 2017, Vol. 37
2017, Vol. 37文章信息
- 宋文琛, 同小娟, 李俊, 张劲松.
- SONG Wenchen, TONG Xiaojuan, LI Jun, ZHANG Jinsong.
- 三源区分土壤呼吸组分研究
- Studies on three-source partitioning of soil respiration
- 生态学报. 2017, 37(22): 7387-7396
- Acta Ecologica Sinica. 2017, 37(22): 7387-7396
- http://dx.doi.org/10.5846/stxb201609201887
-
文章历史
- 收稿日期: 2016-09-20
- 修订日期: 2017-01-16





2. 中国科学院地理科学与资源研究所 陆地水循环及地表过程重点实验室, 北京 100101;
3. 中国林业科学研究院林业研究所 国家林业局林木培育重点实验室, 北京 100091
2. Key Laboratory of Water Cycle and Related Land Surface Processes, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;
3. Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration, Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, China
大气中二氧化碳等温室气体升高所引起的温室效应是多年来生态学研究的热点[1-2],全球每年约有超过120Pg C通过光合作用从大气中被固定[1]。其中约一半以植物呼吸的形式又释放回大气,剩下的另一半以凋落物的形式进入土壤,再以土壤呼吸的形式释放到大气中,由此循环于陆地生态系统与大气之间[1, 3]。作为全球碳循环重要一环的土壤呼吸,一旦产生微小变动,就能对全球碳循环产生显著影响[1, 4]。其中,根际的植物-土壤作用过程调控着全球陆地生态系统近一半的碳释放量[5-6],并且陆地生态系统的多个物质循环过程都受其影响[1, 7]。因此,区分由根系引发的土壤呼吸组分也成为研究全球变化的重要领域[1]。
土壤有机质的分解主要受土壤有机碳含量、土温和土壤含水率等环境因子控制[1-2, 8]。因此,土壤呼吸通常被分成“根呼吸”(自养呼吸组分)和“土壤有机质呼吸”(异养呼吸组分)两部分[1]。然而,土壤存在着被称为“微生物热点”(microbial hotspots)的微生物活跃度异常高于其他区域的地方[9]。随着对各个微生物热点的深入研究,人们发现土壤激发效应是影响土壤有机质分解的重要因素,其在各种不同类型的生态系统中都普遍存在[10]。土壤激发效应(soil priming effect)是指由各种有机物质添加等处理所引起的土壤有机质释放和固定平衡的强烈改变,主要表现为有机质碳释放速率的突然变化[1, 10]。根系附近3 mm内(也就是根际)是激发效应最主要的发生部位[5, 10],而根系附近的微生物热点则可以扩展至根附近10 mm[9]。尽管当前对激发效应的研究已取得一定的进展,但是对激发效应的生态学重要功能依然缺乏足够的了解[1, 11]。在这种背景下,Kuzyakov[12]将土壤呼吸组分分为5个部分,即:根呼吸、根际微生物呼吸、枯落物分解、激发的土壤有机质CO2释放和土壤有机质呼吸[1]。这五部分中最主要的3个组分是根呼吸、根际微生物呼吸和土壤有机质呼吸[1, 13]。传统意义上的“根呼吸”应当被称为“根源呼吸”(rhizosphere respiration),它还可以进一步区分为(纯)根呼吸(root respiration)和根际微生物呼吸(rhizomicrobial respiration)两部分[1, 12]。根呼吸和根际微生物呼吸都是由根系引发的自养呼吸,均发生在根际;土壤有机质呼吸则是由微生物分解土壤有机质产生的异养呼吸[1, 14]。这种区分的意义主要体现在6个方面[1], 即土壤和植物中的碳平衡的评估;对有机物质中根际土壤微生物量的评估;对腐殖质的完全统计;区分自养微生物和异养微生物;研究根际激发效应的机制;生态系统碳循环的模型[1, 14]。区分根源呼吸最主要的意义就是纠正因忽略根际激发效应而引起的误差,而区分根际激发效应的自养和异养组分则是更重要而且有难度的问题[1, 15]。本文结合文献资料,综合论述三源区分土壤呼吸组分的意义、方法和应用,为今后土壤碳循环研究提供参考。
1 三源区分土壤呼吸组分的意义总土壤呼吸区分成植物根(源)呼吸和土壤有机质呼吸的意义主要有以下几个方面。首先,根源呼吸分解的是植物输送到地下的光合产物,不应当被计入土壤微生物分解有机质所释放的CO2,由此土壤中碳固定可用:净土壤碳增量=凋落物输入量-(土壤呼吸-根源呼吸)来表示[1-2, 16]。若不能精确区分出根源呼吸,则会影响土壤碳固定量的估算,进一步影响土壤碳源或碳汇的估算[1-2]。其次,通常在计算碳素周转率时以土壤有机质分解的二氧化碳量衡量土壤有机质分解量[1]。如果不对所测得土壤呼吸进行区分,势必影响计算[1, 16-17]。最后,根源呼吸基本属自养呼吸,土壤有机质呼吸属于异养呼吸,两者对各种环境变化的响应存在一定的区别[1]。因此,在估算土壤碳存储潜力及模型模拟时则必须对两者加以区分[1, 18]。
在是否应当区分根源呼吸的问题上,目前还存在着一定争议[1, 15]。根源呼吸实际上是把植物通过光合作用吸收二氧化碳所转化的有机碳再转化为二氧化碳,把植物固定的能量再释放出去[1]。这一过程对大气碳平衡没有实质性的影响[1, 19]。正因为(纯)根呼吸和根际微生物呼吸同属自养呼吸,所以很多研究者并不认为有必要将两者区分开[1, 15]。但若不对以上两者进行区分,通常会令研究者忽略根际微生物的呼吸作用,也就是根际异养微生物分解植物光合产物所造成自养呼吸的作用,从而造成了研究的不准确[1, 20]。尽管根际微生物呼吸和激发的土壤有机质CO2释放都发生在根际,且都由异养微生物受根系激发而产生,但根际微生物呼吸属于自养呼吸,激发的土壤有机质CO2释放则来自于根际微生物分泌胞外酶分解土壤有机质而产生,属于异养呼吸,应当被归类于土壤有机质呼吸[12, 21]。实际上,根际微生物产生的CO2一部分来自自养呼吸,一部分来自异养呼吸[1]。利用根际物质和利用土壤有机质的土壤微生物并没有明显界限,它们会随营养状况不同而转化[1, 22]。不区分根源呼吸组分不利于了解土壤碳循环过程,造成对根际激发效应的忽略,使得生态学模型的精度下降、不确定性增高[14]。本文认为出现这种争议的主要原因是研究对象和尺度的不同。对于大尺度景观或全球生态学的研究,人们只关心自养和异养组分的贡献,而不关心土壤有机碳的迁移过程和机理,因此,并不需要区分根源呼吸组分[23]。但对于土壤基础研究,人们更关心土壤有机碳的迁移过程和机理以了解碳从根到土壤再到释放的过程。因此,对这类研究来说,很有必要区分根源呼吸[9]。
