 2017, Vol. 37
2017, Vol. 37文章信息
- 唐敬超, 史作民, 罗达, 刘世荣.
- TANG Jingchao, SHI Zuomin, LUO Da, LIU Shirong.
- 遮荫处理对灰木莲幼苗叶片光合氮利用效率的影响
- Photosynthetic nitrogen-use efficiency of Manglietia glauca seedling leaves under different shading levels
- 生态学报. 2017, 37(22): 7493-7502
- Acta Ecologica Sinica. 2017, 37(22): 7493-7502
- http://dx.doi.org/10.5846/stxb201609111838
-
文章历史
- 收稿日期: 2016-09-11
- 网络出版日期: 2017-07-12

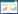




2. 南京林业大学南方现代林业协同创新中心, 南京 210037;
3. 新疆林业科学院经济林研究所, 乌鲁木齐 830063
2. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China;
3. Research Institute of Economic Forestry, Xinjiang Academy of Forestry Science, Urumqi 830063, China
光合氮利用效率(Photosynthetic nitrogen-use efficiency, PNUE)是叶片的固有属性, 是描述植物叶片养分利用、生理特性和生存策略的重要特征[1], 是解释叶片经济学的生物化学关键因素[2-4]。由于PNUE是单位面积叶片光饱和净光合速率(Amax)与单位面积叶片氮含量(Narea)的比值, 因此叶片Amax与Narea的变化就会影响PNUE[5]。而从生物化学的角度来看, 羧化位CO2浓度, 叶片分配到光合作用限制步骤的氮占总叶氮的比例以及光合酶的活性都会影响PNUE[3]。叶片中的氮元素是影响PNUE的关键因素[6]。有研究认为, 遮荫会降低植物叶片光合系统中的Rubisco氮分配比例(PR)和生物力能学组分氮分配比例(PB), 提高捕光组分氮分配比例(PL)[7-9]。但也有研究认为遮荫虽然会提高PL, 但并不会降低PR和PB[10-11]。遮荫对PNUE的影响也较为复杂, 随着生长光强的降低, 植物叶片PNUE不变[12-13], 增加[14]或者降低[15]都有相应的研究结果出现。此外, 细胞壁的合成也需要氮的参与, 这部分氮也可能与光合系统的氮形成权衡关系进而影响PNUE[16-17]。
我国南亚热带地区自20世纪80年代开始进行大规模的植树造林[18], 营造了大量以马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)为主的速生纯林[19]。但长期大面积同龄纯林连栽会导致林分出现多种生态问题[20-23]。为了减少人工纯林的不利影响, 将阔叶树种混交到人工纯林中, 进而调整林分的树种组成及结构便成为一种较好的方法[22]。然而混交需要将这些阔叶树种栽种到纯林中开出的林窗内, 林窗所能提供的光照条件就成为制约阔叶树种生长的重要因素。因此对这些树种幼苗叶片在不同生长光照条件下的光合能力及PNUE进行研究就具有较为重要的参考价值与实践意义。
灰木莲(Manglietia glauca)是木兰科常绿乔木, 是适生于我国南亚热带地区的珍贵树种[24]。灰木莲生长快、干型通直、材质良好、树形优美, 具有移植容易成活且移栽恢复快的优点, 是非常优良的绿化及林分改造树种[25]。近些年来的研究多集中于对灰木莲组织培养[26]、引种栽培[27]、苗木生长[28]、人工林生长规律[25]等的研究, 而对灰木莲光合生理生态, 特别是不同遮荫环境下幼苗叶片PNUE的研究较为少见[29-30]。本研究以灰木莲为对象, 对其幼苗叶片在不同遮荫环境下的PNUE及其影响因素进行研究, 并基于该研究提出以下科学问题:1.灰木莲幼苗叶片的PNUE及影响因素在不同遮荫条件下会怎样变化?2.灰木莲幼苗叶片各含氮组分的氮分配比例是否会随着遮荫程度的变化而协同变化?研究结果可以为这种珍贵树种的栽培育苗, 以及人工纯林的改造提供科学理论依据。
1 研究区概况试验地位于广西凭祥市中国林业科学研究院热带林业实验中心苗圃(106°44′40″—106°44′44″E, 22°7′19″—22°7′22″N), 海拔250—300 m。该地区属于南亚热带季风气候, 干、湿季节分明。年均气温21℃, 平均月最低和最高气温分别为12.1℃和26.3℃;年平均降水量1400 mm, 降水主要发生在4—9月;≥10℃活动积温为6000—7600℃;年均日照时数1419 h。
2 材料与方法 2.1 试验材料灰木莲种子采自同一棵母树, 种子于2014年1月份出苗。3月下旬选取长势良好, 大小相近的60株幼苗移栽到装有河沙的花盆中。花盆高18.5 cm, 口直径与底直径分别为21 cm及15 cm, 盆内的河沙用清水洗净。花盆随机均分为三组, 分别放置在全光下(晴天正午可以达到光饱和点), 60%遮荫棚(达不到光饱和点, 但高于光补偿点)及90%遮荫棚(略高于光补偿点)下。2014年4—6月每天进行浇水以保证水分供应充足, 每周使用相同浓度的营养液保证营养供应充足。
