |
| 11 | 琵琶柴 Reaumuria soongorica | 柽柳科 |
| 12 | 罗布麻 Apocynum venetum | 夹竹桃科 |
| 13 | 花花柴 Karelinia caspica | 菊科 |
| 14 | 芦苇 Phragmites australis | 禾本科 |
| 15 | 沙篷 Agriophyllum squarrosum | 藜科 |
| 16 | 绢蒿 Seriphidium amoenum | 菊科 |
| 17 | 甘草 Glycyrrhiza uralensis | 豆科 |
| 18 | 猪毛菜 Salsola collina | 藜科 |
| 19 | 盐地碱篷 Suaeda salsa | 藜科 |
| 功能特征类型 Functional characteristics types |
数据类型 Type of data |
植物功能特征类型及赋值 Plant functional types and characteristics assignment |
| 科 Family | 名称 | 杨柳科(1)、藜科(2)、柽柳科(3)、豆科(4)、蒺藜科(5)、夹竹桃科(6)、菊科(7)、禾本科(8) |
| 生活型 Life form | 名称 | 多年生(1)、一年或两年生(2) |
| 叶表面类型 Blade surface type | 名称 | 光滑(1)、具毛(2)、具刺(3) |
| 生长型 Growth type | 名称 | 乔木(1)、小乔木(2)、灌木(3)、半灌木(4)、草本(5) |
| 冠幅 Crown width | 数值 | 植物的东西长*南北长, 单位:m2 |
| 株高 Plant height | 数值 | 单位:m |
| 叶厚度 Blade thickness | 数值 | 单位:cm |
| 比叶重 Leaf Mass per Area | 数值 | 单位:g/cm2 |
| 比叶面积 Specific Leaf Area | 数值 | 单位:cm2/g |
| 叶绿素 Chlorophyll | 数值 | 叶绿素相对含量, 单位:% |
采用Rao′s指数对30个样方的功能多样性进行计算, 由于物种功能特征值是数量、名称的混合型数据, 所以计算中用到的距离公式都采用Gower′s距离公式[20-21], 使用R(3.2.5)软件进行计算。Rao′s计算公式如下[22]:
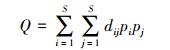
|
(1) |
S为样方内的物种数;pi和pj分别为第i和第j个物种占总物种个体数的比例;dij为Gower′s距离, dij在0(两物种具有完全相同的特征)至1(两物种具有完全不同的特征)之间, 表示物种i, j在一组性状空间中的相异性。Rao系数描述了种间功能性状的相对差异, 比其他功能多样性指数更适合表征群落的种间趋异性, 也能更好的将物种丧失与生态系统功能变化相联系。
1.3.3 功能冗余FR计算采用Bello等[12]的公式计算:
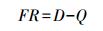
|
(2) |
式中, D为Shannon-Wiener多样性指数, 代表物种多样性;Q为表示功能多样性的Rao系数, 代表功能多样性。D的计算公式如下[23]:
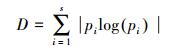
|
(3) |
式中, S为样方内的物种数;pi为第i个物种占总物种个体数的比例。
1.3.4 群落稳定性的两种计算方法(1) 群落稳定性以物种种群密度变异系数(coefficient of variation, CV)的倒数ICV表示[24-26]:
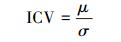
|
(4) |
式中, μ为样方中各物种的平均密度, σ为各物种密度的标准差。ICV值越大, 群落稳定性越高, 因为相对于平均密度, 各物种密度的变异性较小。
(2) 以改进后的Godron稳定性测定方法[27]来测定群落的稳定性。具体方法为:首先把样地中不同种植物的盖度由大到小的顺序排列, 换算成相对盖度, 按相对盖度由大到小的顺序逐步累加起来;然后将样地内植物种类的总和取倒数, 按植物种类的顺序逐步累积;再将植物种类倒数百分率同累积相对盖度一一对应, 画出散点图并用平滑曲线连接起来。然后再与直线y=100-x相交, 交点即为所求群落的稳定性比值。一般地, 稳定性比值越接近20/80, 群落就越稳定。
1.3.5 统计分析统计分析采用SPSS 21.0软件进行。采用线性回归模型拟合功能多样性-物种多样性、功能冗余-物种多样性、功能多样性-功能冗余、群落稳定性-功能冗余、群落稳定性-功能多样性以及群落稳定性-物种多样性间的关系, 以F检验显著(P < 0.05)且决定系数R2来进行比较分析。