一些模型模拟研究认为,随着全地球大气二氧化碳浓度和温度升高,植物生物量和土壤有机碳库中的碳(C)蓄积量也随之提高[24],同时也会增加根际分泌物和(地表)凋落物向土壤的输入量[25-26],土壤有机质受到激发而分解加速,从而对大气二氧化碳浓度和气温升高产生促进作用[1, 10, 27-28]。然而,Kuzyakov和Larionova[20]指出激发效应可以促进以根际微生物为主异养生物的自养呼吸[1]。Yuan等[29]认为微生物自养呼吸是造成碳失汇的重要原因[1]。事实上,激发效应究竟能否影响所研究的土壤碳释放的估计也存在着争议[1, 30-31]。因此,综合估计土壤呼吸各组分的贡献对改进陆地生态系统碳循环模型,准确评估全球变化对陆地生态系统具有深远的影响[1]。
2 区分根源呼吸和土壤有机质呼吸的方法区分根源呼吸和土壤有机质呼吸的方法主要有根分离法、间接模拟法、同位素长期标记和同位素短期标记(表 1)[12, 32-33]。在这些区分方法中,最精准的是14C动态标记法,它甚至可以精准研究土壤呼吸不同组分动态变化[34-35]。最常用的方法是简单方便的壕沟法,其原理是在地块四周挖沟切断根系和真菌菌丝,并埋设物理薄板阻隔根系生长,隔离地块内外土壤呼吸差值即根源呼吸[33]。然而,不同方法区分出的土壤呼吸差异很大,这造成了很强的不确定性[36]。鉴于同位素标记法通常更为准确,在既有条件用同位素法和又能用非同位素法时,应当选择更精确的同位素标记法[35]。
| 方法Method | 原理Principle | 优点Advantages | 缺点Disadvantages |
| 根分离法 Root excised method |
将根呼吸和土壤有机质呼吸分别测定, 土壤呼吸视为两者之和;或对比有根呼吸和无根呼吸地块, 根呼吸视为两者之差 | 原理简单, 成本低, 适宜各种生态系统 | 扰动大、结果不准确、费人工 |
| 间接模拟法 Indirect Modeling |
用生态学模型推断出土壤呼吸各组分 | 方法简单易行、成本低 | 精确度不高, 不适于精确研究 |
| 同位素长期标记 Isotopic tracer continuous labeling |
用自然(比如C3、C4光合途径)或人为(比如上世纪核弹爆炸制造的14C)过程导致的长期均匀的同位素标记区分土壤呼吸各组分 | 省人工, 精确度较高 | 存在同位素分馏问题, 只适用于特定条件下, 成本较高 |
| 同位素短期标记 Isotopic tracer pulse labeling |
人工添加同位素标记并根据其丰度变异计算土壤呼吸各组分 | 结果精准, 适合实验室控制试验 | 成本高, 不适用于野外和长期观测 |
与同位素法相比,非同位素法很难区分纯根呼吸和根际微生物呼吸[1, 37]。于是研究者总结非同位素方法的优缺点,把各种方法综合起来,用以分离纯根呼吸和根际微生物呼吸。如,Kelting等[38]将离体根法和壕沟法结合起来,该研究基于两个假设:1)离体根法中,切除根后迅速测定(一般认为2 h之内)不会影响根呼吸速率;2)壕沟法中,样地内的死根不影响土壤呼吸的测定[1]。两个实验同时进行:1)离体根法中,将根分离出来后迅速测定根呼吸速率,认为测定结果就是纯根呼吸速率;2)壕沟法中,在样地周围挖一圈壕沟,切断根并阻止外根进入[1]。经常对样地表层进行清理,避免新根生长。待碳释放稳定后(残留根完全分解后)测定此时土壤呼吸[1]。原土壤呼吸减去此时测定的土壤呼吸就是“根呼吸”,以这个结果作为根源呼吸值(即纯根呼吸速率与根际微生物呼吸速率之和)[1]。两者之差即为根际微生物呼吸速率[1]。Chen等[37]用类似的原理在实验室条件下用离体根法结合移除根法分别区分了黑麦草(Lolium perenne L.)和辐射松(Pinus radiata D. Don)的根源呼吸[1]。Craine等[40]通过对叶片遮阴处理,发现土壤呼吸的下降量相当于纯根呼吸量[1]。由此再结合空白对照组的呼吸速率即可区分出根际微生物呼吸[1]。Larionova等[41]将成分综合法和移除根法结合起来[1]。方法是:1)先将根分离出来在实验室里培养(培养基质最好是砂或溶液),再测定根呼吸速率,认为该值为纯根呼吸速率;2)用移除根法实验测定“根呼吸”速率值,以该值作为纯根呼吸速率与根际微生物呼吸速率之和;3两者之差即为根际微生物呼吸速率[1]。Barba等[42]设计了3种PVC管:第一种保留所有根与菌丝;第二种保留菌丝但没有根;第三种既没有根也没有菌丝,通过对比3种PVC管的土壤呼吸值来判定土壤呼吸3种组分的呼吸值。
3.2 14C脉冲标记法14C脉冲标记法是指对植物进行14C标记后,追踪14CO2在土壤中的释放,通过研究植物碳的瞬时动态变化,进而区分纯根呼吸和根际微生物呼吸的方法[1]。它可以分为14C稀释法、模拟根际分泌物法、根际分泌物洗脱法和14CO2动态法[1, 37]。
14C稀释法是指将未标记的葡萄糖注入到生长有14C标记植物的土壤中[1]。由于根际微生物不光利用根际分泌物,还利用葡萄糖,这等于稀释了有14C标记的根际分泌物。利用稀释14C所造成的同位素标记变化,就可估算出纯根呼吸[1]。该法基于几个假设,即添加含14C标记的葡萄糖只对根际分泌物起稀释作用而对植物生理、分泌物分解,根际微生物活性等不产生其他影响[1]。假定稀释作用与葡萄糖添加量成正比[43],得出方程[1, 37]:
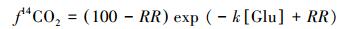
|
(1) |
式中,f14CO2为葡萄糖稀释后14CO2的放出量与未添加葡萄糖时14CO2放出量的百分比,RR为纯根呼吸,Glu为加入的葡萄糖量,k为稀释14C的比例[1]。需要注意的是,添加含14C葡萄糖不仅对纯根呼吸有影响,而且对微生物呼吸也有影响[44],假设不准确严重影响了实验精度[1]。
模拟根际分泌物法假设土壤微生物释放的碳与土壤碳释放总量之比恒定,则可分两组实验:一组, 向土壤中添加有14C标记的模拟分泌物,而植物不标记;另一组,用14C标记土壤上生长的植物,而不标记土壤[1]。由此,得出如下方程[37]:
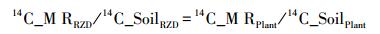
|
(2) |
式中,14C_MRRZD和14C_MRPlant表示土壤微生物分解释放的14C,14C_SoilRZD与14C_SoilPlant表示存留在土壤中的14C[1]。该法实际上存在一个潜在假设,即:土壤微生物对根际14C标记物的吸收与无根土壤是相同的[1]。然而,该假设并不精确,从而与实际值不符[1, 37]。
根际分泌物洗脱法的操作是:先进行14C标记,然后同时进行两样操作,一方面用持续的气流将纯根呼吸释放的CO2吹出并收集;另一方面用持续的水流将根周围的分泌物等物质一齐冲出并收集,最后对所收集物质进行分析检验[1]。需要注意的是,取样时间的选定是一个重点和难点[1]。Kuzyakov[37]发现,吹出的14C峰值出现在标记后第12小时,而冲出的分泌物等物质的14C峰值分别出现在洗涤后第5小时和第20—24小时[1]。根际分泌物洗脱法的最大优点在于它直接分离了两种呼吸,而不需要方程推算[1]。但是它的缺点也很多:(1)洗涤并不完全;(2)洗涤并不均匀;(3)破坏原来根系环境[1]。这些缺点很容易造成根呼吸的14C计算偏高而根际微生物的14C计算偏低[1, 37]。