2.2 气体交换参数的测定在2014年7—8月的晴天利用Licor-6400便携式光合作用测定系统(LI-COR, Lincoln Nebraska, USA)对目标幼苗叶片进行光响应及CO2响应曲线的测定。每个处理选取5棵生长良好的幼苗, 每个幼苗选取1个叶片。所选叶片位于植株向阳面顶端, 为新展开成熟叶, 健康无病虫害。利用Licor-6400光合仪红蓝光源(6400-02B, LI-COR, Lincoln Nebraska, USA), 由高到低按照1500, 1200, 1000, 800, 600, 400, 200, 150, 100, 80, 50, 20, 0 μmol m-2 s-1梯度测定其光响应曲线, 测定时叶室CO2浓度设定为380 μmol/mol。测定光响应曲线后的第二天, 设定叶室光照在光饱和点, 设定叶室CO2浓度依次为380, 200, 150, 100, 80, 50, 380, 600, 800, 1000, 1200, 1500, 1800, 2000 μmol/mol对同一个叶片测定胞间CO2响应曲线[31-32]。所有气体交换参数测定时叶室温度控制在30(±1)℃, 湿度控制在60%—70%。在每个光强或CO2浓度下平衡200 s后测定净光合速率(A), 测定重复十次。利用光响应曲线计算暗呼吸速率(Rd)及光饱和净光合速率(Amax), 利用胞间CO2响应曲线计算最大羧化速率(Vcmax), 最大电子传递速率(Jmax)及无暗呼吸时CO2补偿点Γ*(μmol/mol)等数据。
2.3 荧光产量的测定运用Licor-6400光合仪荧光叶室(6400-40, LI-COR, Lincoln Nebraska, USA)进行测定。测定选用的目标叶片与气体交换参数测定叶片相同, 测定时的叶室温湿度也与气体交换参数测定时的一致, 叶室CO2浓度设定为380 μmol/mol, 叶室内光强设定为光饱和点。叶片经饱和光强充分活化后进行测定, 测定时的脉冲强度(intensity)、调制频率(rate)、信号平均频率(filter)及信号放大倍数(gain)分别设置为:5档(<1 μmol m-2 s-1)、20 kHz、1 Hz、10倍[33]。每个叶片重复记数10次以上, 测得的荧光产量(⊿F/Fm′)用于叶肉细胞导度(gm)的求算。
2.4 植物叶片基本属性的测定使用CI202手持式叶面积仪(CID, Washington State, Camas, USA)测定叶片面积。随后将叶片分为三份:第一份放入80℃烘箱烘干至恒重计算叶片干重, 随后采用凯氏定氮法测定叶片总氮含量;第二份采用分光光度法测定叶绿素含量;第三份使用化学分离法测定细胞壁氮含量。
2.5 叶肉细胞导度的计算叶肉细胞导度(gm)利用Harley等提出[34], Loreto等改进[35]的计算方法求算。在该方法中:
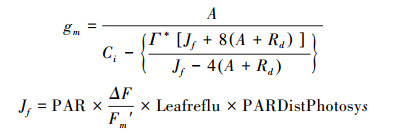
|
式中, Leafreflu为叶片吸收系数, 取值为0.9, PARDistPhotosys为所吸收光能在两个光系统之间的分配比例, 取值为0.5[33];净光合速率A(μmol m-2 s-1), 胞间CO2浓度Ci(μmol/mol), 荧光产量⊿F/Fm′, 光照强度PAR(μmol m-2 s-1)等由相关仪器直接测出;Rd (μmol m-2 s-1)通过光响应曲线计算;Γ*(μmol/mol)通过胞间二氧化碳响应曲线计算。由此可计算出电子传递速率Jf (μmol m-2 s-1), 从而计算出叶肉细胞导度。
2.6 最大羧化速率和最大电子传递速率的计算首先通过胞间二氧化碳响应曲线初始部分(外界CO2浓度在50—200 μmol/mol时)胞间CO2浓度(Ci)与A作拟合直线A =kCi+i, 得到斜率k以及i。其中, Ci为Licor-6400直接测得;k为羧化效率CE;i等于Ci取值为0时的A, 相当于光呼吸速率[14]。然后通过Farquhar[36]提出的方法计算Vcmax:
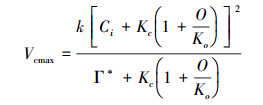
|
Kc为Michaelis-Menten RUBP羧化常量, Ko为Michaelis-Menten RUBP氧化常量, 在30℃时分别取值为469.83 μmol/mol和456.21 mmol/mol[6]。O是胞间氧气浓度, 取值为210 mmol/mol。Γ*(μmol/mol)通过胞间二氧化碳响应曲线计算。
最大电子传递速率通过Loustau提出的方法进行计算[37]:
|
Amax′为光与CO2都饱和时的A, Rd (μmol m-2 s-1)通过光响应曲线计算。
2.7 光合系统内各组分氮分配比例的计算光合系统中各组分的氮分配比例利用Niinemets和Tenhunen提出的计算方法进行求算[6]。在该方法中光合系统中的含氮物质可以按其在光合作用中的功能划分为三部分:Rubisco、生物力能学组分以及捕光组分, 其各部分含氮比例的计算方法如下:
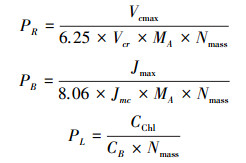
|
其中, PR为氮分配到Rubisco中的比例;PB为氮分配到生物力能学组分中的比例;PL为氮分配到捕光组分中的比例;MA为单位面积叶干重(g/m2);Nmass为单位干重叶片氮含量(g/g);Vcmax单位为(μmol m-2 s-1);Jmax单位为(μmol m-2 s-1);CChl为叶片叶绿素浓度(mmol/g);Vcr是Rubisco比活, 即单位Rubisco的CO2固定活性(μmol CO2 g-1 Rubisco s-1), 与温度相关;Jmc为单位细胞色素f(Cytochrome f, Cyt f)的最大电子传递速率(μmol electrons μmol-1 Cyt f s-1), 与温度相关。Vcr和Jmc的计算方法如下[38-39]:
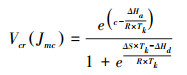
|
其中, R为通用气体常数, 取值为8.314 (J K-1 mol-1);Tk是叶片温度(K), ΔHa、ΔHd、ΔS和c为活化能、去活化能、熵和比例常数, 计算Vcr时的取值分别为74000 (J/mol)、203000 (J/mol)、645 (J K-1 mol-1)和32.9;计算Jmc时的取值分别为24100 (J/mol)、564150 (J/mol)、1810 (J K-1 mol-1)和14.77[6, 40-41]。
CB为捕光组分中叶绿素与氮的比值(mmol Chl/(g N)), 其取值可以利用拟合出的CB的值与MA的值的关系的线性公式来进行计算, 其相关系数较高(r2=0.97), 并且可以在多种植物类型中广泛应用。其公式为:
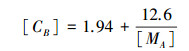
|