采用二元逐步回归分析, 以群落稳定性为因变量, 以功能多样性和功能冗余为自变量, 确定功能多样性和功能冗余对稳定性的相对影响, 影响程度以标准化的偏回归系数确定。
为探讨功能多样性、功能冗余、物种多样性及群落稳定性与土壤含水量及含盐量之间的关系, 首先利用DCA(除趋势对应分析), 查看各排序轴的长度(Axis lengths), 其中最大值为1.325, 小于3, 应优先选用线性RDA模型, 但根据模型效果最优的原则, 最终选取CCA排序分析[28-29]。以上计算及分析采用R(3.2.5)软件进行分析。
在改进后的Godron稳定性测定方法中绘制平滑曲线时, 由于可能存在多种关系如线性、饱和增加及S型曲线等, 我们采用线性、对数、指数、幂函数四种模型拟合了群落中植物种类倒数百分率与累积相对盖度的关系, 以F检验显著(P < 0.05)且决定系数R2值最大的模型为最优模型。
2 结果与分析 2.1 不同水盐梯度下, 群落的功能多样性、功能冗余、物种多样性和群落稳定性不同水盐梯度下, 功能多样性、物种多样性及群落稳定性均表现为高水高盐群落>中水中盐群落>低水低盐群落。除功能冗余不同水盐梯度下无显著性差异外(P>0.05), 功能多样性、物种多样性和群落稳定性均表现为高水高盐和中水中盐群落显著高于低水低盐群落(P < 0.05)(表 4)。
| 变量 Variables |
功能多样性 Functional diversity |
功能冗余 Functional redundancy |
物种多样性 Species diversity |
群落稳定性 Community stability |
| 高水高盐 | 0.7201±0.0999b | 0.3267±0.1298a | 1.0467±0.2125b | 0.5048±0.0930b |
| 中水中盐 | 0.5763±0.1741b | 0.3300±0.1533a | 0.9063±0.2745b | 0.4351±0.02885b |
| 低水低盐 | 0.3245±0.1974a | 0.2478±0.0907a | 0.5723±0.2752a | 0.3245±0.02541a |
| 同一列相同字母表示不同水盐梯度下功能多样性、功能冗余、物种多样性及群落稳定性无显著差异(P>0.05) | ||||
线性回归分析显示(图 2), 在高水高盐梯度上, 功能多样性与物种多样性的相关性0.8131(由相关系数R2决定)小于功能冗余与物种多样性的相关系数0.8924, 功能多样性与群落稳定性的相关性0.770也小于功能冗余与群落稳定性的相关性0.8453;而中水中盐和低水低盐梯度上, 功能多样性与物种多样性的相关性大于功能冗余与物种多样性的相关性, 功能多样性与稳定性的相关性也大于功能冗余与稳定性的关系。由图 2可知, 随着水盐梯度的降低, 功能多样性与稳定性的相关系数增大, 功能冗余与高水高盐稳定性的相关性大于与中水中盐及低水低盐的相关性。不同水盐梯度下, 物种多样性和群落稳定性均呈正相关关系。

|
| 图 2 不同水盐梯度下物种多样性、功能多样性、功能冗余与群落稳定性之间的线性关系 Fig. 2 Linear relations between species diversity, functional diversity, functional redundancy, and community stability under different water-salinity habitats |
逐步回归分析结果显示(表 5), 中水中盐和低水低盐梯度, 功能多样性和功能冗余的标准化偏回归系数均大于0, 说明增加功能多样性和功能冗余均能提高群落稳定性, 决定系数R2的值大小表明能共同解释群落稳定性77.2%—80.3%的变化。高水高盐群落, 由于功能多样性的贡献性相对于功能冗余过小, 在逐步回归过程中剔除。中水中盐的功能多样性和功能冗余的标准化偏回归系数分别为:0.373和0.235;低水低盐的功能多样性和功能冗余的标准化偏回归系数分别为:0.357和0.243;可见中水中盐和低水低盐的功能多样性的标准化偏回归系数均大于功能冗余的标准化偏回归系数, 因此功能多样性对群落稳定性的影响大于功能冗余的影响。由于各逐步回归的自变量的容忍度值较大, 功能多样性和功能冗余之间不存在共线性(表 5)。
| 水盐梯度 Water and salt gradients |
回归方程 Regression equations |
R2 | F-test | P | 容忍度 Tolerance |
| 高水高盐 | y=0.292+0.652x2 | 0.801 | 24.370 | < 0.05 | 0.501 |
