 2017, Vol. 37
2017, Vol. 37文章信息
- 王博涵, 吴丹, 张吉, 殷旭旺, 赵长森, 窦同文
- WANG Bohan, WU Dan, ZHANG Ji, YIN Xuwang, ZHAO Changsen, DOU Tongwen.
- 济南河流大型底栖动物摄食功能群多样性及时空动态
- Diversity and temporal-spatial dynamics of macroinvertebrate functional feeding groups in the rivers of the Jinan Region
- 生态学报. 2017, 37(21): 7128-7139
- Acta Ecologica Sinica. 2017, 37(21): 7128-7139
- http://dx.doi.org/10.5846/stxb201608111643
-
文章历史
- 收稿日期: 2016-08-11
- 网络出版日期: 2017-07-11





2. 北京师范大学, 地理学与遥感科学学院, 北京 100875;
3. 济南市水文局, 济南 250014
2. College of Geography and Remote Sensing Science, Beijing Normal University, School of Geography and Remote Sensing Science, Beijing 100875, China;
3. Ji'nan Hydrographic Office, Ji'nan 250014, China
大型底栖动物在河流生态系统中有着重要的生态学作用, 其通常以有机颗粒、凋落物和藻类等为食[1], 并为水中鱼类提供食物的来源, 其对河流生态系统的物质循环和能量流动和信息传递起着重要的作用[2]。大型底栖动物处于底栖碎屑食物链的关键环节[3], 通过摄取底泥中的有机物质作为营养物质的来源, 从而将水体的底质翻均, 进一步加速水体的自净过程[4]。大型底栖动物作为河流生态系统健康的指示类群, 在生物评价中有着较多优点, 如栖息环境稳定、生命周期长、易于采集、分布广泛等等[5], 因此, 大型底栖动物群落结构及多样性等指标在很大的程度上反映了整个河流生态系统的健康状况[6-7], 并对河流保护、管理和利用具有指导的作用[8]。
在河流生态系统的研究中, 依据食性可将大型底栖动物划分为:刮食者(Scrapers)、撕食者(Shredders)、收集者(Gather-collectors)、滤食者(Filter-collectors)和捕食者(Predators), 即为功能摄食类群(functional feeding groups, FFGs)[9-11]。大型底栖动物功能摄食类群可以反映出大型底栖动物的群落结构特征和生境适应性特征[9-11], 其有别于传统的大型底栖动物形态分类方法, 能更精确地表现大型底栖动物群落与不同水体环境压力的相关性。大型底栖动物摄食功能类群的划分及多样性能够反映出人类活动的干扰强度, 并准确的反应河流生态系统的受损状况。目前, 国际上关于大型底栖动物功能摄食类群研究主要分为两类:一是研究大型底栖动物功能摄食类群对人类活动的响应;二是建立功能类群的评价体系及应用[9-11]。在我国专门针对淡水大型底栖动物功能摄食类群的报道不多[12]。本研究于春季(5月)、夏季(8月)和秋季(11月)对黄河流域及淮河流域流经的济南地区的河流进行了调查研究, 对大型底栖动物功能摄食类群时空分布进行研究, 分析大型底栖动物群落与水环境因子之间的相关性, 以期分析出对大型底栖动物群落结构起主要影响的环境因子, 为济南地区生态环境的保护和管理提供基础数据。
1 材料和方法 1.1 地区概况及点位设置济南位于山东省中西部, 总面积为8177.21 km2, 南依泰山, 北跨黄河。黄河流域与淮河流域贯穿汇入其中, 水生态系统类型复杂。近年来, 由于人类活动导致济南地区植被覆盖率降低、地下水资源持续枯竭、城区污染加剧及生物多样性降低等问题严重阻碍了济南市的可持续发展。为了能够更好地区分济南地区大型底栖动物群落结构的空间差异, 根据济南地区地理特征、水质状况及污染分布情况, 在济南市区及各县城的代表性区域共设定24个监测点位, 其中黄河流域12个采样点位, 其流域两岸植被较少, 底质为泥沙底, 水流相对缓慢, 部分点位有泥沙淤积;淮河流域12个采样点位, 流域内多为山区、丘陵区, 高程一般在100 m左右, 两岸植被茂盛, 支流长而多, 水流速度慢, 底质为砂石底(见图 1)。

|
| 图 1 济南地区采样点分布 Fig. 1 The sampling sites distribution in Jinan Region |