其中, [MA]为单位面积叶片干重(MA, 单位g/m2)的值, [CB]为CB的值[6]。这样就可以通过MA的值来计算出CB的值, 以便进一步计算PL[42]。
2.8 细胞壁氮含量的测定各研究对象的细胞壁氮含量采用Onoda等使用的方法进行测定[17]。取1 g鲜样研磨至匀浆, 离心后丢弃上清液, 沉淀依次添加3%(W/V)十二烷基磺酸钠(SDS)、淀粉转葡萄糖苷酶(35 unit/mL, Rhizopus mould, Sigma, St Louis, USA)分别进行加热离心, 剩余沉淀再用蒸馏水及酒精分别进行清洗并离心, 最后将沉淀烘干, 用凯氏定氮法测定其中的氮含量。
2.9 数据处理使用Excel 2007进行数据整理, SPSS 17.0进行数据相关性及方差分析, Orgin 9.0进行作图。
3 结果 3.1 不同遮荫条件下灰木莲幼苗叶片的PNUE不同遮荫条件下的灰木莲幼苗叶片PNUE并没有显著差异(表 1)。60%遮荫条件下的灰木莲幼苗叶片具有显著较高的Narea及Amax。全光处理下的灰木莲幼苗叶片的Amax及Narea与90%遮荫处理下的灰木莲幼苗没有显著差异。
| 遮荫程度 Shading levels |
光饱和净光合速率 Amax/(μmol m-2 s-1) |
单位面积叶片氮含量 Narea/(g/m2) |
光合氮利用效率 PNUE/(μmol g-1 s-1) |
| 全光No shading | 4.07±0.36b | 2.60±0.25b | 1.67±0.31a |
| 60%遮荫60% shading | 6.03±0.56a | 3.65±0.29a | 1.68±0.18a |
| 90%遮荫90% shading | 3.50±0.46b | 2.33±0.39b | 1.76±0.36a |
| F | 8.051** | 4.762* | 0.024 |
| 统计不同遮荫条件下的各性状差异性(均值±标准误)并做方差分析(n=5), 小写字母表示差异在P < 0.05水平上显著。F值及标注分别代表*P < 0.05, **P < 0.01, | |||
由不同遮荫条件下灰木莲幼苗叶片的光合参数(表 2)可以看出, 不同遮荫处理并没有显著影响灰木莲幼苗叶片的Ci和Cc, 90%遮荫条件下的灰木莲幼苗叶片gs和gm低于其他两种处理。60%遮荫条件下的灰木莲幼苗叶片具有显著较高的Vcmax, 比全光条件下的灰木莲幼苗高出37.0%, 是90%遮荫条件下灰木莲幼苗叶片Vcmax的2倍多。全光条件下灰木莲幼苗叶片具有最高的Jmax, 但与60%遮荫处理的灰木莲幼苗无显著差异。90%遮荫条件下的灰木莲幼苗叶片Jmax显著低于其他两种处理。
| 遮荫程度 Shading levels |
气孔导度 gs/ (molCO2 m-2 s-1) |
叶肉细胞导度 gm/ (molCO2 m-2 s-1) |
胞间CO2浓度 Ci/ (μmol/mol) |
羧化位CO2浓度 Cc/ (μmol/mol) |
最大羧化速率 Vcmax/ (μmol m-2 s-1) |
最大电子传递速率 Jmax/ (μmol m-2 s-1) |
| 全光No shading | 0.046±0.005ab | 0.051±0.005ab | 289.56±6.44a | 208.65±8.46a | 24.03±0.98b | 63.85±3.71a |
| 60%遮荫60% shading | 0.058±0.006a | 0.059±0.007a | 274.70±7.21a | 187.01±7.72a | 32.93±0.94a | 61.83±4.01a |
| 90%遮荫90% shading | 0.039±0.003b | 0.039±0.004b | 285.78±5.88a | 205.35±8.16a | 16.29±1.98c | 36.68±3.84b |
| F | 3.743 | 2.394 | 1.852 | 2.523 | 31.958*** | 15.395*** |
| 统计不同遮荫条件下的各性状差异性(均值±标准误)并做方差分析(n=5), 小写字母表示差异在P < 0.05水平上显著; F值及标注分别代表*P < 0.05, **P < 0.01, ***P < 0.001。CO2导度相关数据测定条件为饱和光强及空气CO2浓度为380 μmol/mol | ||||||
不同遮荫条件下灰木莲幼苗叶片氮分配到光合系统的比例(PP)远高于分配到细胞壁中的比例(PCW)。其中, 全光条件下的灰木莲幼苗叶片PP是其PCW的3.3倍, 60%遮荫条件下为5.8倍, 90%遮荫条件下上升到6.0倍。90%遮荫处理下的灰木莲幼苗叶片PP显著高于其他两种处理, 其次为60%遮荫及全光处理。但不同的遮荫处理下PCW并没有显著差异。三个处理下灰木莲幼苗叶片中剩余组分的氮比例(POther, 叶片中除了光合系统氮及细胞壁氮之外的氮比例)高于其相应处理的PP及PCW, 不同遮荫处理的POther大小顺序为:全光(0.755 g/g)>60%遮荫(0.683 g/g)>90%遮荫(0.596 g/g)。
将光合系统细分为Rubisco、生物力能学组分及捕光组分后, 再对不同遮荫条件下灰木莲幼苗叶片的氮分配进行比较(图 2)。三个处理下的灰木莲幼苗叶片都是PL最高, 其次为PR和PB。三个处理下的灰木莲幼苗叶片PR和PB并没有显著差异, 但遮荫则显著增加了叶片分配到捕光组分中氮的比例, 三个处理下的PL大小顺序为:90%遮荫(0.296 g/g)>60%遮荫(0.216 g/g)>全光(0.132 g/g)。

|
| 图 2 不同遮荫条件下灰木莲幼苗叶片氮在Rubisco、生物力能学组分及捕光组分中的分配比例 Fig. 2 Nitrogen allocation proportion of Rubisco, bioenergetics and light-harvesting components in Manglietia glauca seedling leaves of different shading levels |