14CO2动态法主要利用的是根际微生物相对于纯根呼吸在碳释放方面的延迟[1]。也就是说在最初标定时14CO2基本来自纯根呼吸,而大约2至5天时根际微生物呼吸才有所反映[1]。这是因为第二个过程还包括几个阶段:(1)分泌物从根向根际的释放;(2)微生物对分泌物的消耗;(3)微生物分解的分泌物释放14CO2[1, 14]。这些过程的耗时使得根际微生物呼吸的响应有所延迟[1]。将14CO2流区分至土壤呼吸各组分依照的是根际碳通量的动态模型(包括适当标记后的根呼吸率和根分泌物分解率)[1, 45]。这些模型参量需符合土壤14CO2释放的经验数据[1]。获取数据后,采用区分模型模拟纯根呼吸和微生物呼吸的动态过程,进而确定出二者之间的比值。研究表明,14CO2动态法精度高,可以作为区分纯根呼吸和根际微生物呼吸的标准验证方法[1, 46]。
3.3 自然13C丰度法自然13C丰度法正是利用了根系、土壤、凋落物具有不同的δ13C值,据此差别估计各来源的贡献率[1]。通常的做法是,将C3植物种植在原生长C4植物的土壤上(或C4植物种植在原生长C3植物的土壤上)[1]。原理是C3和C4植物土壤具有不同的δ13C值,C3植物δ13C约为-24‰~—29‰,C4植物约为-12‰~—14‰,两者大约相差14‰[1, 47]。基于此,根据二元混合模型(Linear two components mixing model)根源呼吸占土壤总呼吸的比重(fr)可以用下式表示:
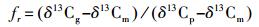
|
(3) |
式中,δ13Cg是土壤CO2的δ13C值;δ13Cp是现生长植物呼出的δ13C值;δ13Cm是土壤有机物分解呼出的δ13C值[1]。
在上述方法的基础上,Kuzyakov[48]结合微生物生物量的区分方法,建立了进一步区分纯根呼吸和根际微生物呼吸的方法。该法基于两个假设:(1)纯根呼吸的δ13C与根系组织的δ13C差异不大;(2)微生物呼吸的δ13C与微生物生物量δ13C相关联[1, 13]。计算步骤为:第一步与稳定同位素法一致,区分根源呼吸和土壤有机质分解呼吸;第二步区分微生物生物量;第三、第四步利用两个假设推导计算得出结论[1, 48]。
以将C4植物种在原本生长C3植物的土壤上为例(相反也可以)[1]。理论上,根据此方法只需测量四个参量就能计算出结果[1]。Kuzyakov[48]依据以上步骤推得计算公式:

|
(4) |
式中,RR表示纯根呼吸所占根源呼吸比重;RMR表示根际微生物呼吸所占根源呼吸比重;δCO2表示土壤总呼吸的δ13C值;δ3SOM表示曾生长C3植物土壤有机质分解的δ13C值;δ4Rhiz表示C4植物根源呼吸的δ13C值;δMO表示土壤微生物生物量δ13C值[1]。
Werth等[49]则认为,根际微生物活性过低会导致实验失败。实验室条件下分解土壤有机质的微生物活性很弱,使得土壤有机质分解时出现δ13C的分馏,在这种情况下上述前提假设(2)不成立,所以得出的结论会严重高估纯根呼吸而严重低估微生物呼吸[1]。Werth和Kuzyakov[13]在考虑分馏影响后,在种植玉米(Zea mays L.)的农田成功将这个方法应用于野外研究[1]。若条件允许,也可以通过添加适量13C标记提高精度[50-51]。宋文琛等[52]、Song等[53]和Tomè等[54]将自然13C丰度法原理用于区分人工林根源呼吸组分,并认为该方法适用于森林生态系统。自然13C丰度法目前还处于探索阶段,还需要进一步的完善[1, 52-53, 55]。
4 三源区分土壤呼吸组分的应用 4.1 根源呼吸对土壤有机质呼吸的影响根际微生物呼吸部分是根际激发效应所产生的自养呼吸,而根际激发效应同时还会增加土壤有机质呼吸的强度[1]。因此,区分出根际微生物呼吸并研究其大小和变化有助于精确衡量根际激发效应的作用[10, 50]。Kerré等[51]通过添加不同物质,利用13C丰度法原理不仅得出根际激发效应强度随时间变化,而且得出生物炭添加引起的土壤有机质呼吸增强程度随时间变化。Tian等[56]通过用同位素法三源区分土壤呼吸组分,发现低粒径土壤、更多的有机物添加会刺激土壤有机质的分解。研究结果表明,在植物-微生物系统中,土壤有机质的分解主要与土壤氮的有效性有关[21, 57-58]。由于植物与土壤微生物存在激烈的竞争,根际往往成为氮受限的区域[22]。在氮受限的环境中, 植物会将较多的光合产物投资到地下,促进微生物的生长和活性,加速土壤有机质分解释放无机氮,以获取所需的氮和其他养分[21, 26, 59]。因此,土壤氮含量的增加会抑制根际微生物呼吸,进而抑制土壤有机质呼吸强度[60-61]。另外,在温度、水分等发生变化时,植物-微生物系统会适时调整利用有机物的策略,造成根际微生物呼吸和土壤有机质呼吸的弹性变化[42, 52, 54]。
4.2 野外条件下三源区分土壤呼吸组分的应用由于在野外条件下三源区分土壤呼吸组分难度较大,所以相对于传统的二源区分,三源区分土壤呼吸组分的研究相对较少[1]。在野外条件下,农田和人工林的根呼吸、根际微生物呼吸和土壤有机质呼吸占总呼吸比重在生长季不同时期具有明显的变化[1, 13, 52, 54]。在农田、人工林植物生长较旺盛的时期,根际微生物呼吸占根源呼吸的35%—45%[1]。在生长季末期,人工林根际微生物占根源呼吸比重则超过70%[13, 52-53]。这种现象可能是树木根际微生物比农田活跃造成的[52],也可能是与生长季末期根际微生物不活跃导致13C分馏值改变而造成的误差有关[13, 52]。
农田和森林土壤的环境条件可以影响纯根呼吸、根际微生物呼吸和土壤有机质呼吸的动态变化[1]。Barba等[42]发现干旱会抑制森林土壤根际微生物呼吸和土壤有机质呼吸,进而使总呼吸下降。Tian等[56]通过三源区分土壤呼吸组分,发现土壤粒径越小越有利于玉米根际有机质分解,进而促进土壤有机质呼吸。Song等[53]发现根际微生物呼吸占土壤呼吸比重在40—50 cm处基本保持着20%左右的贡献,而在其他区间的贡献平均只有10%,而纯根呼吸占土壤呼吸比重则在≤50 cm土深时变化不大,大约为10%的贡献,但在>50 cm土深时则有大幅增加的趋势。另外,传统方法通常忽略了激发效应、以及根际微生物呼吸的影响,不能反映土壤深度变化对土壤呼吸的影响,因而很可能大大低估了华北人工林土壤的自养呼吸强度[53]。在苹果园的研究表明纯根呼吸和根际微生物呼吸分别占总呼吸的12%和11%,两者大体相当,因此,根际微生物呼吸不可忽略[54]。
5 不同研究的比较由表 2可见,不同方法纯根呼吸所占根源呼吸比重为23%—81%,根际微生物呼吸则占17%—77%。Sapronov等[46]通过对成分综合法、去除根法和14CO2脉冲标记法的对比分析得出:土壤及根际微生物所占总呼吸的18%—50%;纯根呼吸占总呼吸的8%—32%;根际微生物所释放的碳占了土壤微生物总呼吸的50%—80%。研究结果的差异主要来自于实验所采用的方法以及土壤有机物的含量[1, 46]。目前,大部分的实验还只是停留在室内实验水平上,实验材料以草本植物为主(表 1)[1]。通过分析他人研究成果,Werth和Kuzyakov[13]指出实验室条件下草本植物的纯根和根际微生物呼吸各占根源呼吸的50%;野外条件下草本植物的纯根和根际微生物呼吸占根源呼吸比重分别为56%和44%[1]。然而,有研究认为野外条件下纯根和根际微生物呼吸占根源呼吸比重与植物物种的关系并不明显[52]。通过对比不同方法的实验效果,在实验室条件下14CO2动态法表现出其他区分方法难以企及的精确度,可以作为标准方法[37, 46]。在野外标准条件下,无论用什么方法所得到的纯根和根际微生物呼吸占根源呼吸比重均为60%(纯根呼吸)和40%(根际微生物呼吸)(表 2)[1]。综合考虑方法的可靠性、试验条件、研究结论等,在实验室条件下,纯根和根际微生物呼吸占根源呼吸比重在标准状况下约为45%(纯根呼吸)和55%(根际微生物呼吸);野外条件下,通常两者约占根源呼吸的60%(纯根呼吸)和40%(根际微生物呼吸)(表 2)[1]。