在所选定点位的100 m范围之内, 使用索伯网(Surber net, 网口尺寸为30 cm× 30 cm, 网孔径为500 μm)随机采集2个平行样本, 现场用75%的酒精固定。样品带回实验室后, 将肉眼可见的底栖动物与石块等杂质分离, 并置于100 mL样品瓶中, 用75%的酒精固定保存, 在显微镜(OLYMPUS-CX21)或解剖镜(OLYMPUS-SZX7)下进行分类和计数, 依据相关的文献资料, 样品尽量鉴定到最低分类单元[13-16]。
1.3 水体理化性质的测定水体理化指标, 采取现场测定和实验室测定相结合的方式。现场利用手持式YSI多参数水质监测分析仪(YSI ProPlus)现场测定水温(Temp)、pH、溶解氧(DO), 各点位采集2 L水样, 低温保存, 48 h内送回实验室根据标准方法测定电导率(Cond)、钙离子(Ca2+)、氯离子(Cl-)、硬度(TD)、碱度(ALK)、总磷(TP)、总氮(TN)和高锰酸钾指数(CODMn)[17-19]。
1.4 大型底栖动物功能摄食类群划分及生态系统属性划分根据大型底栖动物的食性类型, 参考Cummins和Bode等[9-11]大型底栖动物功能摄食类群的分类方法, 将济南地区大型底栖动物划分为以下5类(表 1)。
| 功能摄食类 Functional feeding group |
分类标准 Group standard |
代表类群 Representative group |
| 刮食者Scrapers | 主要以各种营固着生活的生物类群为食, 如着生藻类等 | 萝卜螺属、环棱螺属 |
| 撕食者Shredders | 主要以各种凋落物和粗有机质颗粒为食(粒径>1 mm) | 长臂虾属、白虾属 |
| 收集者Gather-collectors | 主要取食河底的各种有机颗粒物 | 摇蚊属、直突摇蚊属 |
| 滤食者Filter-collectors | 以水流中的细有机颗粒物为食(0.45 mm<粒径<1 mm) | 水丝蚓属 |
| 捕食者Predators | 以捕食其他水生动物为食 | 蜉蝣属、蜓属 |
根据Yoshimura等[20]的研究分析, 本研究从物质循环、物质循环纵向输送能力、沿岸物质输入和粗有机颗粒输入量/细有机颗粒输入量等方面进行分析, 应用基于大型底栖动物功能摄食类群的参数对河流生态系统进行评价(表 2)。
| 基于功能摄食类群的参数 Metrics based on functional feeding groups |
生态系统属性 Ecosystem attributes |
| 物质循环Material cycling | |
| F1刮食者密度, G2 Density of scrapers, G2 | 初级生产者 |
| F2生物量Biomass | 次级生产者 |
| F3刮食者密度与滤食者、收集者之和的比率, G2/(G3+G4) Ratio of scrapers to filterers and gathering-collectors, G2/(G3+G4) |
自养作用/异养作用 |
| F4撕食者密度和收集者密度之和, G1+G4 Density of shredders and gathering-collectors, G1+G4 |
分解能力 |
| 物质纵向输入能力Longitudinal transport | |
| F5滤食者密度, G3 Density of filterers, G3 | 纵向输入能力 |
| F6滤食者与撕食者和收集者之和的比率, G3/(G1+G4) Ratio of filterers to shredders and gathering-collectors, G3/(G1+G4) |
相对纵向输送能力 |
| 沿岸物质的输入Lateral input | |
| F7撕食者密度, G1 Density of shredders, G1 | 沿岸物质的输入 |
| F8撕食者密度与总密度之比, G1/总密度 Ratio of shredders to total abundance, G1/total density |
相对物质输入量 |
| 其他Others | |
| F9撕食者与滤食者和收集者之和的比率, G1/(G3+G4) Ratio of shredders to filterers and gathering-collectors, G1/(G3+G4) |
粗有机颗粒输入量/细有机颗粒输入量 |
| G1:撕食者shredders;G2:刮食者scrapers;G3:滤食者filterers;G4:收集者gathering-collectors;G5:捕食者predators;CPOM:粗有机颗粒输入量coarse particulate organicmatter;FPOM细有机颗粒输入量fine particulate organicmatter | |
