 2017, Vol. 37
2017, Vol. 37文章信息
- 侯嫚嫚, 李晓宇, 王均伟, 刘帅, 赵秀海.
- HOU Manman, LI Xiaoyu, WANG Junwei, LIU Shuai, ZHAO Xiuhai.
- 长白山针阔混交林不同演替阶段群落系统发育和功能性状结构
- Phylogenetic development and functional structures during successional stages of conifer and broad-leaved mixed forest communities in Changbai Mountains, China
- 生态学报. 2017, 37(22): 7503-7513
- Acta Ecologica Sinica. 2017, 37(22): 7503-7513
- http://dx.doi.org/10.5846/stxb201609141860
-
文章历史
- 收稿日期: 2016-09-14
- 修订日期: 2017-01-16





2. 北京林业大学理学院, 北京 100083
2. College of Science, Beijing Forestry University, Beijing 100083, China
群落构建机制一直是生态学研究的热点问题[1], 生态位理论认为生境过滤、竞争排斥等确定性因素主导群落构建[2], 中性理论则支持扩散和随机作用是群落构建的决定因素[3]。根据系统发育生态位保守理论, 群落系统发育聚集结构可能是由于环境过滤为主的生态过程, 而系统发育发散可能是由竞争排斥作用主导[4-7]。随机的系统发育结构可能是随机的群落构建过程产生或是生境过滤和竞争排斥综合作用的结果[3, 8]。系统发育和功能性状结构的综合研究可以更好地揭示群落构建成因[9-10]。
森林群落演替进程中, 生态过程相对重要性发生变化, 早期在群落中占据主导作用的生态过程可能不再重要[11], 因此, 系统研究不同演替阶段群落构建机制十分必要。近年来, 生态学家在温带和热带森林开展了系统发育和功能性状α多样性的研究, 一些研究结果并不支持随机过程群落构建的作用[9-10, 12]。而Schleicher等发现不同演替阶段的功能性状的α多样性与随机的预测没有差别[13]。对亚热带森林演替阶段系统发育α多样性的研究发现演替后期物种的亲缘关系比演替早期更远[8, 14-15]。此外, 系统发育和功能性状β多样性也为探讨群落构建提供新视角。Swenson针对β多样性研究发现, 演替阶段之间的系统发育周转是随机的, 而功能性状的周转比预测的高或低[16]。Oliver等研究发现不同演替阶段之间的功能性状周转比预测的高, 表明不同演替阶段对群落构建起主导作用的因素不同[17]。
早期对长白山针阔混交林不同演替阶段的研究主要基于物种的群落组成和性状变化[11, 18-22]。然而, 单纯的基于物种类群的途径并没有考虑到物种之间的生态位差异, 而基于性状的研究并未顾及到性状与进化之间的关联性[23-24]。这些限制可能会使群落构建过程的结论出现偏差[25]。群落系统发育和功能性状结构的综合分析可以更好的理解演替动态的群落构建[8, 14-15, 26-27]。
本文采用空间代替时间的“年代序列样地法”构建了演替序列, 以长白山区典型次生杨桦林(secondary Populus davidiana-Betula platyhylla forest)、次生针阔混交林(secondary conifer and broad-leaved mixed forest)、原始椴树红松林(Primary Tilia amurensis-Pinus koraiensis mixed forest)建立的永久森林检测样地为研究对象, 分析了不同演替阶段不同径级水平下的群落系统发育和功能性状结构, 探究以下内容:(1)各功能性状是否具有系统发育信号; (2)不同演替阶段不同径级的系统发育和功能性状结构;(3)不同演替阶段主导群落构建的生态学过程。
1 材料与方法 1.1 试验样地概况本研究样地位于长白山自然保护区, 该地区是受季风影响的温带大陆性山地气候;年平均气温为3.6℃, 最冷月1月的平均气温为-15.4℃, 最热月7月的平均气温为19.6℃, 极端高温为32.3℃, 极端低温为-37.6℃;年降雨量为707 mm;该地区土壤类型为山地暗棕色森林土, 土层厚度为20—100 cm[19]。
1.2 样地设置于2005、2006和2007年, 分别在长白山地区典型的次生杨桦林、次生针阔混交林和原始椴树红松林内建立5.2 hm2(260 m × 200 m)固定监测样地, 它们的林龄约为80 a、120 a和300 a, 构成了一个由次生林逐渐向顶级植被类型过渡的典型演替系列[19]。将每个样地划分成130个20 m × 20 m的样方, 将样方内所有胸径(DBH)≥1 cm的木本植物挂牌标记, 并调查其种类、胸径、树高和坐标等[19]。研究在20 m × 20 m的尺度进行, 以样地内所有胸径≥1 cm的木本植物为研究对象, 次生杨桦林利用2009年的复测数据, 次生针阔混交林利用2011年的复测数据, 原始椴树红松林利用2012年的复测数据。样地概况如表 1所示[11]。
| 林型 Forest type |
经纬度 Longitude and latitude |
平均海拔 Average elevation/m |
总胸高断面积 Total basal area/ (m2/hm2) |
优势树种 Dominant species |
原始植被 Primary vegetation |
| 次生杨桦林 Secondary Populus davidiana-Betula platyphylla forest |
42°19.162′ N 128°07.82′ E |