不同遮荫条件下的灰木莲叶片PR及PB都与PNUE有着显著的正相关关系(图 3)。但PL及PP与PNUE并没有显著的相关关系。

|
| 图 3 不同遮荫条件下灰木莲幼苗叶片光合系统氮分配比例(PP)、捕光组分氮分配比例(PL)、Rubisco氮分配比例(PR)及生物力能学组分氮分配比例(PB)与光合氮利用效率(PNUE)的回归分析 Fig. 3 Regression analysis of fraction of leaf nitrogen allocated to photosynthetic apparatus (PP), light-harvesting components (PL), Rubisco (PR) and bioenergetics (PB) with photosynthetic nitrogen-use efficiency (PNUE) in Manglietia glauca seedling leaves of different shading levels |
羧化位CO2浓度及光合酶的活性通过影响叶片的光合能力进而影响PNUE[43-44]。不同程度的遮荫处理并没有显著影响灰木莲幼苗叶片的CC。Yamori等对烟草(Nicotiana tabacum)[44]及Monti等对埃塞尔比亚芥(Brassica carinata)[45]的研究同样表明, 不同生长光照对CC并没有显著影响。较高程度的遮荫处理降低了灰木莲幼苗叶片的Vcmax及Jmax, 这也被Delagrange对不同光梯度下糖枫(Acer saccharum)及加拿大黄桦(Betula alleghaniensis)的研究所证明[10]。全光及60%遮荫处理下灰木莲幼苗叶片的Jmax并没有显著变化, 但60%遮荫处理下的灰木莲幼苗叶片的Vcmax显著高于全光条件下的灰木莲幼苗叶片, 因此60%遮荫处理下灰木莲幼苗叶片的Amax显著高于全光条件下的。这种现象可能与高光照下灰木莲叶片的光合机构发生光损伤有关[46-47]。而90%遮荫处理下的灰木莲幼苗叶片Vcmax及Jmax都显著低于其他两个处理, 因此Amax最低。叶片光合能力的提高可以显著提高植物的生长速度[48], 因此在培育灰木莲幼苗时要进行适度遮荫, 进行纯林改造时开出的林窗不宜过大, 要选择较为荫蔽的林下环境进行栽植。
遮荫处理同时也对Narea产生较显著的影响, 灰木莲幼苗叶片的Narea在60%遮荫条件下最高, 达到了3.65 g/m2, 而在全光下及90%遮荫下的Narea都较低。许多研究已经证明叶片氮含量会受到生长光强的影响。一般来说, 较高生长光强处理下的植物其Narea也较高[12, 14], 从而可以获得更高的光合能力, 但同样有研究表明全光处理下的植物Narea会低于其他遮荫处理[15], 并认为这种现象与植物本身的光合生理特性有关。正是由于灰木莲幼苗叶片的Amax及Narea随着生长光照的变化而同步变化, 因此不同遮荫处理下的灰木莲幼苗叶片PNUE并没有显著差异, 这也表明灰木莲幼苗叶片在不同的生长光强下光合作用对氮的利用能力相近。Wyka等对四种阔叶落叶树种及七种常绿针叶树种的研究[12], 以及Montgomery对不同光照梯度下三种树种(Dipteryx panamensis, Virola koschnyii, Brosimum alicastrum)的研究[13]也得出相同的结论。但也有研究认为生长光强的降低可以降低PNUE[14], 或者提高PNUE[15]。还有研究证明生长光照的作用与季节相关, Muller等对不同季节在不同遮荫处理下培养的常绿植物桃叶珊瑚(Aucuba japonica)的研究表明, 在夏天高光照培养的苗木PNUE较高, 而冬天则是低光照培养的较高[49]。
叶片中的氮并不全参与光合作用, 最近的研究表明光合系统的氮分配比例, 特别是光合系统中Rubisco氮比例是影响PNUE的关键因素[17, 50], 这也与本次研究的结果相同(图 3)。Rubisco是C3植物体内数量最多的蛋白质[51], 并且是光合作用的限制因素[36], 因此提高其含氮量占叶片中总氮的比例可以显著增加PNUE。虽然随着遮荫程度的增加, 灰木莲幼苗叶片光合系统氮分配比例显著增加, 但主要增加捕光组分中的氮分配比例, Rubisco及生物力能学组分氮分配比例并没有发生显著变化, 因此PNUE并没有发生显著变化。Delagrange的研究同样发现在生长光强增加后, 加拿大黄桦中的PR和PB无显著变化[10]。Feng对两种泽兰属植物(Eupatorium adenophorum及E. japonicum)的研究得出了相同的结论[11]。但Le等对桃子(Prunus persica)的研究[7]、Osada等对库页蓼(Polygonum sachalinense)的研究[8]及Chen等对三七(Panax notoginseng)的研究表明[9], 遮荫会降低PR和PB。低光环境下植物需要提高PL增加捕光能力, 而此时叶片的羧化能力及电子传递能力可能并不限制叶片光合能力, 因此有可能会减少分配到其中的氮比例;高光照环境下则可能减少PL, 增加PR和PB以提高PNUE[9]。灰木莲的PR和PB在不同光照环境下的可塑性较低, 这也限制了其提高PNUE的能力。
叶片内部各含氮组分也可能相互影响。Feng等[16]对紫茎泽兰(E. adenophorum)的研究认为叶片氮分配到光合系统与细胞壁中的比例存在一种权衡关系, 即其中一个氮分配比例增加, 则另一个氮分配比例减少。但其研究中紫茎泽兰叶片氮分配到光合系统的比例达到0.57—0.65 g/g, 细胞壁氮比例也达到了0.065—0.124 g/g, 而本研究中灰木莲的光合系统氮比例仅为0.19—0.35 g/g, 细胞壁氮比例只有0.047—0.058 g/g, 二者占的比例较小, 因此可能并不会对叶片中的氮进行竞争。所以此次研究中虽然灰木莲幼苗叶片分配到光合系统的氮比例随着遮荫程度的增加显著增加, 但分配到细胞壁的氮比例并无显著变化(图 1)。Onoda等[17]与Takashima等[52]学者认为Rubisco氮与细胞壁氮也存在一种权衡关系, 但也有Harrison[53]与Hikosaka[54]等学者认为这种关系仅存在单种植物叶片氮营养不足的条件下。由于此次试验中所有处理下灰木莲幼苗叶片POther都较高, 因此可以认为叶片中氮营养十分充足, 权衡关系形成的条件可能并不充分, 因此此次研究中的PR与PCW并没有随着光环境的变化而变化。而在营养缺乏的条件下灰木莲幼苗叶片氮分配的变化及各氮库相互关系则需要进一步的研究。此外, 研究中光合系统的氮比例随着遮荫程度的增加而增加主要是由于PL的增加所导致, 而PR及PB都并未发生较明显的变化, 这几部分氮比例并没有相互影响, 因此PL增加的部分来源于叶片的剩余组分(POther)。因此在育苗及林分改造时对灰木莲幼苗进行遮荫的同时也要适度增施氮肥, 以补充因捕光组分氮比例提高而造成的叶片氮消耗, 防止由于捕光组分氮供应不足而造成的生长不良。