| 植物种类 Species |
研究条件 Site |
区分方法 Methods |
fRMR | fRR | 参考文献 References |
| 辐射松Pinups radiata D.Don | 实验室 | 离体根法+移除根法 | 31—55 | 45—69 | [39] |
| 黑麦草Lolium perenne L | 实验室 | 离体根法+移除根法 | 35—51 | 49—65 | |
| 玉米Zea mays L | 实验室 | 成分综合法+移除根法 | 40 | 60 | [41] |
| 春大麦Hordeum vulgare L | 实验室 | 成分综合法+移除根法 | 39 | 61 | |
| 荞麦Fagopyrum esculentum L | 实验室 | 成分综合法+移除根法 | 77 | 23 | |
| 黑麦草Lolium perenne L | 实验室 | 14C稀释法 | 63 | 37 | [37] |
| 黑麦草Lolium perenne L | 实验室 | 模拟分泌物法 | 17—29 | 71—83 | |
| 黑麦草Lolium perenne L | 实验室 | 14CO2动态法 | 55 | 45 | |
| 黑麦草Lolium perenne L | 实验室 | 根际分泌物洗脱法 | 19 | 81 | |
| 玉米Zea mays L | 实验室 | 14CO2动态法+成分综合法 | 56 | 44 | [46] |
| 玉米Zea mays L | 实验室 | 14CO2动态法+成分综合法 | 56 | 44 | |
| 玉米Zea mays L | 实验室 | 13C标记法 | 35—55 | 45—65 | [52] |
| 玉米Zea mays L | 野外 | 自然13C丰度法 | 36 | 64 | [13] |
| 小须芒草Schizachyrium scoparium (Michx.) Nash | 野外 | 离体根法+遮荫法 | 52 | 48 | [40] |
| 美国红橡木Quercus rubra L | 野外 | 离体根法+壕沟法 | 38 | 62 | [38] |
| 刺槐Robinia pseudoacacia L | 野外 | 自然13C丰度法 | 57 | 43 | [52] |
| 刺槐Robinia pseudoacacia L | 野外 | 自然13C丰度法 | 48 | 52 | |
| 刺槐Robinia pseudoacacia L | 野外 | 自然13C丰度法 | 73 | 27 | |
| 刺槐Robinia pseudoacacia L | 野外 | 自然13C丰度法 | 39 | 61 | [53] |
| 苹果Malus pumila Mill. | 野外 | 自然13C丰度法 | 49 | 51 | [54] |
| fRMR代表根际微生物呼吸占根源呼吸比重;fRR代表纯根呼吸占根源呼吸比重 | |||||
目前,在三源区分土壤呼吸组分的研究领域内,多数研究都局限在实验室条件下,而对此项研究最需要的生态学方面(尤其是全球变化生态学)却难以将土壤学的研究成果应用于野外实践[1]。应用同位素法区分纯根和根际微生物呼吸可以解决扰动生境的问题,做到原位测量[1, 62]。但是,大部分同位素区分方法都需要人为同位素标记,这种脉冲标记不但昂贵而且标记的时空尺度很小,对于森林生态系统基本不起作用[1, 63]。由此可见,同位素法区分野外木本植物根源呼吸组分一直是本领域的重点和难点[1]。
未来三源区分土壤呼吸组分的研究可以有以下几个方面的应用:(1)研究根际激发效应过程中的碳平衡与养分平衡,综合评价根际过程中植物和微生物各自的收支[1]。尽管植物和微生物有相似的养分需求,但两者的限制因子还是存在一定的差异,前者更多地受养分限制,后者则主要受碳和能源的限制[1, 37]。只有将根系和微生物的效应区分开来才能定量评价植物—土壤系统的碳收支状况[1]。(2)探明根际来源的碳在土壤中的去向[1]。引发根际激发效应的这部分碳并没有完全被微生物分解释放到大气中,有一部分碳还是残留在土壤中[64],而这部分碳在土壤中的去向至关重要[1, 21]。(3)研究根际激发效应的微生物学机制[1]。究竟有多少微生物参与了根际激发效应?有关研究目前依然缺乏充分的实验数据[1]。(4)土壤有机质分解对温度的敏感性是预测陆地生态系统碳循环对全球变暖影响的关键环节,但目前的研究尚未达成一致的认识[1, 30-31]。根际效应的敏感性在全球变化背景下有重要地位[5],需要大量研究去破解其内在的机制,为生态系统过程模型的精确模拟预测提供基础[1]。(5)森林生态系统是最大、最稳定的陆地碳汇,对全球碳平衡有举足轻重的作用[65]。然而,野外区分木本植物根源呼吸组分无论从方法上还是应用上都存在一些未克服的问题[1]。因此,该方向是未来三源区分土壤呼吸研究的重点[1, 53]。
| [1] | 宋文琛. 自然13C丰度法区分剌槐人工林土壤呼吸三组分的研究[D]. 北京: 北京林业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10022-1015319607.htm |
| [2] | 刘佳, 同小娟, 张劲松, 孟平, 李俊, 郑宁. 太阳辐射对黄河小浪底人工混交林净生态系统碳交换的影响. 生态学报, 2014, 34(8): 2118–2127. |
| [3] | Le Quéré C, Peters G P, Andrew RM, Boden T A, Ciais P, Friedlingstein P, Houghton R A, Marland G, Moriarty R, Sitch S, Tans P, Arneth A, Arvanitis A, Bakker D C E, Bopp L, Canadell J G, Chini L P, Doney S C, Harper A, Harris I, House J I, Jain A K, Jones S D, Kato E, Keeling R F, Goldewijk K, Körtzinger A, Koven C, Lefèvre N, Maignan F, Omar A, Ono T, Park G H, Pfeil B, Poulter B, Raupach M R, Regnier P, Rödenbeck C, Saito S, Schwinger J, Segschneider J, Stocker B, Tilbrook B, Van Heuven S M A C, Viovy N, Wanninkhof R, Wiltshire A, Zaehle S, Yue C. Global carbon budget 2013. Earth System Science Data, 2014, 6(7): 235–263. |