899 | 24.74 | 白桦(Betula platyphylla) 紫椴(Tilia amurensis) 水曲柳(Fraxinus mandshurica) 山杨(Populus daviana) |
阔叶红松林 Broad-leaved Pinus koraiensis forest |
| 次生针阔混交林 Secondary conifer and broad-leaved mixed forest |
42°20.91′ N 128°07.99′ E |
748 | 32.07 | 紫椴(Tilia amurensis) 白桦(Betula platyphylla) 大青杨(Populus ussuriensis) 臭松(Abies nephrolepis) |
|
| 原始椴树红松林 Primary Tilia amurensis-Pinus koraiensismixed forest |
42°13.68′ N 128°04.57′ E |
1042 | 56.64 | 红松(Pinus koraiensis) 蒙古栎(Quercus mongolica) 紫椴(Tilia amurensis) 水曲柳(Fraxinus mandshurica) |
本研究参照Cornelissen的功能性状采集手册[28], 测量了长白山不同演替阶段各物种的6个功能性状:叶面积(leaf area, cm2)、比叶面积(specific leaf area, cm2/g)、叶片厚度(leaf thickness, mm)、最大树高(maximum height, m)、叶片氮含量(leaf nitrogen concentration, mg/g)以及叶片磷含量(leaf phosphorus concentration, mg/g)。在2015年7月初进行样品采集, 采集对象为样地中胸径≥1 cm的木本植物, 每一个物种选择6—12株长势良好且尽量包含每一个径级的成年个体, 使用高枝剪从个体的冠层且向阳面剪取至少2根比较完整(包括当年叶和往年叶)的末端小枝, 装入自封袋中带回室内进行处理。树种径级划分如表 2。
| 径级划分 Diameter class |
胸径/cm Diameter at breast height |
径级划分 Diameter class |
胸径/cm Diameter at breast height |
| 小径级Small diameter | 1≤DBH≤5 | 大径级Large diameter | DBH>15 |
| 中等径级Medium diameter | 5 < DBH≤15 |
处理方法:(1)叶面积代表了植物对光的捕获能力[28], 在采集叶片的当天, 每一个样本选取至少20片完整的叶片, 剪去叶柄, 利用佳能扫描仪(Lide120, Canon, Japan)在300 dpi灰度模式下扫描以计算叶面积。(2)比叶面积是单位质量叶片的面积, 反映了植物叶片单位质量干物质所捕获光能的能力[28]。(3)叶片厚度反映植物高投入和抗干扰的能力[28]。选用精度为0.02 mm的游标卡尺, 在叶片上沿着主脉的方向均匀选3个点, 测量叶片主脉一侧约0.25 cm处的厚度, 3个厚度的平均值即为叶片的厚度, 每个物种20片, 其平均值即该物种叶片的平均厚度。(4)氮元素和磷元素影响植物的新陈代谢, 对植物光合速率有重要作用[29]。叶片氮、磷含量使用流动分析仪测定。(5)最大树高是和光竞争相关的重要功能性状指标[28]。在植物生长季结束后的11月初, 样地内的每个物种选取最大胸径同等径级的植株25株, 利用测高仪测定每株树的树高, 选取最大值作为该物种的最大树高。
1.4 系统发育树构建本研究通过物种的科属信息, 利用Phylomatic和Phylocom软件构建系统发育树。将每个样地中的全部物种的科属信息输入到网页版Phylomatic软件中, 该软件以被子植物分类系统Ⅲ(APGⅢ)为基础, 自动构建物种的系统发育拓扑结构[6]。然后在Phylocom软件中按照BLADJ算法[30], 根据分子及化石定年数据, 拟合系统发育拓扑结构的分支结构, 得到系统发育树[31]。不同演替阶段群落物种径级分布如图 1。

|
| 图 1 不同演替阶段各径级的物种分布 Fig. 1 Species distribution of different diameter classes in different successional stages (a):次生杨桦林Secondary Populus davidiana-Betula platyphylla forest; (b):次生针阔混交林Secondary conifer and broad-leaved mixed forest; (c):原始椴树红松林Primary Tilia amurensis-Pinus koraiensis mixed forest; SS:小径级, Small diameter; MS:中等径级, Medium diameter; LS:大径级, Large diameter;黑色圆圈代表物种 |
首先将功能性状数据标准化后进行主成分分析(principal component analysis, PCA), 选取能够解释变异95%的主成分, 然后计算欧式距离, 将功能性状矩阵转化为距离矩阵, 最后根据功能性状距离进行层次聚类生成功能性状聚类树[32]。
1.6 功能性状的系统发育信号检验本文利用Blomberg等提出的Brownian motion进化模型K值检验系统发育信号[32]。K=1对应布朗运动(Brownian motion)进化模型;K>1, 表明功能性状具有比按布朗运动模型进化更强的系统发育信号;K < 1, 表明功能性状具有比按布朗运动模型进化更弱的系统发育信号。为了衡量功能性状系统发育信号的显著性, 将系统发育树分支末端的物种随机置换999次生成零模型, 然后比较实际值与零模型的K值, 如果实际值大于95%的零模型的系统发育信号(P < 0.05), 表示功能性状具有显著的系统发育信号, 否则功能性状没有显著的系统发育信号[33]。