| 中水中盐 | y=0.143+0.373x1+0.235x2 | 0.803 | 147.014 | < 0.001 | 0.804 |
| 低水低盐 | y=0.148+0.357x1+0.243x2 | 0.772 | 332.293 | < 0.001 | 0.363 |
在CCA排序中(图 3), 主要分析整体上物种多样性、功能多样性、功能冗余及其与群落稳定性在空间上的含义, 其中各排序轴包含物种多样性、功能多样性、功能冗余及稳定性和样方水盐矩阵中所涵盖的信息量的大小, 可以通过其特征值体现。在排序过程中可以看出两条排序轴的信息量, CCA排序的第一轴特征根值占总特征根值的22.7%, 即第一轴能够解释样方与水分的相关关系, 它与土壤含水量有极显著相关关系, 说明第一轴主要反映土壤含水量对稳定性、功能多样性及功能冗余的影响, 第二轴与盐分存在相关关系, 但仅能解释1.3%, 说明土壤含水量对功能多样性、功能冗余和稳定性的影响大于土壤含盐量对其影响。由排序图也可以看出整体来讲物种多样性、功能多样性与群落稳定性的相关性较强, 功能冗余与群落稳定性的相关性较弱。

|
| 图 3 各样方的功能多样性、功能冗余及稳定性与水盐的典范对应分析(CCA)排序图 Fig. 3 Canonical correspondence analysis (CCA) ordination diagram of 30 plots and species diversity, functional diversity, functional redundancy and community stability □代表高水高盐群落;〇代表中水中盐群落;△代表低水低盐群落;W代表水分;S代表盐分;D代表物种多样性;Q代表功能多样性;FR代表功能冗余;ICV代表群落稳定性 |
从稳定性分析结果可见(表 6), 高水高盐的稳定比最接近20/80, 其稳定性最高, 低水低盐的稳定比远离20/80, 其稳定性最低, 中水中盐居于两者之间。
| 样方编号 Plots |
曲线 Curve |
决定系数R2 Coefficient of determination |
P | 交点坐标 Intersection coordinate |
| 高水高盐 | Y=14.632ln(x)+36.632 | 0.9747 | < 0.01 | (19.75, 80.25) |
| 中水中盐 | Y=-0.0279x2+3.7621x+3.2609 | 0.9680 | < 0.01 | (23.57, 76.43) |
| 低水低盐 | Y=-0.0129x2+1.9663x+30.284 | 0.8320 | < 0.01 | (26.57, 73.43) |
大量研究表明, 物种多样性对群落稳定性具有正效应[24, 29], 而物种多样性对群落稳定性的正效应是通过功能多样性和功能冗余的增加实现的, 且功能多样性是起互补效应的作用, 功能冗余是起保险效应的作用[30]。Bello等研究结果表明, 物种多样性与功能多样性和功能冗余正相关[12]。本研究为此提供了新的证据, 结果表明, 不同水盐梯度下, 物种多样性与群落稳定性正相关, 功能多样性和功能冗余对群落稳定性的影响大小不同。高水高盐条件下, 通过线性回归和逐步回归(图 2、表 5)的结果表明功能多样性对群落稳定性的影响很小, 而功能冗余对群落稳定性的影响占主导地位, 这与Pillar得出的结论群落稳定性仅由功能冗余所决定相一致[31], 可能是在现有土壤水盐条件下, 物种相对丰富, 干旱区物种多样性已达到饱和状态, 从而功能冗余对群落稳定性的影响相对来讲更大。Petchey等认为[14], 一个群落的功能冗余程度由内在的冗余和外在的冗余共同决定, 内在冗余指种间的功能相似性, 而外在冗余产生的原因是物种组成的非随机变化引起的植物功能性状的非随机变化。如骆驼等牲畜选择性的采食适口性物种以及放牧对竞争优势物种的抑制效应则会增加物种的相对多度或多样性, 引起性状组成的非随机变化, 产生外在冗余。功能冗余是群落本身的内在属性, 因此, 只有在群落物种多样性趋于饱和后, 功能冗余才会成为影响群落稳定性的主要因素。而中水中盐和低水低盐条件下, 功能多样性与物种多样性的相关性强于功能冗余与物种多样性的相关性, 说明物种间通过更强的功能互补性维持群落稳定性;功能多样性与群落稳定性的相关性也强于功能冗余与群落稳定性的相关性更进一步说明了互补效应的作用大于保险效应的作用, 这与姚天华等的研究结果一致[32], 即群落物种功能多样性和功能冗余对群落稳定性的影响中, 功能多样性对群落稳定性的影响较大。得出种间功能性状差异的互补效应才是维持群落稳定性的主要动力。综合以上结果及证据, 我们认为群落物种功能多样性和功能冗余对扰动发生响应变化的程度确实与其对群落稳定性影响的相对强弱正相关, 即功能多样性和功能冗余两者中与物种多样性关系更为密切者对群落稳定性的影响也将越大。
