 2017, Vol. 37
2017, Vol. 37文章信息
- 李浩, 胡顺军, 朱海, 李茜倩
- LI Hao, HU Shunjun, ZHU Hai, LI Xiqian.
- 基于热扩散技术的梭梭树干液流特征研究
- Characterization of stem sapflow Haloxylon ammodendron by using thermal dissipation technology
- 生态学报. 2017, 37(21): 7187-7196
- Acta Ecologica Sinica. 2017, 37(21): 7187-7196
- http://dx.doi.org/10.5846/stxb201608301764
-
文章历史
- 收稿日期: 2016-08-30
- 网络出版日期: 2017-07-11





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
古尔班通古特沙漠是我国干旱区唯一以固定、半固定线形沙丘为主的沙漠, 其稳定状况直接影响着天山北麓绿洲地区的生态安全。由于人类活动和气候变化的影响, 使得荒漠化问题在这一地区日益严重, 梭梭是古尔班通古特沙漠中最主要的植被类型, 在防风固沙、遏制沙漠对绿洲的入侵中发挥着不可替代的作用。随着绿洲区地下水过度开采利用, 绿洲浅层地下水位下降, 绿洲向沙漠侧渗的地下水减少, 古尔班通古特沙漠南缘以梭梭为建群种的植物群落已表现出衰退迹象。水分作为干旱区生态环境变化的最主要最直接的影响因素, 其量的大小能否满足植物生长的需要直接决定了区域生态环境的发展, 植物的蒸腾作用是水分消耗的主要部分。树木茎干液流是由蒸腾作用所引起的树干边材的液流, 其流量占到树木蒸腾量的90%以上, 利用茎流计监测树木茎干液流能反映树木的蒸腾过程[1-2]。
对树干液流特征的研究, 一直都是生态学研究的主要内容之一[3]。在古尔班通古特沙漠边缘, 梭梭作为优势种广泛分布, 梭梭属于落叶小乔木或大灌木, 具有主干短、枝条细、分枝性强等生物特征。已有学者采用热脉冲茎流计和包裹式热平衡茎流计针对梭梭的树干液流特征及其与环境因子间的关系进行了研究, 但是热脉冲式茎流计成本过于昂贵且对液流的响应滞后[4], 茎热包裹式茎流计容易对茎干造成伤害[5], 而Granier在热脉冲法的基础改造的热扩散茎流计可以避免以上的两种方法的缺点[6-7], 更适宜开展大量的野外监测。热扩散探针法可在树木自然生长条件下连续测定树干液流量, 基本不破坏树木正常生长状态, 具有时间分辨率高, 准确度高和易操作等优点, 已成为目前树木耗水研究中最常用的研究方法之一[8-9]。
树干液流动态变化受植物生物学结构、土壤水分、气象因素影响, 生物学结构决定液流的潜在能力, 土壤供水决定液流的总体水平, 而气象因素决定液流的瞬间变动[10]。探究树干液流特征及其对环境因子的响应是准确地测算单株树木的蒸腾耗水量前提, 构建树干液流速率与环境因子的模型影响, 可以实现通过环境指示对单木蒸腾的预测。本文在对古尔班通古特沙漠南缘不同胸径梭梭茎干液流速率和环境因子进行连续监测的基础上, 研究自然条件下梭梭液流变化规律及其对环境因子的响应, 构建树干液流速率与环境因子模型, 利用尺度扩展的方法推算出梭梭群落的日、季节蒸腾耗水量, 探究梭梭的生态适应能力, 为沙丘的治理提供依据。
1 研究区概况试验地位于古尔班通古特沙漠南缘北沙窝(44°22′N, 87°55′E), 深入沙漠5 km, 距中国科学院阜康荒漠生态站直线距离15 km, 距乌鲁木齐直线距离80 km, 是新疆北部典型的沙漠生态系统, 保持着原始的沙漠植被景观。北沙窝地貌类型以南北走向的树枝状沙垄为主, 表现为固定沙丘、半固定沙丘, 其间广泛分布着由梭梭、生物结皮、以及其他沙生植物组成的小乔木群落, 梭梭是建群种。平均气温7.19℃, 极端最高气温41.5℃。多年平均降水量128.6 mm, 蒸发量为1746.4 mm。春夏降雨量约占总降水量的2/3, 秋季干燥, 冬季积雪厚度一般在20 cm以上, 积雪覆盖持续100—160 d, 春季的覆雪融水和春夏季的降雨是维持本区植被生存的主要水分来源[11]。