| [4] | Atarashi-Andoh M, Koarashi J, Ishizuka S, Hirai K. Seasonal patterns and control factors of CO2 effluxes from surface litter, soil organic carbon, and root-derived carbon estimated using radiocarbon signatures. Agricultural and Forest Meteorology, 2012, 152: 149–158. DOI:10.1016/j.agrformet.2011.09.015 |
| [5] | Cheng W X, Parton W J, Gonzalez-Meler MA, Phillips R, Asao S, McNickle G G, Brzostek E, Jastrow J D. Synthesis and modeling perspectives of rhizosphere priming. New Phytologist, 2014, 201(1): 31–44. DOI:10.1111/nph.12440 |
| [6] | Hopkins F, Gonzalez-Meler M A, Flower C E, Lynch D J, Czimczik C, Tang J W, Subke J A. Ecosystem-level controls on root-rhizosphere respiration. New Phytologist, 2013, 199(2): 339–351. DOI:10.1111/nph.12271 |
| [7] | Chapin F S Ⅲ, Matson P A, Vitousek P M. Principles of Terrestrial Ecosystem Ecology.2nd ed. New York: Springer Press, 2012. |
| [8] | Kuzyakov Y, Friedel J K, Stahr K. Review of mechanisms and quantification of priming effects. Soil Biology and Biochemistry, 2000, 32(11-12): 1485–1498. DOI:10.1016/S0038-0717(00)00084-5 |
| [9] | Kuzyakov Y, Blagodatskaya E V. Microbial hotspots and hot moments in soil:Concept & review. Soil Biology and Biochemistry, 2015, 83: 184–199. DOI:10.1016/j.soilbio.2015.01.025 |
| [10] | Kuzyakov Y. Priming effects:interactions between living and dead organic matter. Soil Biology and Biochemistry, 2010, 42(9): 1363–1371. DOI:10.1016/j.soilbio.2010.04.003 |
| [11] | Blagodatskaya E V, Kuzyakov Y. Active microorganisms in soil:critical review of estimation criteria and approaches. Soil Biology and Biochemistry, 2013, 67: 192–211. DOI:10.1016/j.soilbio.2013.08.024 |
| [12] | Kuzyakov Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biology and Biochemistry, 2006, 38(3): 425–448. DOI:10.1016/j.soilbio.2005.08.020 |
| [13] | Werth M, Kuzyakov Y. Three-source partitioning of CO2 efflux from maize field soil by 13C natural abundance. Journal of Plant Nutrition and Soil Science, 2009, 172(4): 487–499. DOI:10.1002/jpln.v172:4 |
| [14] | Kuzyakov Y V, Larionova A A. Contribution of rhizomicrobial and root respiration to the CO2 emission from soil (A Review). Eurasian Soil Science, 2006, 39(7): 753–764. DOI:10.1134/S106422930607009X |
| [15] | 金钊, 董云社, 齐玉春. 区分纯根呼吸和根际微生物呼吸的争议. 土壤, 2008, 40(4): 517–522. |
| [16] | Hanson P J, Edwards N T, Garten C T, Andrews J A. Separating root and soil microbial contributions to soil respiration:A review of methods and observations. Biogeochemistry, 2000, 48(1): 115–146. DOI:10.1023/A:1006244819642 |
| [17] | Anderson J M. The effects of climate change on decomposition processes in grassland and coniferous forests. Ecological Applications, 1991, 1(3): 326–347. DOI:10.2307/1941761 |
| [18] | Li P, Yang Y H, Fang J Y. Variations of root and heterotrophic respiration along environmental gradients in Chinas forests. Journal of Plant Ecology, 2013, 6(5): 358–367. DOI:10.1093/jpe/rtt009 |
| [19] | Högberg P, Buchmann N, Read D J. Comments on Yakov Kuzyakov's review 'Sources of CO2 efflux from soil and review of partitioning methods':. Soil Biology and Biochemistry, 2006, 38(9): 2997–2998. DOI:10.1016/j.soilbio.2006.04.001 |