1.7 群落系统发育和功能性状结构本研究采用净亲缘关系指数(net relatedness index, NRI)来计算系统发育结构, 并用平均成对性状距离指数(mean pairwise trait distance, PW)来计算功能性状结构。首先计算样方中所有物种对的平均系统发育距离和平均成对性状距离, 保持物种数量及个体数目不变, 将系统发育树或功能性状聚类树末端的物种随机置换999次, 从而获得样方中物种在随机零模型下各物种对的平均系统发育距离或平均成对性状距离, 利用随机分布结果将观察值标准化, 获得NRI和S.E.S PW的值[33]。其计算公式分别为:

|
MPD表示物种间的平均系统发育距离, MPDobs代表实际观察到的MPD值, MPDrand代表随机产生的999个零群落的MPD值, mean(MPDrand)为零群落MPD的平均值, sd(MPDrand)为零群落的MPD的标准差。PW表示平均成对性状距离[33]。PWobs表示实际观察的PW值, PWrand代表随机产生的999个零群落的PW值, PWrand为零群落PW的平均值, sd(PWrand)为零群落的标准差[33]。NRI>0表示系统发育结构聚集;NRI < 0表示系统发育结构发散;NRI=0表示系统发育结构随机。S.E.S PW>0表示功能性状结构聚集;S.E.S PW < 0表示功能性状结构发散;S.E.S PW=0表示功能性状结构随机[33]。
1.8 群落系统发育和功能性状的β多样性本研究采用平均成对距离(mean pairwise distance, Dpw)来表示3个不同演替阶段群落间系统发育和功能性状的周转。计算方式如下:
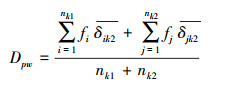
|
其中, ik2表示群落k1中物种i与群落k2中所有物种之间的平均成对距离;nk1表示群落k1的物种数目;fi表示在群落k1物种i的相对多度[34]。假定每个群落的物种数量不变, 群落间共有的物种数量也不变, 通过随机置换999次系统发育树和功能性状树末端的物种来构建零模型, 比较实际观测到的群落系统发育或功能性状的Dpw与零模型产生的Dpw, 以确认群落间系统发育和功能性状的周转是否随机。
计算每个样地130个样方中心两两之间的欧式距离作为空间距离。将土壤有机质、全氮、全磷、全钾、pH、土壤水分、林分开阔度作为环境因子进行标准化处理, 将标准化后环境矩阵的欧式距离作为环境距离。计算130个样方两两之间的平均成对距离, 采用空间距离、环境距离以及两者结合的3种方式对群落系统发育和功能性状的β多样性值进行方差分解, 分析空间距离和环境距离对群落系统发育和功能性状结构的解释程度。这里使用MRM算法(multiple regression on distance matrices, MRM)进行方差分解, 该算法是对偏Mantel检验(partial Mantel test)的简单扩展, 可以有效地对多个矩阵进行方差分解[33]。具体计算使用R-3.2.4 (R Development Core Team, 2015)完成。
2 结果 2.1 功能性状系统发育信号的K值检验长白山阔叶红松林不同演替阶段样地7类功能性状的系统发育信号强度如表 3, 结果表明:在3个样地中, 叶面积、比叶面积、叶片厚度、叶片氮含量、叶片磷含量、氮/磷和最大树高均能检测到显著的系统发育信号(P < 0.05), 表明功能性状与物种的进化历史有密切的关系。其中, 比叶面积的K值最大, 接近于1;次生针阔混交林的性状K值相对较大, 且叶片氮含量和叶片磷含量的K值相对较大, 分别为0.62和0.54;其余性状的K值均小于1, 低于布朗模型的预期值, 说明分析的功能性状与随机模型比较具有显著的系统发育信号, 即可以认为亲缘关系相近的物种具有相似的功能性状。
| 功能性状 Functional traits |
次生杨桦林 Secondary Populus davidiana-Betula platyphylla forest |
次生针阔混交林 Secondary conifer and broad-leaved mixed forest |
原始椴树红松林 Primary Tilia amurensis-Pinus koraiensis mixed forest |
||||||||
| 性状 平均值 |
K值 | P值 | 性状 平均值 |
K值 | P值 | 性状 平均值 |
K值 | P值 | |||
| 叶面积Leaf area /cm2 | 28.3 | 0.22 | 0.018 | 26.1 | 0.35 | 0.023 | 23.7 | 0.22 | 0.019 | ||
| 比叶面积/(cm2/g) Specific leaf area |
0.0 | 0.91 | 0.023 | 0.0 | 0.83 | 0.021 | 0.0 | 0.91 | 0.021 | ||
| 叶片厚度/mm Leaf thickness |
0.3 | 0.41 | 0.013 | 0.2 | 0.89 | 0.002 | 0.2 | 0.41 | 0.007 | ||
| 叶片氮含量/(mg/g) Leaf nitrogen concentration |
18.9 | 0.12 | 0.004 | 17.7 | 0.62 | 0.034 | 19.1 | 0.12 | 0.013 | ||
| 叶片磷含量/(mg/g) Leaf phosphorus concentration |
24.6 | 0.18 | 0.002 | 24.6 | 0.54 | 0.021 | 24.1 | 0.18 | 0.024 | ||
| 氮/磷N/P | 0.8 | 0.18 | 0.013 | 0.8 | 0.38 | 0.017 | 0.8 | 0.18 | 0.037 | ||
| 最大树高/m Maximum height |