2 试验材料与方法 2.1 试验材料与试验地选择试验材料为梭梭(Haloxylon ammodendron)。梭梭是落叶小乔木或大灌木, 耐贫瘠、耐旱性极强, 广泛分布在荒漠地区的防风固沙林种。在北沙窝试验地选取一块地势相对平坦开阔的林地安放波文比气象观测系统, 以波文比为中心拉20 m×20 m左右的梭梭林样地, 自然生长密度0.25株/m2, 平均基径3.3 cm, 平均高度192 cm, 平均冠幅199 cm×192 cm, 植被盖度20%—30%。安插探针的6棵样木距离波文比3—5 m左右, 使用游标卡尺测得基径分别为4.55、8.91、8.44、9.55、8.28、6.49 cm。采用植物生长锥对6棵样木边材厚度进行测量, 发现梭梭边材和心材具有颜色差异, 边材厚度分别为1.1、1.8、1.55、1.95、1.9、1.65 cm。
2.2 茎流计安装与测定本试验选用澳达植物感应仪器公司生产的Plant Sensors PS-TDP8树木茎流监测系统, 探针长度2 cm, 梭梭边材厚度较窄, 为避免探针插入心材造成的误差和方位影响, 探针统一安装于树干北侧基部, 插入深度为1 cm, 探针距地面20—30 cm。TDP探针用泡沫软塑料包裹, 再包上锡纸和塑料纸, 防止热辐射和雨水的浸入而干扰数据。探针通过电缆和数据自动采集仪进行数据记录和储存, 每30 s获取1次数据, 每30 min进行平均并存储数据。
2.3 环境因子测定环境因子采用全自动波文比综合观测系统监测, 该系统采用新型一体化结构设计, 可采集空气温度Ta(Air temperature)、相对湿度RH(Relative humidity)、风速WS(Wind speed)、净辐射TBB(Net radiation)、实际水汽压e(Vapor pressure)、土壤温度Ts(Soil temperature)、土壤含水率SWC(Soil water condition)等信息, 饱和水汽压亏缺VPD(Vapor pressure deficit)由空气温度和空气相对湿度计算求得。设定数据采集间隔时间为10 min, 测定时间与TDP同步进行, 数据自动存储。
2.4 液流速率与林分蒸腾的计算数据采集后, 利用澳达植物感应仪器公司提供的分析软件对原始数据处理与计算。液流速率计算公式如下:
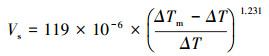
|
(1) |
其中, Vs为树干边材的液流速率(m/s), ΔTm为24 h内最大探针温差值, ΔT为两探针间的瞬时温差值, ΔT由TDP两探针所输出的电压差除以经验常数0.04所得。林分蒸腾计算公式如下:
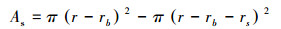
|
(2) |
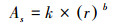
|
(3) |
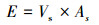
|
(4) |

|
(5) |
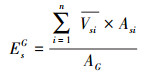
|
(6) |
式(2)是边材面积的测定公式, r是基径, rb是树皮厚度, rs是边材厚度;式(3)是边材面积与胸径的关系式, k=0.5827、b=1.9258是通过实测的边材厚度和基径线性回归方程分析得出的系数;式(4)是单树蒸腾耗水的测定公式, Vs是树干基部边材液流速率, As树干基部边材面积;式(5)林分内不同径阶样树的总蒸腾耗水测定公式, 
采用EXCEL软件对各项环境因子数据和梭梭树干边材液流速率数据进行预处理, 采用SPSS软件进行多元回归分析, 得到梭梭液流速率与气象因子及土壤环境因子之间的统计关系, 采用Origin软件作图。SPSS进行回归分析中通径系数为直接通径系数, 是用标准回归系数来表示。
3 结果与分析 3.1 梭梭树干液流速率的变化规律梭梭树干边材液流速率具有明显的昼夜节律变化和季节变化规律。在春季和秋季, 晴朗无云的晴天, 梭梭树干液流的日变化曲线呈典型的“单峰型”;在夏季存在“单峰型”和“双峰型”两种变化曲线。梭梭在遭受高温和干旱胁迫的情况下, 液流的峰值会提前出现, 或者进行“午休”出现双峰波形。这种午休现象通过调整叶片气孔的闭合, 减小叶片蒸腾速率, 避免了过多的蒸腾耗水, 从而能够适应高温、干旱的气候[12]。树木边材液流时空变化特征包括启动时间、液流峰出现时间、峰值, 液流高峰持续的时间和进入低谷的时间等, 很好地反应了树木蒸腾耗水特性, 受环境因子的作用, 不同季节间的差异很大。如图 1所示, 夏季7月份液流的启动时间最早, 持续时间最长, 峰值最大, 日树干液流量最大, 液流启动在7:00—8:00之间, 10:00—11:00左右出现波峰, 最大液流速率在0.75—0.85 dm/h之间, 0:00左右停止。春季液流启动时间在8:30—9:00之间, 12:30—13:00之间达到最高值, 23:30—0:00之间停止, 最大液流速率约为0.5 dm/h。秋季液流启动时间最晚, 峰值最小, 启动时间在9:30—10:00之间, 13:30—14:00之间出现峰值, 1:00左右停止, 最大液流速率约为0.35 dm/h。春季和秋季, 在夜间液流基本停止;而在夏季, 由于高温和干旱的胁迫, 白天强烈的蒸腾致使树体水分损失, 夜间树干出现微弱的液流对树体水分进行补充[13]。