| [20] | Kuzyakov Y, Larionova A A. Root and rhizomicrobial respiration:A review of approaches to estimate respiration by autotrophic and heterotrophic organisms in soil. Journal of Plant Nutrition and Soil Science, 2005, 168(4): 503–520. DOI:10.1002/(ISSN)1522-2624 |
| [21] | 孙悦, 徐兴良, KuzyakovY. 根际激发效应的发生机制及其生态重要性. 植物生态学报, 2014, 38(1): 62–75. |
| [22] | Kuzyakov Y, Xu X L. Competition between roots and microorganisms for nitrogen:mechanisms and ecological relevance. New Phytologist, 2013, 198(3): 656–669. DOI:10.1111/nph.12235 |
| [23] | Xu M, Shang H. Contribution of soil respiration to the global carbon equation. Journal of Plant Physiology, 2016, 203: 16–28. DOI:10.1016/j.jplph.2016.08.007 |
| [24] | Gerber S, Joos F, Prentice I C. Sensitivity of a dynamic global vegetation model to climate and atmospheric CO2. Global Change Biology, 2004, 10(8): 1223–1239. DOI:10.1111/gcb.2004.10.issue-8 |
| [25] | Macdonald C A, Anderson I C, Bardgett R D, Singh B K. Role of nitrogen in carbon mitigation in forest ecosystems. Current Opinion in Environmental Sustainability, 2011, 3(5): 303–310. DOI:10.1016/j.cosust.2011.08.013 |
| [26] | Phillips R P, Finzi A C, Bernhardt E S. Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation. Ecology Letters, 2011, 14(2): 187–194. DOI:10.1111/j.1461-0248.2010.01570.x |
| [27] | Sayer E J, Heard M S, Grant H K, Marthews T R, Tanner E V J. Soil carbon release enhanced by increased tropical forest litter fall. Nature Climate Change, 2011, 1(6): 304–307. DOI:10.1038/nclimate1190 |
| [28] | Bengtson P, Barker J, Grayston S J. Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects. Ecology and Evolution, 2012, 2(8): 1843–1852. DOI:10.1002/ece3.311 |
| [29] | Yuan H Z, Ge T D, Chen C Y, O'Donnell A G, Wu J S. Significant role for microbial autotrophy in the sequestration of soil carbon. Applied and Environmental Microbiology, 2012, 78(7): 2328–2336. DOI:10.1128/AEM.06881-11 |
| [30] | Von Lützow M, Kögel-Knabner I. Temperature sensitivity of soil organic matter decomposition-What do we know?. Biology and Fertility of Soils, 2009, 46(1): 1–15. DOI:10.1007/s00374-009-0413-8 |
| [31] | Conant R T, Ryan M G, Ågren G I, Birge H E, Davidson E A, Eliasson P E, Evans S E, Frey S D, Giardina C P, Hopkins F M, Hyvönen R, Kirschbaum M U F, Lavallee J M, Leifeld J, Parton W J, Steinweg J M, Wallenstein M D, Wetterstedt J Å M, Bradford M A. Temperature and soil organic matter decomposition rates-synthesis of current knowledge and a way forward. Global Change Biology, 2011, 17(11): 3392–3404. DOI:10.1111/j.1365-2486.2011.02496.x |
| [32] | 王兵, 姜艳, 郭浩, 赵广东, 白秀兰. 土壤呼吸及其三个生物学过程研究. 土壤通报, 2011, 42(2): 483–490. |
| [33] | 陈敏鹏, 夏旭, 李银坤, 梅旭荣. 土壤呼吸组分分离技术研究进展. 生态学报, 2013, 33(22): 7067–7077. |
| [34] | Remus R, Hüve K, Pörschmann J, Augustin J. Determining the timepoint when 14C tracer accurately reflect photosynthate use in the plant-soil system. Plant and Soil, 2016, 408(1-2): 457–474. DOI:10.1007/s11104-016-3002-2 |
| [35] | Remus R, Augustin J. Dynamic linking of 14C partitioning with shoot growth allows a precise determination of plant-derived C input to soil. Plant and Soil, 2016, 408(1-2): 493–513. DOI:10.1007/s11104-016-3006-y |