18.4 | 0.16 | 0.022 | 17.8 | 0.37 | 0.019 | 21.8 | 0.14 | 0.004 | ||
不同演替阶段群落系统发育和功能性状结构如图 2, 次生杨桦林和次生针阔混交林NRI>0, 而原始椴树红松林NRI < 0, 表明随着演替的进行, 谱系结构由聚集到发散变化。次生杨桦林和次生针阔混交林的S.E.S PW均大于0, 原始椴树红松林S.E.S PW < 0, 表明随着演替的进行, 群落功能性状结构由聚集到发散变化。

|
| 图 2 长白山针阔混交林不同演替阶段群落的系统发育和功能性状结构 Fig. 2 Phylogenetic and functional structure of different successional stages in Changbai Mountains (a):不同演替阶段群落系统发育结构(平均值±标准误;“*”表示P < 0.05, “**”表示P < 0.01);(b):不同演替阶段群落功能性状结构(平均值±标准误;“*”表示P < 0.05, “**”表示P < 0.01);S1:次生杨桦林, Secondary Populus davidiana-Betula platyphylla forest; S2:次生针阔混交林, Secondary conifer and broad-leaved mixed forest;S3:原始椴树红松林, Primary Tilia amurensis-Pinus koraiensis mixed forest |
不同径级群落系统发育和功能性状结构如图 3, 在各演替阶段, 不同径级的系统发育结构均非随机(P < 0.05, 图 3a)。小径级物种NRI均为正值, 表现出显著的聚集结构, 随着径级的增加, NRI逐渐减小, 直至变为负值, 谱系聚集程度减弱到发散的状态。次生杨桦林和次生针阔混交林, 小径级系统发育结构聚集, 中等径级和大径级系统发育结构发散。原始椴树红松林小径级系统发育结构聚集, 中等径级的群落系统发育结构发散, 而大径级群落系统发育结构聚集。随着演替的推进, 群落系统发育结构由聚集到发散变化, 随着径级的增加, 系统发育结构也是由聚集到发散变化。

|
| 图 3 长白山针阔混交林不同演替阶段不同径级树种的系统发育和功能性状结构 Fig. 3 Phylogenetic(a)and functional(b)structure of different diameter class during successional stages in Changbai Mountains (a):不同径级群落系统发育结构(平均值±标准误;“*”表示P < 0.05, “**”表示P < 0.01);(b):不同径级群落功能性状结构(平均值±标准误;“*”表示P < 0.05, “**”表示P < 0.01);S1, 次生杨桦林, Secondary Populus davidiana-Betula platyphylla forest; S2:次生针阔混交林, Secondary conifer and broad-leaved mixed forest;S3:原始椴树红松林, Primary Tilia amurensis-Pinus koraiensis mixed forest |
不同径级的功能性状结构均非随机(P < 0.05, 图 3b)。小径级物种的S.E.S PW均大于0, 表现为功能性状结构聚集;除原始椴树红松林外, 次生杨桦林和次生针阔混交林中等径级和大径级物种的S.E.S PW均小于0, 表现为功能性状结构发散。随着演替的推进, 群落的功能性状结构由聚集到发散, 且从小树到大树, 物种的功能性状结构也呈现由聚集到发散的变化。
2.4 不同演替阶段群落系统发育和功能性状β多样性不同演替阶段的系统发育和功能性状的标准化平均成对距离如表 4, S.E.S.DPW均偏离零值, 说明系统发育和功能性状的周转在各个演替阶段均为非随机。系统发育和功能性状在各演替阶段之间的变化如图 4。次生杨桦林和原始椴树红松林的系统发育S.E.S.DPW平均值均为正值, 次生针阔混交林的系统发育S.E.S.DPW平均值为负值, 说明演替早期和演替后期的系统发育在各小样方之间的变化比预期的快, 而演替中期系统发育在各小样方之间的变化比预期的慢。而各演替阶段的功能性状S.E.S.DPW平均值均为正值, 说明各演替阶段的功能性状在各小样方之间的周转比预期的快。

|
| 图 4 长白山样地各演替阶段之间系统发育和功能性状的周转 Fig. 4 Temporal phylogenetic and functional turnover between successional stages (a):系统发育结构周转(平均值±标准误);(b):功能性状周转(平均值±标准误);S1:次生杨桦林, Secondary Populus davidiana-Betula platyphylla forest; S2:次生针阔混交林, Secondary conifer and broad-leaved mixed forest;S3:原始椴树红松林, Primary Tilia amurensis-Pinus koraiensis mixed forest |
| 样地Sample plot | 系统发育标准化平均成对距离 S.E.S. Dpw of phylogene |
功能性状标准化平均成对距离 S.E.S. Dpw of functioal traits |
| 次生杨桦林 Secondary Populus davidiana-Betula platyphylla forest |
0.501±0.91 | 0.835±2.14 |
| 次生针阔混交林 Secondary conifer and broad-leaved mixed forest |
-1.55±1.82 | 0.764±2.45 |