|
| 图 1 不同遮荫条件下灰木莲幼苗叶片氮在光合系统、细胞壁及剩余组分中的分配比例 Fig. 1 Nitrogen allocation proportion of photosynthetic system, cell wall and other parts in Manglietia glauca seedling leaves of different shading levels |
| [1] | Hikosaka K. Interspecific difference in the photosynthesis-nitrogen relationship:patterns, physiological causes, and ecological importance. Journal of Plant Research, 2004, 117(6): 481–494. DOI:10.1007/s10265-004-0174-2 |
| [2] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osad N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [3] | Hikosaka K. Mechanisms underlying interspecific variation in photosynthetic capacity across wild plant species. Plant Biotechnology, 2010, 27(3): 223–229. DOI:10.5511/plantbiotechnology.27.223 |
| [4] | Hikosaka K. Optimal nitrogen distribution within a leaf canopy under direct and diffuse light. Plant, Cell & Environment, 2014, 37(9): 2077–2085. |
| [5] | Wright I J, Reich P B, Cornelissen J H C, Falster D S, Garnier E, Hikosaka K, Lamont B B, Lee W, Oleksyn J, Osaka N, Poorter H, Villar R, Warton D I, Westoby M. Assessing the generality of global leaf trait relationships. New Phytologist, 2005, 166(2): 485–496. DOI:10.1111/j.1469-8137.2005.01349.x |
| [6] | Niinemets V, Tenhunen J D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant, Cell & Environment, 1997, 20(7): 845–866. |
| [7] | Le Roux X, Walcroft A S, Sinoquet H, Chaves M M, Rodrigues A, Osorio L. Photosynthetic light acclimation in peach leaves:importance of changes in mass:area ratio, nitrogen concentration, and leaf nitrogen partitioning. Tree Physiology, 2001, 21(6): 377–386. DOI:10.1093/treephys/21.6.377 |
| [8] | Osada N, Onoda Y, Hikosaka K. Effects of atmospheric CO2 concentration, irradiance, and soil nitrogen availability on leaf photosynthetic traits of Polygonum sachalinense around natural CO2 springs in northern Japan. Oecologia, 2010, 164(1): 41–52. DOI:10.1007/s00442-010-1635-z |
| [9] | Chen J W, Kuang S B, Long G Q, Meng Z G, Li L G, Chen Z J, Zhang G H, Yang S C. Steady-state and dynamic photosynthetic performance and nitrogen partitioning in the shade-demanding plant Panax notoginseng under different levels of growth irradiance. Acta Physiologiae Plantarum, 2014, 36(9): 2409–2420. DOI:10.1007/s11738-014-1614-9 |