在本研究区, 高水高盐群落在逐步回归结果中, 功能多样性对群落稳定性的贡献很小, 而且在线性回归模型中, 功能多样性对群落稳定性的相关性小于功能冗余对群落稳定性的相关性, 我们推测可能是因为干旱荒漠区物种多样性较低, 高水高盐群落的物种多样性已经趋于饱和, 功能冗余开始发挥主要作用。但目前研究区物种饱和的精确分析还需进一步的试验研究。
3.2 CCA分析对稳定性及环境的解释CCA排序中同时具有样方的功能多样性、功能冗余、物种多样性及群落稳定性矩阵, 也有各样方的水盐矩阵, 一方面可以解释整体群落中功能多样性、功能冗余、物种多样性和稳定性的关系, 同时也可以较好的表达出各指标与水盐梯度的关系。总体而言, 样方间水盐越接近, 样方间的功能多样性、功能冗余、物种多样性及稳定性越接近, 这是群落的功能多样性、功能冗余、物种多样性及稳定性与水盐之间形成的生态关系。本CCA排序中, 因只有两个环境因子即水分和盐分含量, 第一轴与水分含量有关, 从左到右表达了水分递减的一个梯度;第二轴主要与盐分含量有关, 从下到上表达了盐分递增的梯度。综合来说, 可能是土壤含盐量的范围不足以对功能多样性、功能冗余、物种多样性及群落稳定性产生限制性的影响(表 2), 土壤含水量是影响功能多样性、功能冗余、物种多样性及稳定性的主要的环境因子。这可能与植物对土壤水分的生态适应性相关。水分胁迫可能是导致不同水盐梯度下功能多样性和功能冗余的主要原因。这与张雪妮等[33]的研究结果一致, 即艾比湖荒漠植物多样性在土壤含盐量为0.03%—1.24%的范围内植物的生长不受影响, 干旱胁迫可能是导致两种生境下植物多样性指数差异显著的主要原因。在各个样方的排序图上沿着CCA第一排序轴出现了较规律的分布, 且出现了交错的现象, 显示出群落的功能多样性、功能冗余及稳定性与其所处环境之间相互关系的复杂性。除了涉及水分和盐分差距, 可能生物间的相互作用等也会影响这些群落特征。
CCA排序中, 物种多样性与群落稳定性的相关性最大, 功能冗余与群落稳定性的相关性小于功能多样性与群落稳定性的相关性。功能冗余受水盐条件的影响较小, 可能是物种长期与环境及种群之间的相互作用, 形成了极为复杂的关系, 不易受水盐条件的影响而改变。而功能多样性对水盐变化较敏感。
3.3 改进后的Godron稳定性结果分析干旱区荒漠植物群落稳定性主要是受种间竞争、环境压力和干扰活动等的影响[27], 本研究表明, 利用改进的Godron稳定性测定方法测得的三类群落的稳定性与通过物种种群密度变异系数得到的结果一致, 可能和种间联接及物种多样性有关, 稳定性由高到低依次为:高水高盐群落>中水中盐群落>低水低盐群落。可能是高水高盐梯度土壤含盐量在植物生长所需的限度内, 水分相对充足, 高水高盐的物种多样性最高, 最适合物种的生存, 其稳定性最高;中水中盐和低水低盐梯度, 土壤含水量较低, 限制了某些物种的生长, 稳定性较低。
4 结论综上所述, 从不同水盐梯度荒漠植物群落稳定性对功能多样性和功能冗余响应的研究得出以下结论:
(1) 高水高盐梯度下, 功能多样性对群落稳定性的影响小于功能冗余对群落稳定性的影响;中水中盐和低水低盐梯度下, 功能多样性对群落稳定性的影响大于功能冗余对群落稳定性的影响。
(2) 功能多样性和功能冗余均可以提高群落稳定性。且两者中与物种多样性关系更为密切者对群落稳定性的影响也越大, 即冗余假说(冗余可以提高群落稳定性)在该干旱荒漠区域的隐域性植物群落中是成立的。
(3) 群落的稳定性、功能多样性、功能冗余及物种多样性受水盐的协同作用, 但土壤含水量大于土壤含盐量的影响。
| [1] | Elton C S. The Ecology of Invasions by Animals and Plants. Chicago: The University of Chicago Press, 2000. |
| [2] | MacArthur R. Fluctuations of animal populations and a measure of community stability. Ecology, 1955, 36(3): 533–536. DOI:10.2307/1929601 |
| [3] | Mougi A, Kondoh M. Diversity of interaction types and ecological community stability. Science, 2012, 337(6092): 349–351. DOI:10.1126/science.1220529 |
| [4] | 张景慧, 黄永梅. 生物多样性与稳定性机制研究进展. 生态学报, 2016, 36(13): 3859–3870. |