| 原始椴树红松林 Primary Tilia amurensis-Pinus koraiensis mixed forest |
0.235±2.34 | 0.453±3.42 |
长白山样地系统发育和功能性状β多样值与空间距离环境距离的方差分解检验结果如表 5, 次生杨桦林:空间距离对系统发育和功能性状的β多样性的解释力度小于环境距离的解释力度;次生针阔混交林:空间距离对系统发育β多样性的解释力度大于环境距离, 而对功能性状的β多样性的解释力度小于环境距离;原始椴树红松林:环境距离对系统发育和功能性状β多样性的解释力度小于空间距离。
| 样地 Sample plot |
解释变量 Explanatory variable |
群落系统发育β多样性 Phylogenetic beta diversity |
群落功能性状β多样性 Functional beta diversity |
| 次生杨桦林 | M.S.E | 0.0032 | 0.0079 |
| Secondary Populus davidiana- | M.S | 0.0021 | 0.0076 |
| Betula platyphylla forest | M.E | 0.0020 | 0.0015 |
| M.P.S | 0.0010 | 0.0013 | |
| M.P.E | 0.0014 | 0.0043 | |
| 次生针阔混交林 | M.S.E | 0.0019 | 0.0386 |
| Secondary conifer and broad- | M.S | 0.0008 | 0.0032 |
| leaved mixed forest | M.E | 0.0006 | 0.0362 |
| M.P.S | 0.0013 | 0.0024 | |
| M.P.E | 0.0010 | 0.0355 | |
| 原始椴树红松林 | M.S.E | 0.0235 | 0.0050 |
| Primary Tilia amurensis-Pinus | M.S | 0.0074 | 0.0047 |
| koraiensis mixed forest | M.E | 0.0175 | 0.0006 |
| M.P.S | 0.0260 | 0.0104 | |
| M.P.E | 0.0161 | 0.0044 | |
| M.S.E:环境距离和空间距离解释的部分, Multiple regression on distance matrices of environment and space; M.S:空间距离解释的部分, Multiple regression on distance matrices of space; M.E:环境距离解释的部分, Multiple regression on distance matrices of environment; M.P.S:去除环境距离影响后空间距离解释的部分, Multiple regression on distance matrices of pure space; M.P.E:去除空间距离影响后环境距离解释的部分, Multiple regression on distance matrices of pure environment | |||
随着演替的进行, 空间距离对系统发育和功能性状结构转换的解释力度减弱, 而环境距离对系统发育和功能性状结构转换的解释力度增加, 演替早期空间距离的解释力度小于环境距离, 说明生境过滤在群落构建中的重要性, 而在演替后期空间距离的解释力度大于环境距离, 验证了扩散限制在群落构建中的重要性。
3 结论与讨论 3.1 功能性状的系统发育信号研究表明3个研究样地的7个功能性状均表现出显著的系统发育信号(表 3), 证明了群落植物种类功能性状的谱系保守性, 即亲缘关系近的物种具有相似的功能性状, 在此前提下, 系统发育结构的聚集验证了环境过滤作用在群落构建过程中的重要性[9, 35], 而发散则证明了竞争排斥在群落构建过程的重要性[36]。
3.2 不同演替阶段群落系统发育和功能性状结构植物的径级大小体现了物种生长的时间尺度[37]。本文研究已经证明共存物种间的功能性状具有显著的系统发育信号, 即是功能性状谱系保守。在这一结论基础上, 不同径级水平的系统发育结构支持随着个体发育种间作用增强, 大树之间由于对有限资源的争夺, 导致发散的系统发育结构, 说明了竞争排斥作用在径级尺度上的群落构建过程中起作用。Swenson等研究5个径级尺度下植物的系统发育结构, 发现小径级的系统发育结构聚集或随机, 而大径级的系统发育结构则趋向于发散格局[10]。本文研究与前人研究结果相似:小径级物种系统发育均表现出显著的聚集结构, 随着径级的增加, 聚集程度减弱到发散的状态, 这可能是由于种子受到扩散限制的影响, 小树聚集生长, 表现为系统发育结构聚集;随着个体的生长, 树木彼此之间的竞争加强, 存活的物种之间地理距离变远, 表现为系统发育结构发散[38-39]。
长白山针阔混交林不同发育阶段群落系统发育和功能性状结构存在较大差异, 主导群落构建的生态学过程也不同。演替早期的次生杨桦林, 先锋树种占主导作用, 先锋树种种子小、传播广、生长快, 对环境的适应能力强, 在干扰过后可以直接建立一个植物群落, 使得早期群落是由亲缘关系相近的物种所组成, 从而增强了次生杨桦林聚集的谱系结构。早期演替的先锋树种的寿命很短, 导致森林更新过程中早期演替物种的减少, 逐渐被演替后期的物种所代替。大多数演替后期的物种都有较好的耐阴性, 并且种子的传播受扩散限制的作用减弱, 物种数目增加。因此, 物种对光、营养、水分等环境资源的竞争会增加, 这一阶段物种间的竞争排斥作用占主导。竞争排斥作用会使拥有相似生态位的物种(亲缘关系近)分散, 最终导致群落谱系的发散。本研究显示, 随着演替的推进, 群落系统发育和功能性状结构由聚集到发散发生变化, 结果与前人的研究结果相一致[15, 26], 表明在演替早期环境过滤作用在群落构建过程中占主导, 而在演替的后期阶段竞争排斥作用占主导[15]。此外, MRM方差分解结果显示, 次生林中环境距离对群落的系统发育和功能性状转换的解释力度较高, 而原始椴树红松林中空间距离对群落系统发育和功能性状转换的解释力度较高, 验证了以上结论。