|
| 图 1 样树树干液流速率不同月份的日变化与不同季节的连续日变化 Fig. 1 The diurnal changein different months and consecutive days change of sap flow velocity in different seasons |
由于试验地所处的地理环境限制, 浅层地下水埋深9 m左右, 150 cm以下的土壤含水率日变化极小、季节变化不显著[14], 梭梭树干液流瞬时速率与土壤含水率的相关性不显著。因此, 梭梭树干液流速率日变化主要受气象因素(风速、净辐射、实际水汽压、饱和水汽压亏缺值、空气相对湿度和空气温度)的影响。分别在5月、7月和9月3个月份, 选择连续5个晴天内的树干液流速率数据与相应的气象数据进行相关性分析, 结果如表 1和图 2所示。在整个生长季, 梭梭树干液流速率与风速、净辐射、空气温度、饱和水汽压亏缺值等因素呈显著正相关, 与实际水汽压和空气湿度呈极显著负相关。, 在春季影响梭梭树干液流速率日变化的主要影响因子是饱和水汽压亏缺值、空气温度和净辐射, 风速对液流速率的影响最小;在夏季影响梭梭树干液流速率日变化的因子是空气湿度、空气温度和净辐射, 风速对液流速率的影响最小;在秋季影响梭梭树干液流速率日变化的因子是饱和水汽压亏缺值、净辐射, 实际水汽压对瞬时液流速率变化的影响最小。由此可见, 在整个生长季中, 饱和水汽压亏缺值、空气温度和净辐射是导致树干瞬时液流速率变化的关键因子。
| 项目 Items |
风速/(m/s) Wind speed |
净辐射/(w/s) Net radiation |
水汽压/kPa Vapor pressure |
水汽压亏缺/kPa Vapor pressure deficit |
空气湿度/% Relative humidity |
空气温度/℃ Air temperature |
|
| 春季Spring | 相关系数 | 0.157** | 0.813** | -0.540** | 0.664** | -0.673** | 0.696** |
| 通径系数 | -0.040 | 0.593 | -0.537 | -1.856 | 0.141 | 1.910 | |
| 夏季Summer | 相关系数 | 0.591** | 0.861** | -0.224** | 0.441** | -0.449** | 0.488** |
| 通径系数 | 0.016 | 0.865 | -0.453 | -2.315 | 0.204 | 2.051 | |
| 秋季Autumn | 相关系数 | 0.598** | 0.789** | -0.525** | 0.808** | -0.763** | 0.784** |
| 通径系数 | 0.097 | 0.463 | 0.001 | 0.691 | 0.108 | -0.075 | |
| **表示在0.01水平上显著相关 | |||||||

|
| 图 2 梭梭树干液流速率与主要气象因子间的关系 Fig. 2 The relationship between the meteorological factors and the 30-min averages of stem sap flow of Haloxylon ammodendron a—e分别代表风速、空气湿度、实际水汽压、空气温度和净辐射 |
梭梭树干液流速率瞬时变化主要受气象因子的影响, 但是更大时间尺度上的变化受树体自身的生理结构和功能、气象环境因子和土壤环境因子共同作用影响[15-16]。不同的时间和空间尺度, 影响树木蒸腾耗水的主导因素不同, 土壤水分状况和土壤温度在以日为单元的时间尺度上对树干液流速率的变化影响微乎其微, 但在以季节为单位的时间尺度上土壤温度和土壤水分含量与日均液流速率有显著的相关性[17-18]。因此, 在研究梭梭树干液流速率较大时间尺度上的变化时, 需要考虑土壤环境因子对树木蒸腾量的影响。