| [5] | Ehrlich P R, Ehrlich A H. Extinction. New York: Ballantine, 1981: 384. |
| [6] | Walker B H. Biodiversity and ecological redundancy. Conservation biology, 1992, 6(1): 18–23. DOI:10.1046/j.1523-1739.1992.610018.x |
| [7] | Lawton J H. What do species do in ecosystems?. Oikos, 1994, 71(3): 367–374. DOI:10.2307/3545824 |
| [8] | Sala O E, Lauenroth W K, McNaughton S J, Rusch G, Zhang X S, Mooney H A, Cushman J H, Medina E, Schulz E D. Biodiversity and ecosystem functioning in grasslands//Mooney H A, Cushman J H, Medina E, Sala O E, Schulze E D, eds. Functional Roles of Biodiversity:A Global Perspective. Chichester:Wiley, 1996:129-149. |
| [9] | Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441(7093): 629–632. DOI:10.1038/nature04742 |
| [10] | Mori A S, Furukawa T, Sasaki T. Response diversity determines the resilience of ecosystems to environmental change. Biological Reviews, 2013, 88(2): 349–364. DOI:10.1111/brv.2013.88.issue-2 |
| [11] | Schulze E D, Mooney H A. Biodiversity and Ecosystem Function. Berlin Heidelberg: Springer, 1994. |
| [12] | De Bello F, Lepš J, Lavorel S, Moretti M. Importance of species abundance for assessment of trait composition:an example based on pollinator communities. Community Ecology, 2007, 8(2): 163–170. DOI:10.1556/ComEc.8.2007.2.3 |
| [13] | 江小雷, 张卫国. 功能多样性及其研究方法. 生态学报, 2010, 30(10): 2766–2773. |
| [14] | Petchey O L, Evans K L, Fishburn I S, Gaston K J. Low functional diversity and no redundancy in British avian assemblages. Journal of Animal Ecology, 2007, 76(5): 977–985. DOI:10.1111/jae.2007.76.issue-5 |
| [15] | Elmqvist T, Folke C, Nyström M, Peterson G, Bengtsson J, Walker B, Norberg J. Response diversity, ecosystem change, and resilience. Frontiers in Ecology and the Environment, 2003, 1(9): 488–494. DOI:10.1890/1540-9295(2003)001[0488:RDECAR]2.0.CO;2 |
| [16] | Naeem S. Species redundancy and ecosystem reliability. Conservation Biology, 1998, 12(1): 39–45. DOI:10.1046/j.1523-1739.1998.96379.x |
| [17] | Thibaut L M, Connolly S R, Sweatman H P A. Diversity and stability of herbivorous fishes on coral reefs. Ecology, 2012, 93(4): 891–901. DOI:10.1890/11-1753.1 |