| [10] | Delagrange S. Light-and seasonal-induced plasticity in leaf morphology, N partitioning and photosynthetic capacity of two temperate deciduous species. Environmental and Experimental Botany, 2011, 70(1): 1–10. DOI:10.1016/j.envexpbot.2010.05.008 |
| [11] | Feng Y L. Photosynthesis, nitrogen allocation and specific leaf area in invasive Eupatorium adenophorum and native Eupatorium japonicum grown at different irradiances. Physiologia Plantarum, 2008, 133(2): 318–326. DOI:10.1111/j.1399-3054.2008.01072.x |
| [12] | Wyka T P, Oleksyn J, ytkowiak R, Karolewski P, Jagodziński A M, Reich P B. Responses of leaf structure and photosynthetic properties to intra-canopy light gradients:a common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species. Oecologia, 2012, 170(1): 11–24. DOI:10.1007/s00442-012-2279-y |
| [13] | Montgomery R. Relative importance of photosynthetic physiology and biomass allocation for tree seedling growth across a broad light gradient. Tree Physiology, 2004, 24(2): 155–167. DOI:10.1093/treephys/24.2.155 |
| [14] | 张亚杰, 冯玉龙. 喜光榕树和耐荫榕树光适应机制的差异. 植物生理与分子生物学学报, 2004, 30(3): 297–304. |
| [15] | 王松, 蔡艳飞, 李枝林, 李世峰. 光照条件对高山杜鹃光合生理特性的影响. 西北植物学报, 2012, 32(10): 2095–2101. DOI:10.3969/j.issn.1000-4025.2012.10.025 |
| [16] | Feng Y L, Lei Y B, Wang R F, Callaway R M, Valiente-Banuet A, Inderjit, Li Y P, Zheng Y L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1853–1856. DOI:10.1073/pnas.0808434106 |
| [17] | Onoda Y, Hikosaka K, Hirose T. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Functional Ecology, 2004, 18(3): 419–425. DOI:10.1111/fec.2004.18.issue-3 |
| [18] | Li Z A, Peng S L, Rae D J, Zhou G Y. Litter decomposition and nitrogen mineralization of soils in subtropical plantation forests of southern china, with special attention to comparisons between legumes and non-legumes. Plant and Soil, 2001, 229(1): 105–116. DOI:10.1023/A:1004832013143 |
| [19] | 邹碧, 李志安, 丁永祯, 谭万能. 南亚热带4种人工林凋落物动态特征. 生态学报, 2006, 26(3): 715–721. |
| [20] | 刘丽, 段争虎, 汪思龙, 胡江春, 胡治刚, 张倩茹, 王书锦. 不同发育阶段杉木人工林对土壤微生物群落结构的影响. 生态学杂志, 2009, 28(12): 2417–2423. |
| [21] | Zhou T, Sheng W T. On the plantation sustainability in China. World Forestry Research, 2008, 21(3): 49–53. |
| [22] | 何友均, 梁星云, 覃林, 李智勇, 邵梅香, 谭玲. 南亚热带人工针叶纯林近自然改造早期对群落特征和土壤性质的影响. 生态学报, 2013, 33(8): 2484–2495. |
| [23] | 王卫霞, 史作民, 罗达, 刘世荣, 卢立华. 南亚热带3种人工林土壤微生物生物量和微生物群落结构特征. 应用生态学报, 2013, 24(7): 1784–1792. |
| [24] | 卢志芳, 卢立华, 覃书源. 城市绿化树种——灰木莲. 中国城市林业, 2006, 4(5): 63. |
| [25] | 韦善华, 覃静, 朱贤良, 秦武明, 邓力, 张丽琼. 南宁地区灰木莲人工林生长规律研究. 西北林学院学报, 2011, 26(5): 174–178. |
| [26] | 乔梦吉. 广西优良珍贵树种灰木莲的组织培养. 南方农业学报, 2013, 44(6): 989–993. |
| [27] | 杨耀海, 刘明义, 常森有. 灰木莲引种栽培试验研究. 西南林学院学报, 2007, 27(3): 29–32, 51. |