| [36] | Carbone M S, Richardson A D, Chen M, Davidson E A, Hughes H, Savage K E, Hollinger D Y. Constrained partitioning of autotrophic and heterotrophic respiration reduces model uncertainties of forest ecosystem carbon fluxes but not stocks. Journal of Geophysical Research:Biogeosciences, 2016, 121(9): 2476–2492. DOI:10.1002/2016JG003386 |
| [37] | Kuzyakov Y. Separating microbial respiration of exudates from root respiration in non-sterile soils:A comparison of four methods. Soil Biology and Biochemistry, 2002, 34(11): 1621–1631. DOI:10.1016/S0038-0717(02)00146-3 |
| [38] | Kelting D L, Burger J A, Edwards G S. Estimating root respiration, microbial respiration in the rhizosphere, and root-free soil respiration in forest soils. Soil Biology and Biochemistry, 1998, 30(7): 961–968. DOI:10.1016/S0038-0717(97)00186-7 |
| [39] | Chen C R, Condron L M, Xu Z H, Davis M R, Sherlock R R. Root, rhizosphere and root-free respiration in soils under grassland and forest plants. European Journal of Soil Science, 2006, 57(1): 58–66. DOI:10.1111/ejs.2006.57.issue-1 |
| [40] | Craine J M, Wedin D A, Chapin F S. Predominance of ecophysiological controls on soil CO2 flux in a Minnesota grassland. Plant and Soil, 1999, 207(1): 77–86. |
| [41] | Larionova A A, Sapronov D V, de Gerenyu L, Kuznetsova L G, Kudeyarov V N. Contribution of plant root respiration to the CO2 emission from soil. Eurasian Soil Science, 2006, 39(10): 1127–1135. DOI:10.1134/S1064229306100103 |
| [42] | Barba J, Yuste J C, Poyatos R, Janssens I A, Lloret F. Strong resilience of soil respiration components to drought-induced die-off resulting in forest secondary succession. Oecologia, 2016, 182(1): 27–41. DOI:10.1007/s00442-016-3567-8 |
| [43] | Cheng W X, Zhang Q L, Coleman D C, Carroll C R, Hoffman C A. Is available carbon limiting microbial respiration in the rhizosphere?. Soil Biology and Biochemistry, 1996, 28(10-11): 1283–1288. DOI:10.1016/S0038-0717(96)00138-1 |
| [44] | Kuzyakov Y, Cheng W. Photosynthesis controls of rhizosphere respiration and organic matter decomposition. Soil Biology and Biochemistry, 2001, 33(14): 1915–1925. DOI:10.1016/S0038-0717(01)00117-1 |
| [45] | Kuzyakov Y, Domanski G. Model for rhizodeposition and CO2 efflux from planted soil and its validation by 14C pulse labelling of ryegrass. Plant and Soil, 2002, 239(1): 87–102. DOI:10.1023/A:1014939120651 |
| [46] | Sapronov D V, Kuzyakov Y V. Separation of root and microbial respiration:comparison of three methods. Eurasian Soil Science, 2007, 40(7): 775–784. DOI:10.1134/S1064229307070101 |
| [47] | O'Leary M H. Carbon isotopes in photosynthesis. Bioscience, 1988, 38(5): 328–336. DOI:10.2307/1310735 |
| [48] | Kuzyakov Y. Theoretical background for partitioning of root and rhizomicrobial respiration by δ13C of microbial biomass. European Journal of Soil Biology, 2005, 41(1-2): 1–9. DOI:10.1016/j.ejsobi.2005.07.002 |
| [49] | Werth M, Subbotina I, Kuzyakov Y. Three-source partitioning of CO2 efflux from soil planted with maize by 13C natural abundance fails due to inactive microbial biomass. Soil Biology and Biochemistry, 2006, 38(9): 2772–2781. DOI:10.1016/j.soilbio.2006.04.032 |