对生长季内30、70 cm和110 cm土壤日均含水率变化进行比较分析, 结果如图 3所示。受积雪消融和降雨影响, 30 cm和70 cm的土壤含水量在5月初最高, 110 cm土壤含水量较低, 随着水分下渗和蒸散发的进行, 表层土壤的含水率逐渐减少, 深层土壤含水率逐渐增加, 不同深度间土壤含水率差异逐渐缩小。进入夏季, 表层30 cm土壤含水量小于深层的土壤含水率, 秋季降雨增多, 受雨水和凝结水的影响, 表层土壤的含水率回升, 出现第二个峰值。有关研究表明梭梭根系分布具有分层现象, 分为浅层根系和深层根系, 深层根系可达2 m以下[19]。在整个生长季, 110 cm深土壤的含水率大体呈先增大后减小的趋势, 与生长季梭梭树干液流速率的变化趋势相一致, 如表 3所示。在春季和秋季110 cm处的日平均土壤含水率与液流速率具有极显著的正相关性, 在夏季与液流速率相关性不显著。在阴雨天, 大气温度、净辐射强度降低, 空气相对湿度和水汽压升高, 梭梭树干液流速率降低, 且远远小于晴天的液流速率, 导致生长季的日均液流速率曲线出现了数次较大幅度的波动。

|
| 图 3 整个生长季梭梭树干液流日均速率与主要环境因子间的关系 Fig. 3 The relationship between the main environmental factors and daily average rate of stem sap flow |
| 季节Season 深度Depth/cm |
春季Spring | 夏季Summer | 秋季Autumn | |||||||||||
| 30 | 70 | 110 | 150 | 30 | 70 | 110 | 150 | 30 | 70 | 110 | 150 | |||
| 相关系数 Correction coefficient |
-0.722** | -0.812** | 0.793** | 0.729** | 0.08 | 0.056 | -0.085 | -0.095 | 0.892** | 0.902** | 0.936** | 0.949** | ||
| 显著性Significance | 0 | 0 | 0 | 0 | 0.669 | 0.763 | 0.651 | 0.612 | 0 | 0 | 0 | 0 | ||
| 自由度Freedom | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | ||
以生长季内5—9月152 d的观测数据为基础, 对日均液流速率和气象因子、土壤含水率进行相关性分析。如图 3和表 2所示, 梭梭日均液流速率与净辐射、空气温度、VPD、实际水汽压、土壤含水率和土壤温度等呈极显著的正相关性;与空气湿度等呈极显著的负相关性;而与风速的相关性不显著。从通径系数来看, 影响梭梭树干液流速率季节变化的关键因子是净辐射、VPD和空气温度。
| 项目Items | 风速 Wind speed/ (m/s) |
净辐射 Net solar radiation/ (w/s) |
水汽压/kPa Vapor pressure |
水汽压亏缺/kPa Vapor pressure deficit |
空气湿度/% Relative humidity |
空气温度 Air temperature/ ℃ |
土壤含水率 Soil water condition/% |
土壤温度 Soil temperature/℃ |
| 相关系数 Correction coefficients |
0.116 | 0.646** | 0.402** | 0.568** | -0.319** | 0.731** | 0.379** | 0.597** |
| 显著性Significance | 0.156 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| 通径系数Path coefficients | 0.128 | 0.496 | -0.329 | -0.750 | 0.293 | 1.325 | 0.106 | 0.108 |
在长期连续对梭梭树干液流速率的系统观测基础上, 分别以3个季节中连续5 d的瞬时树干液流速率为因变量, 以瞬时气象数据为自变量进行逐步回归, 得到不同季节树干液流速率和气象因子的回归方程, 如式(7)—(9)所示。以整个生长季的树干液流速率日均值为因变量, 以气象因子和土壤因子的数据日均值为自变量进行逐步回归分析, 得到生长季树干液流速率与环境因子的回归方程, 如式(10)所示。