3.3 群落构建中的生态学过程Mo等研究表明系统发育结构由演替早期的年轻次生林聚集到老的次生林中发散, 最后在演替后期老龄林中系统发育结构表现为随机状态[40], 研究表明随机状态可能是由生境过滤和竞争排斥共同作用的结果, 结果与中性理论一致。在本文的研究中, 群落系统发育结构和功能性状结构在不同演替阶段均表现出非随机的结构, 这一结果与中性理论预测不一致。而杨洁等的研究发现, 在小尺度上(5 m × 5 m), 中等径级树种呈现随机的系统发育和功能性状结构[39]。说明中性过程可能在小尺度上发挥作用, 而本文的研究尺度是20 m × 20 m, 中性过程在这一尺度不起作用或作用不明显。因此, 本研究的结果支持生态位过程在长白山针阔混交林不同演替阶段群落构建过程中的作用。综上所述, 对系统发育和功能性状结构的研究表明, 环境过滤和竞争排斥过程对长白山针阔混交林不同演替阶段的的群落构建起到了重要作用, 而中性过程并未表现出明显的作用。
此外, 闫琰等对长白山针阔混交林不同演替阶段多度分布格局的研究证实了中性过程在长白山温带针阔混交林群落结构形成中具有重要作用, 但未否认生态位机制在群落构建中的贡献[19]。原因可能是单纯基于物种类群的途径并未考虑到生态位之间的差异, 从而导致群落构建过程中的结论可能出现偏差, 因此需要综合分析群落系统发育和功能性状结构以便更好的理解演替动态的群落构建。
| [1] | Kunstler G, Lavergne S, Courbaud B, Thuiller W, Vieilledent G, Zimmermann N E, Kattge J, Coomes D A. Competitive interactions between forest trees are driven by species' trait hierarchy, not phylogenetic or functional similarity:implications for forest community assembly. Ecology Letters, 2012, 15(8): 831–840. DOI:10.1111/j.1461-0248.2012.01803.x |
| [2] | Tilman D. Resource Competition and Community Structure. Princeton: Princeton University Press, 1982: 1–296. |
| [3] | Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton: Princeton University Press, 2001: 340–348. |
| [4] | Vamosi S M, Heard S B, Vamosi J C, Webb C O. Emerging patterns in the comparative analysis of phylogenetic community structure. Molecular Ecology, 2009, 18(4): 572–592. DOI:10.1111/mec.2009.18.issue-4 |
| [5] | Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2003, 33: 475–505. |
| [6] | Webb C O, Donoghue M J. Phylomatic:tree assembly for applied phylogenetics. Molecular Ecology Notes, 2005, 5(1): 181–183. DOI:10.1111/men.2005.5.issue-1 |
| [7] | Kembel S W, Hubbell S P. The phylogenetic structure of a neotropical forest tree community. Ecology, 2006, 87(sp7): S86–S99. DOI:10.1890/0012-9658(2006)87[86:TPSOAN]2.0.CO;2 |
| [8] | Letcher S G. Phylogenetic structure of angiosperm communities during tropical forest succession. Proceedings of the Royal Society B:Biological Sciences, 2010, 277(1678): 97–104. DOI:10.1098/rspb.2009.0865 |
| [9] | Kraft N J B, Ackerly D D. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest. Ecology Monographs, 2010, 80(3): 401–422. DOI:10.1890/09-1672.1 |
| [10] | Swenson N G, Enquist B J, Thompson J, Zimmerman J K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology, 2007, 88(7): 1770–1780. DOI:10.1890/06-1499.1 |
| [11] | 张春雨, 赵秀海, 赵亚洲. 长白山温带森林不同演替阶段群落结构特征. 植物生态学报, 2009, 33(6): 1090–1100. |