| [18] | Gonzalez A, Loreau M. The causes and consequences of compensatory dynamics in ecological communities. Annual Review of Ecology, Evolution, and Systematics, 2009, 40: 393–414. DOI:10.1146/annurev.ecolsys.39.110707.173349 |
| [19] | 中国科学院新疆综合考察队, 中国科学院植物研究所. 新疆植被及其利用. 北京: 科学出版社, 1978. |
| [20] | Gower J C. A general coefficient of similarity and some of its properties. Biometrics, 1971, 27(4): 857–871. DOI:10.2307/2528823 |
| [21] | Podani J. Extending Gower's general coefficient of similarity to ordinal characters. Taxon, 1999, 48(2): 331–340. DOI:10.2307/1224438 |
| [22] | Ricotta C. A note on functional diversity measures. Basic and Applied Ecology, 2005, 6(5): 479–486. DOI:10.1016/j.baae.2005.02.008 |
| [23] | Zhang J T. Quantitative Ecology. Beijing: Science Press, 2004. |
| [24] | Yang Z L, Van Ruijven J, Du G Z. The effects of long-term fertilization on the temporal stability of alpine meadow communities. Plant and Soil. 2011: 315–324. |
| [25] | Sasaki T, Lauenroth W K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oeclogia, 2011, 166(3): 761–768. DOI:10.1007/s00442-011-1916-1 |
| [26] | 王海东, 张璐璐, 朱志红. 刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制. 植物生态学报, 2013, 37(4): 279–295. |
| [27] | 吕光辉, 杜昕, 杨建军, 马媛, 孟吉翔. 阜康绿洲——荒漠交错带荒漠植被群落稳定性. 干旱区地理, 2007, 30(5): 660–665. |
| [28] | 国际生物多样性计划中国委员会. 中国生物多样性保护与研究进展. 北京: 气象出版社, 2014. |
| [29] | 张钦弟, 段晓梅, 白玉芳, 王祎玲, 张金屯. 山西太岳山脱皮榆群落的功能多样性. 植物学报, 2016, 51(2): 218–225. DOI:10.11983/CBB15033 |
| [30] | Tilman D. The ecological consequences of changes in biodiversity:a search for general principles. Ecology, 1999, 80(5): 1455–1474. |
| [31] | Pillar V D, Blanco C C, Müller S C, Sosinski E E, Joner F, Duarte L D S. Functional redundancy and stability in plant communities. Journal of Vegetation Science, 2013, 24(5): 963–974. DOI:10.1111/jvs.12047 |
| [32] | 姚天华, 朱志红, 李英年, 潘石玉, 孔彬彬, 卫欣华, 杜家丽. 功能多样性和功能冗余对高寒草甸群落稳定性的影响. 生态学报, 2016, 36(6): 1547–1558. |
| [33] | 张雪妮, 吕光辉, 杨晓东, 贡璐, 秦璐, 何学敏, 刘昊奇. 基于盐分梯度的荒漠植物多样性与群落、种间联接响应. 生态学报, 2013, 33(18): 5714–5722. |