春季:
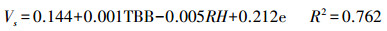
|
(7) |
夏季:
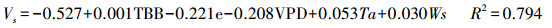
|
(8) |
秋季:
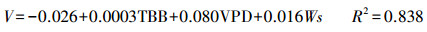
|
(9) |
生长季:
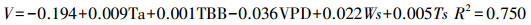
|
(10) |
式(7)—(9)是不同季节的瞬时速率回归方程, 式(10)是整个生长季日均速率的回归方程。其中, Vs为梭梭树干液流的瞬时速率(dm/h);V为梭梭树干液流的日均速率(dm/h);TBB为净辐射强度;RH为空气相对湿度;e为水汽压;WS为风速;VPD为饱和水汽压亏缺;Ts为土壤温度。以上4个方程式的F检验都达到了0.01水平上显著, 复相关系数R2均在0.75以上, 拟合效果较好。秋季的日耗水模型的模拟效果优于春季和夏季, 季节耗水模型的复相关系数略小于日周期耗水模型。建立日周期模型的数据全部选自不同季节连续晴天的数据, 不受天气条件差异的影响, 建立季节耗水模型则受阴雨天气数据的影响, 导致季节模型模拟效果略差于日周期的模型。
3.5 梭梭树群落的季节蒸腾耗水量特征利用热扩散茎流计数据依靠尺度扩展的方法, 计算的除了生长季内不同时间的林分耗水总量和蒸腾总量变化情况如图 4和表 4所示。整个生长季林分的日蒸腾呈先增大后减小的趋势, 与110 cm深处土壤含水率的变化规律相一致, 从五月上旬的日蒸腾为0.21 mm/d, 在六月中旬到达峰值0.45 mm/d, 峰值一直持续40 d左右, 从7月下旬开始下降, 九月下旬林分日蒸腾降到0.10 mm/d。从表 4可以看出, 20 m×20 m的样方内整个生长季的全部耗水量达到4428.924 L, 5月耗水736.361 L、6月耗水1073.685 L、7月耗水1145.842 L、8月耗水9117.580 L、9月耗水511.651 L, 152 d的平均耗水量为29.137 L/d。

|
| 图 4 生长季的林分日蒸腾与110 cm土壤含水率的的动态变化 Fig. 4 The dynamic changes of transpiration and soil moisture content at 110 cm depth |
| 月份Month | 日期Date | 日耗水 Daily water consumption/(mL/d) |
日蒸腾 Daily transpiration/mm |
| 5 | 1—10 | 17654.637 | 0.214 |
| 11—20 | 24887.710 | 0.302 | |
| 21—31 | 28718.391 | 0.348 | |
| 6 | 1—10 | 34380.693 | 0.417 |
| 11—20 | 35893.513 | 0.435 | |
| 21—30 | 37094.278 | 0.450 | |
| 7 | 1—10 | 37050.954 | 0.449 |
| 11—20 | 37037.030 | 0.449 | |
| 21—31 | 36799.939 | 0.446 | |
| 8 | 1—10 | 33052.926 | 0.401 |
| 11—20 | 29050.286 | 0.352 | |
| 21—30 | 29654.766 | 0.360 | |
| 9 | 1—10 | 26449.559 | 0.321 |
| 11—20 | 18185.863 | 0.221 | |
| 21—29 | 8293.946 | 0.101 |
梭梭树干液流速率的日变化曲线主要呈单峰型, 夏季偶有双峰型, 夜间液流微弱, 昼夜差异显著;梭梭树干液流的日变化格型和最高峰值有明显的季节差异, 夏季液流启动时间、峰值出现时间较早。曹晓明[11]、丁访军[20]、龚道枝[21]等的研究结果表明, 树干液流变化存在明显的昼夜节律, 不同季节液流差异显著, 夏季的蒸腾速率大于春季和秋季, 与本研究结果相一致。孙鹏飞等[22]在古尔班通古特沙漠南缘基于热脉冲技术对梭梭树干液流的研究结果表明, 5—9月份液流量昼夜变化不显著, 液流变化曲线呈平缓波动的多峰型, 液流量逐日降低, 与本研究结果不符。孙鹏飞等的试验区域地下水埋深大于16 m, 梭梭根系不能直接利用地下浅水, 而本试验区域地下浅水位在9 m左右, 梭梭可能直接利用地下浅水, 不同的土壤水分条件和不同的树干液流监测方法可能导致了梭梭树干液流变化规律的差异。梭梭在不同的水分胁迫条件下, 液流速率变化格局存在差异, 但都能正常生长, 说明梭梭能适应不同的水分环境, 耐干旱胁迫能力强。