| [12] | Cavender-Bares J, Kozak K H, Fine P V A, Kembel S W. The merging of community ecology and phylogenetic biology. Ecology Letters, 2009, 12(7): 693–715. DOI:10.1111/ele.2009.12.issue-7 |
| [13] | Schleicher A, Peppler-Lisbach C, Kleyer M. Functional traits during succession:is plant community assembly trait-driven?. Preslia, 2011, 83: 347–370. |
| [14] | Ding Y, Zang R G, Letcher S G, Liu S R, He F L. Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests. Oikos, 2012, 121(8): 1263–1270. DOI:10.1111/more.2012.121.issue-8 |
| [15] | Letcher S G, Chazdon R L, Andrade A C S, Bongers F, van Breugel M, Finegan B, Laurance S G, Mesquita R C G, Martínez-Ramos M, Williamson G B. Phylogenetic community structure during succession:evidence from three Neotropical forest sites. Perspectives in Plant Ecology, Evolution and Systematics, 2012, 14(2): 79–87. DOI:10.1016/j.ppees.2011.09.005 |
| [16] | Swenson N G, Stegen J C, Davies S J, Erickson D L, Forero-Montaña J, Hurlbert A H, Kress W J, Thompson J, Uriarte M, Wright S J, Zimmerman J K. Temporal turnover in the composition of tropical tree communities:functional determinism and phylogenetic stochasticity. Ecology, 2012, 93(3): 490–499. DOI:10.1890/11-1180.1 |
| [17] | Purschke O, Schmid B C, Sykes M T, Poschlod P, Michalski S G, Durka W, Kühn I, Winter M, Prentice H C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession:insights into assembly processes. Journal of Ecology, 2013, 101(4): 857–866. DOI:10.1111/1365-2745.12098 |
| [18] | 胡耀升, 么旭阳, 刘艳红. 长白山不同演替阶段森林植物功能性状及其与地形因子间的关系. 生态学报, 2014, 34(20): 5915–5924. |
| [19] | 闫琰, 张春雨, 赵秀海. 长白山不同演替阶段针阔混交林群落物种多度分布格局. 植物生态学报, 2012, 36(9): 923–934. |
| [20] | 郭利平, 姬兰柱, 王珍, 王智轩. 长白山红松阔叶林不同演替阶段优势种的变化. 应用生态学报, 2011, 22(4): 866–872. |
| [21] | 夏富才, 赵秀海, 潘春芳, 贾玉珍, 汪金松. 长白山阔叶红松林林分空间结构. 应用与环境生物学报, 2010, 16(4): 529–534. |
| [22] | 胡耀升, 么旭阳, 刘艳红. 长白山森林不同演替阶段植物与土壤氮磷的化学计量特征. 应用生态学报, 2014, 25(3): 632–638. |
| [23] | Marquard E, Weigelt A, Temperton V M, Roscher C, Schumacher J, Buchmann N, Fischer M, Weisser W W, Schmid B. Plant species richness and functional composition drive overyielding in a six-year grassland experiment. Ecology, 2009, 90(12): 3290–3302. DOI:10.1890/09-0069.1 |
| [24] | Reich P B, Tilman D, Naeem S, Ellsworth D S, Knops J, Craine J, Wedin D, Trost J. Species and functional group diversity independently influence biomass accumulation and its response to CO2 and N. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(27): 10101–10106. DOI:10.1073/pnas.0306602101 |