| [28] | 李俊贞, 覃静, 邓力, 秦武明, 黄建友, 韦鹏飞. 盐分胁迫对灰木莲苗木生长及生理指标的影响. 西部林业科学, 2011, 40(2): 40–43. |
| [29] | 郭淑红, 薛立, 张柔, 杨振意. 4种幼苗对低温胁迫的光合生理响应. 华南农业大学学报, 2012, 33(3): 373–377. |
| [30] | 陈伟光, 张卫强, 李召青, 曾令海, 周平, 周毅, 陈光胜. 木荷等6种阔叶树种光合生理特性比较. 广东林业科技, 2010, 26(4): 12–17. |
| [31] | Monclus R, Dreyer E, Villar M, Delmotte F M, Delay D, Petit J M, Barbaroux C, Le Thiec D, Bréchet C, Brignolas F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides×Populus nigra. New Phytologist, 2006, 169(4): 765–777. DOI:10.1111/nph.2006.169.issue-4 |
| [32] | Broeckx L S, Fichot R, Verlinden M S, Ceulemans R. Seasonal variations in photosynthesis, intrinsic water-use efficiency and stable isotope composition of poplar leaves in a short-rotation plantation. Tree Physiology, 2014, 34(7): 701–715. DOI:10.1093/treephys/tpu057 |
| [33] | 冯秋红, 程瑞梅, 史作民, 刘世荣, 王卫霞, 刘兴良, 何飞. 四川巴郎山齿果酸模叶片氮素及其分配的海拔响应. 植物生态学报, 2013, 37(7): 591–600. |
| [34] | Harley P C, Loreto F, Di Marco G, Sharkey T D. Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2. Plant Physiology, 1992, 98(4): 1429–1436. DOI:10.1104/pp.98.4.1429 |
| [35] | Loreto F, Di Marco G Tricoli D, Sharkey T D. Measurements of mesophyll conductance, photosynthetic electron transport and alternative electron sinks of field grown wheat leaves. Photosynthesis Research, 1994, 41(3): 397–403. DOI:10.1007/BF02183042 |
| [36] | Farquhar G D, Van Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149(1): 78–90. DOI:10.1007/BF00386231 |
| [37] | Loustau D, Brahim M B, Gaudillère J P, Dreyer E. Photosynthetic responses to phosphorus nutrition in two-year-old maritime pine seedlings. Tree Physiology, 1999, 19(11): 707–715. DOI:10.1093/treephys/19.11.707 |
| [38] | Johnson F H, Eyring H, Williams R W. The nature of enzyme inhibitions in bacterial luminescence:sulfanilamide, urethane, temperature and pressure.. Journal of Cellular Physiology, 1942, 20(3): 247–268. DOI:10.1002/(ISSN)1553-0809 |
| [39] | Harley P C, Thomas R B, Reynolds J F, Strain B R. Modelling photosynthesis of cotton grown in elevated CO2. Plant, Cell & Environment, 1992, 15(3): 271–282. |
| [40] | Jordan D B, Ogren W L. The CO2/O2 specificity of ribulose 1, 5-bisphosphate carboxylase/oxygenase:Dependence on ribulosebisphosphate concentration, pH and temperature. Planta, 1984, 161(4): 308–313. DOI:10.1007/BF00398720 |
| [41] | Nolan W G, Smillie R M. Multi-temperature effects on hill reaction activity of barley chloroplasts. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1976, 440(3): 461–475. DOI:10.1016/0005-2728(76)90034-7 |
| [42] | 史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909–5919. |