梭梭树干液流速率的瞬时变化主要受净辐射、空气温度和VPD的影响, 不同季节树干液流速率对气象因子的响应有一定的差异。梭梭树干瞬时液流速率与风速、净辐射、饱和水汽压亏缺值、空气温度等因素呈显著正相关关系, 与实际水汽压和空气湿度呈极显著负相关。梭梭树干日均液流速率的变化主要受净辐射、VPD和空气温度的影响, 与净辐射、空气温度、VPD、实际水汽压、土壤含水率和土壤温度等呈极显著的正相关性, 与空气湿度等呈极显著的负相关性;而与风速的相关性不显著。不同的时间和空间尺度, 不同的环境条件下影响树木蒸腾耗水的主导因素不同, 证实了树木蒸腾耗水影响和调控机制的复杂性和相对性[23]。
不同季节的多元线性回归模型的复相关系数均在0.764以上, 可以较好地模拟梭梭树干边材液流速率与气象因子的响应特征;生长季日均液流速率的模型复相关系数为0.75, 可以较好地模拟某一季度或生长季的耗水量。日均液流速率模型是以日平均气象资料为基础, 可以忽略具体天气波动的干扰, 但是减少了模型建立的数据样本, 降低了模拟的准确度。因此, 构建一套普遍适用的比较系统和科学的树木蒸腾耗水调控机制, 需要考虑在不同时间尺度上对环境因子和液流速率的关系进行模拟。
梭梭树干液流速率变化和个体蒸腾的研究是一个热点问题, 但对于梭梭的生态适应性、群落衰退原因的探讨而言, 群落的日、季节、年蒸腾耗水量更值得研究。梭梭个体耗水量测定和尺度转换过程中的空间纯量的选择, 是决定能否准确估算梭梭群落耗水量的关键。根据植物特征选择合适的个体耗水测量工具和对经验系数的校正有利于更加精确的测定个体的耗水量, 选择合适的纯量能提高试验的可操作性和尺度扩展结果的准确性。本文以热扩散茎流计作为工具, 测定梭梭个体的蒸腾量, 采用尺度扩展的方法以树干基径为纯量, 推算出试验样方内梭梭的林分日均蒸腾量是0.35 mm/d, 整个研究期间总蒸腾量是53.35 mm。在梭梭的生长期间, 古尔班通古特沙漠南缘高温干旱, 降雨量小且蒸发强, 但梭梭群落的日均蒸腾量较小, 仅达到0.35 mm/d, 可见梭梭对干旱高温的沙漠环境具有较强的生态适应能力。
| [1] | 曹文强, 韩海荣, 马钦彦, 康峰峰, 蔺琛. 山西太岳山辽东栎夏季树干液流通量研究. 林业科学, 2004, 40(2): 174–177. DOI:10.11707/j.1001-7488.20040231 |
| [2] | 马玲, 赵平, 饶兴权, 蔡锡安, 曾小平. 乔木蒸腾作用的主要测定方法. 生态学杂志, 2005, 24(1): 88–96. |
| [3] | 鲍玉海, 杨吉华, 李红云, 郑兆亮, 宗萍萍. 不同灌木树种蒸腾速率时空变异特征及其影响因子的研究. 水土保持学报, 2005, 19(3): 184–187. |
| [4] | Baker J M, Van Bavel C H M. Measurement of mass flow of water in the stems of herbaceous plants. Plant, Cell & Environment, 1987, 10(9): 777–782. |
| [5] | Lundblad M, Lagergren F, Lindroth A. Evaluation of heat balance and heat dissipation methods for sapflow measurements in pine and spruce. Annals of Forest Sciences, 2001, 58(6): 625–638. DOI:10.1051/forest:2001150 |
| [6] | Granier A. A new method of sap flow measurement in tree stems. Annales des Sciences Forestieres, 1985, 42(2): 193–200. DOI:10.1051/forest:19850204 |
| [7] | Granier A. Sap flow measurements in Douglas-fir tree trunks by means of a new thermal method. Annales des Sciences Forestieres, 1987, 44(1): 1–14. DOI:10.1051/forest:19870101 |