| [25] | Cornwell W K, Schwilk D W, Ackerly D D. A trait-based test for habitat filtering:convex hull volume. Ecology, 2006, 87(6): 1465–1471. DOI:10.1890/0012-9658(2006)87[1465:ATTFHF]2.0.CO;2 |
| [26] | Whitfeld T J S, Kress W J, Erickson D L, Weiblen G D. Change in community phylogenetic structure during tropical forest succession:evidence from New Guinea. Ecography, 2012, 35(9): 821–830. DOI:10.1111/ecog.2012.35.issue-9 |
| [27] | Norden N, Letcher S G, Boukili V, Swenson N G, Chazdon R. Demographic drivers of successional changes in phylogenetic structure across life-history stages in plant communities. Ecology, 2012, 93(sp8): S70–S82. DOI:10.1890/10-2179.1 |
| [28] | Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335–380. DOI:10.1071/BT02124 |
| [29] | Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001–11006. DOI:10.1073/pnas.0403588101 |
| [30] | Wikstrom N. Evolution of the angiosperms:calibrating the family tree. Proceedings of the Royal Society B:Biological Sciences, 2001, 268(1482): 2211–2220. DOI:10.1098/rspb.2001.1782 |
| [31] | Webb C O, Ackerly D D, Kembel S W. Phylocom:software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 2008, 24(18): 2098–2100. DOI:10.1093/bioinformatics/btn358 |
| [32] | Blomberg S P, Garland T Jr, Ives A R. Testing for phylogenetic signal in comparative data:behavioral traits are more labile. Evolution, 2003, 57(4): 717–745. DOI:10.1111/evo.2003.57.issue-4 |
| [33] | Swenson N G. Functional and Phylogenetic Ecology in R. New York:Springer, 2014. |
| [34] | Swenson N G. Phylogenetic beta diversity metrics, trait evolution and inferring the functional beta diversity of communities. PLoS One, 2011, 6(6): e21264. DOI:10.1371/journal.pone.0021264 |
| [35] | Swenson N G, Enquist B J, Pither J, Thompson J, Zimmerman J K. The problem and promise of scale dependency in community phylogenetics. Ecology, 2006, 87(10): 2418–2424. DOI:10.1890/0012-9658(2006)87[2418:TPAPOS]2.0.CO;2 |
| [36] | Mayfield M M, Levine J M. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecology Letters, 2010, 13(9): 1085–1093. DOI:10.1111/j.1461-0248.2010.01509.x |
| [37] | 牛红玉, 王峥峰, 练琚愉, 叶万辉, 沈浩. 群落构建研究的新进展:进化和生态相结合的群落谱系结构研究. 生物多样性, 2011, 19(3): 275–283. |
| [38] | 宋凯, 米湘成, 贾琪, 任海保, BebberD, 马克平. 不同程度人为干扰对古田山森林群落谱系结构的影响. 生物多样性, 2011, 19(2): 190–196. |
| [39] | 杨洁, 卢孟孟, 曹敏, 李捷, 林露湘. 中山湿性常绿阔叶林系统发育和功能性状的α及β多样性. 科学通报, 2014, 59(24): 2349–2358. |
| [40] | Mo X X, Shi L L, Zhang Y J, Zhu H, Slik J W F. Change in phylogenetic community structure during succession of traditionally managed tropical rainforest in southwest China. PLoS One, 2013, 8(7): e71464. DOI:10.1371/journal.pone.0071464 |