| [43] | Kositsup B, Montpied P, Kasemsap P, Thaler P, Améglio T, Dreyer E. Photosynthetic capacity and temperature responses of photosynthesis of rubber trees (Hevea brasiliensis, Müll. Arg.) acclimate to changes in ambient temperatures. Trees, 2009, 23(2): 357–365. DOI:10.1007/s00468-008-0284-x |
| [44] | Yamori W, Evans J R, Von Caemmerer S. Effects of growth and measurement light intensities on temperature dependence of CO2 assimilation rate in tobacco leaves. Plant, Cell & Environment, 2010, 33(3): 332–343. |
| [45] | Monti A, Bezzi G, Venturi G. Internal conductance under different light conditions along the plant profile of Ethiopian mustard (Brassica carinata A. Brown.). Journal of Experimental Botany, 2009, 60(8): 2341–2350. DOI:10.1093/jxb/erp032 |
| [46] | Katahata S, Naramoto M, Kakubari Y, Mukai Y. Photosynthetic acclimation to dynamic changes in environmental conditions associated with deciduous overstory phenology in Daphniphyllum humile, an evergreen understory shrub. Tree Physiology, 2005, 25(4): 437–445. DOI:10.1093/treephys/25.4.437 |
| [47] | Zhou Y, Lam H M, Zhang J. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice. Journal of Experimental Botany, 2007, 58(5): 1207–1217. DOI:10.1093/jxb/erl291 |
| [48] | Hua Q, Ying C, Yan X, Yao Y H, An S Q. Leaf nitrogen partition between photosynthesis and structural defense in invasive and native tall form spartina alterniflora populations:effects of nitrogen treatments. Biological Invasions, 2012, 14(10): 2039–2048. DOI:10.1007/s10530-012-0210-4 |
| [49] | Muller O, Oguchi R, Hirose T, Werger M J A, Hikosaka K. The leaf anatomy of a broad-leaved evergreen allows an increase in leaf nitrogen content in winter. Physiologia Plantarum, 2009, 136(3): 299–309. DOI:10.1111/ppl.2009.136.issue-3 |
| [50] | Hikosaka K, Shigeno A. The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity. Oecologia, 2009, 160(3): 443–451. DOI:10.1007/s00442-009-1315-z |
| [51] | Evans J R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia, 1989, 78(1): 9–19. DOI:10.1007/BF00377192 |
| [52] | Takashima T, Hikosaka K, Hirose T. Photosynthesis or persistence:nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant, Cell & Environment, 2004, 27(8): 1047–1054. |
| [53] | Harrison M T, Edwards E J, Farquhar G D, Nicotra A B, Evans J R. Nitrogen in cell walls of sclerophyllous leaves accounts for little of the variation in photosynthetic nitrogen-use efficiency. Plant, Cell & Environment, 2009, 32(3): 259–270. |
| [54] | Hikosaka K, Osone Y. A paradox of leaf-trait convergence:why is leaf nitrogen concentration higher in species with higher photosynthetic capacity?. Journal of Plant Research, 2009, 122(3): 245–251. DOI:10.1007/s10265-009-0222-z |