| [50] | Blagodatskaya E, Khomyakov N, Myachina O, Bogomolova I, Blagodatsky S, Kuzyakov Y. Microbial interactions affect sources of priming induced by cellulose. Soil Biology and Biochemistry, 2014, 74: 39–49. DOI:10.1016/j.soilbio.2014.02.017 |
| [51] | Kerré B, Hernandez-Soriano M C, Smolders E. Partitioning of carbon sources among functional pools to investigate short-term priming effects of biochar in soil:A 13C study. Science of Total Environment, 2016, 547: 30–38. DOI:10.1016/j.scitotenv.2015.12.107 |
| [52] | 宋文琛, 同小娟, 张劲松, 孟平, 李俊. 用自然13C丰度法区分人工林根源呼吸的原理与应用. 中国水土保持科学, 2015, 13(4): 37–43. |
| [53] | Song W C, Tong X J, Zhang J S, Meng P. Three-source partitioning of soil respiration by 13C natural abundance and its variation with soil depth in a plantation. Journal of Forestry Research, 2016, 27(3): 533–540. DOI:10.1007/s11676-015-0206-x |
| [54] | Tomè E, Ventura M, Folegot S, Zanotelli D, Montagnani L, Mimmo T, Tonon G, Tagliavini M, Scandellari F. Mycorrhizal contribution to soil respiration in an apple orchard. Applied Soil Ecology, 2016, 101: 165–173. DOI:10.1016/j.apsoil.2016.01.016 |
| [55] | Werth M, Kuzyakov Y. 13C fractionation at the root-microorganisms-soil interface:A review and outlook for partitioning studies. Soil Biology and Biochemistry, 2010, 42(9): 1372–1384. DOI:10.1016/j.soilbio.2010.04.009 |
| [56] | Tian J, Pausch J, Yu G R, Blagodatskaya E, Kuzyakov Y. Aggregate size and glucose level affect priming sources:A three-source-partitioning study. Soil Biology and Biochemistry, 2016, 97: 199–210. DOI:10.1016/j.soilbio.2016.03.013 |
| [57] | Dijkstra F A, Carrillo Y, Pendall E, Morgan J A. Rhizosphere priming:a nutrient perspective. Frontiers in Microbiology, 2013, 4(1): 65–72. |
| [58] | Sullivan B W, Hart S C. Evaluation of mechanisms controlling the priming of soil carbon along a substrate age gradient. Soil Biology and Biochemistry, 2013, 58: 293–301. DOI:10.1016/j.soilbio.2012.12.007 |
| [59] | Burns R G, DeForest J L, Marxsen J, Sinsabaugh R L, Stromberger M E, Wallenstein M D, Weintraub M N, Zoppini A. Soil enzymes in a changing environment:current knowledge and future directions. Soil Biology and Biochemistry, 2013, 58: 216–234. DOI:10.1016/j.soilbio.2012.11.009 |
| [60] | Spohn M, Pötsch E M, Eichorst S A, Woebken D, Wanek W, Richter A. Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland. Soil Biology and Biochemistry, 2016, 97: 168–175. DOI:10.1016/j.soilbio.2016.03.008 |
| [61] | Zang H D, Wang J Y, Kuzyakov Y. N fertilization decreases soil organic matter decomposition in the rhizosphere. Applied Soil Ecology, 2016, 108: 47–53. DOI:10.1016/j.apsoil.2016.07.021 |
| [62] | 耿元波, 史晶晶. 碳同位素在草地土壤呼吸区分中的应用. 中国农业科学, 2012, 45(17): 3541–3550. |
| [63] | 魏书精, 罗碧珍, 魏书威, 孙龙, 文正敏, 胡海清. 森林生态系统土壤呼吸测定方法研究进展. 生态环境学报, 2014, 23(3): 504–514. |
| [64] | Qiao N, Schaefer D, Blagodatskaya E, Zou X M, Xu X L, Kuzyakov Y. Labile carbon retention compensates for CO2 released by priming in forest soils. Global Change Biology, 2014, 20(6): 1943–1954. DOI:10.1111/gcb.12458 |
| [65] | Pan Y D, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson P B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988–993. DOI:10.1126/science.1201609 |