| [8] | Smith D M, Allen S J. Measurement of sap flow in plant stems. Journal of Experimental Botany, 1996, 47(12): 1833–1844. DOI:10.1093/jxb/47.12.1833 |
| [9] | Lu P, Urban L, Zhao P. Granier's thermal dissipation probre (TDP) method for measuring sap flow in trees: Theory and practice. Acta Botanica Sinica, 2004, 46(6): 631–646. |
| [10] | 郑阿宝, 钟育谦, 阮宏华, 姜志林. 次生栎林蒸腾强度与生态因子的关系. 植物资源与环境学报, 2000, 9(2): 27–29. |
| [11] | 曹晓明, 陈曦, 王卷乐, 王权, 王珊珊. 古尔班通古特沙漠南缘非灌溉条件下梭梭(Haloxylon ammodendron)蒸腾耗水特征. 干旱区地理, 2013, 36(2): 292–302. |
| [12] | 常学向, 赵文智, 张智慧. 荒漠区固沙植物梭梭(Haloxylon ammodendron)耗水特征. 生态学报, 2007, 27(5): 1826–1837. |
| [13] | 许浩, 张希明, 闫海龙, 孙红叶, 单立山. 塔克拉玛干沙漠腹地梭梭(Haloxylon ammodendron)蒸腾耗水规律. 生态学报, 2008, 28(8): 3713–3720. |
| [14] | 朱海, 胡顺军, 陈永宝. 古尔班通古特沙漠南缘固定沙丘土壤水分时空变化特征. 土壤学报, 2016, 53(1): 117–126. DOI:10.11766/trxb201504030679 |
| [15] | Meinzer F C, Goldstein G, Andrade J L. Regulation of water flux through tropical forest canopy trees: do universal rules apply?. Tree Physiology, 2001, 21(1): 19–26. DOI:10.1093/treephys/21.1.19 |
| [16] | McDowell N G, White S, Pockman W T. Transpiration and stomatal conductance across a steep climate gradient in the southern Rocky Mountains. Ecohydrology, 2008, 1(3): 193–204. DOI:10.1002/eco.v1:3 |
| [17] | 黄玉清, 张中峰, 何成新, 赵平, 袁维园, 焦继飞, 尤业明. 岩溶区青冈栎整树蒸腾的季节变化. 应用生态学报, 2009, 20(2): 256–264. |
| [18] | 格日乐, 乌仁陶德, 张力, 吉米斯, 刘军. 几种沙漠植物蒸腾作用特性及其环境响应机制的研究. 水土保持研究, 2007, 14(1): 184–186. |
| [19] | 郭泉水, 谭德远, 王春玲, 史作民, 马超. 肉苁蓉采挖坑对梭梭根际土壤水分的影响研究. 林业科学研究, 2005, 18(3): 315–320. |
| [20] | 丁访军, 王兵, 赵广东. 毛竹树干液流变化及其与气象因子的关系. 林业科学, 2011, 47(7): 73–81. DOI:10.11707/j.1001-7488.20110711 |
| [21] | 龚道枝, 王金平, 康绍忠, 胡笑涛, 张富仓, 李志军. 不同水分状况下桃树根茎液流变化规律研究. 农业工程学报, 2001, 17(4): 34–38. |
| [22] | 孙鹏飞, 周宏飞, 李彦, 李妙伶. 古尔班通古特沙漠原生梭梭树干液流及耗水量. 生态学报, 2010, 30(24): 6901–6909. |
| [23] | 李广德, 富丰珍, 席本野, 王烨, 贾黎明. 基于热扩散技术的三倍体毛白杨单木及林分蒸腾耗水研究. 生态学报, 2016, 36(10): 2945–2953. |