计算Shannon-Wiener指数(H′)、Pielou均匀度(J)研究大型底栖动物群落多样性。
Shannon-Wiener多样性指数(H′):
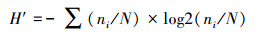
|
Pieiou均匀度指数(J):
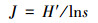
|
优势种确定:根据每个种的优势度值(Y)确定大型底栖动物的优势种:
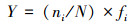
|
式中, s为种类数;N为大型底栖动物的总密度, ni为第i种的密度, fi为第i种出现的频率。Y>0.02的种类则确定为调查的优势种。
采用箱体图和折线图分析大型底栖动物群落及功能摄食类群在3个季节的分布特征。柱状图分析和比较了各季节生态系统属性。采用独立样本t检验分析3个季节环境因子的相关性以及与大型底栖动物之间的相关性。各采样点位筛选出的水环境参数和大型底栖动物群落数据在Canoco 4.5软件上进行典范对应分析(Canonical correspondence analysis, CCA)。
应用大型底栖动物密度进行CCA分析时, 先进行除趋势对应分析(Detrended correspondence analysis, DCA), 得出物种的单峰响应值(梯度SD), SD>2, 即可进行CCA, 否则采用冗余度分析(Redundancy analysis, RDA), 并对数据进行999次蒙特卡洛置换检验(Monte Carlo permutation test), 以判定显著影响大型底栖动物群落空间分布特征的环境因子。在进行CCA分析之前, 除pH以外, 所有水体环境数据和大型底栖动物密度数据均进行数据对数转换[log (x+1)][21]。
用SPSS 16.0 Pearson相关性分析对数据进行单因素方差分析, DCA和CCA在Canoco 4.5上进行, Shannon-Wiener指数和均匀度指数计算在Biodiversity pro上进行, 柱状图、箱体图和折线图分析OriginPro 7.5上进行。
2 结果 2.1 大型底栖动物群落结构特征及其功能摄食类群划分 2.1.1 大型底栖动物物种组成及其群落结构济南地区春季、夏季和秋季共鉴定出大型底栖动物57种, 其中春季45种、夏季35种、秋季33种, 3个季节均以节肢动物门种类最多, 其次是软体动物门, 第三是环节动物门。常见种为东方蜉(Ephemera orientalis)、溪流摇蚊、苍白摇蚊、大耳萝卜螺(Radix auricularia)、喜盐摇蚊和豆螺。春季优势种为霍甫水丝蚓(Y=0.08)和豆螺(Y=0.07);夏季优势种为豆螺(Y=0.05);秋季优势种为喜盐摇蚊(Y=0.07)和豆螺(Y=0.02)(表 3)。春季:大型底栖动物物种密度平均值为2.49×103个体/m2, 生物量平均值为495.59 g/m2, Shannon-Wiener指数平均值为1.37, 均匀度平均值0.55;夏季:大型底栖动物物种密度平均值为0.56×103个体/m2, 生物量平均值为137.26 g/m2, Shannon-Wiener指数平均值为1.33;均匀度平均值0.67;秋季:大型底栖动物物种密度平均值为1.03×103个体/m2, 生物量平均值为109.45 g/m2, Shannon-Wiener指数平均值为1.17, 均匀度平均值0.59。
| 季节Season | 优势种Dominant species | 优势度Y |
| 春季Spring | 霍甫水丝蚓Limnodrilus hoffmeisteri | 0.08 |
| 豆螺Bithynia fuchsiana | 0.07 | |
| 夏季Summer | 豆螺Bithynia fuchsiana | 0.05 |
| 溪流摇蚊Chironomus riparius | 0.03 | |
| 秋季Autumn | 喜盐摇蚊Chironomus salinarius | 0.07 |
| 豆螺Bithynia fuchsiana | 0.02 |
就全地区而言, 3个季节大型底栖动物具有较明显的空间异质性。春季与夏季大型底栖动物生物量和Shannon-Wiener指数差异显著(P=0.03, P=0.02), 其他季节大型底栖动物密度、生物量、Shannon-Wiener指数和均匀度指数均无差异(P春季与夏季=0.21, P春季与秋季=0.34, P夏季与秋季=0.42, P春季与秋季=0.68, P夏季与秋季=0.91, P春季与秋季=0.51, P夏季与秋季=0.65, P春季与夏季=0.32, P春季与秋季=0.42, P夏季与秋季=0.26)。对各流域而言, 黄河流域和淮河流域大型底栖动物密度和生物量差异均不显著(P春季=0.11, P夏季=0.42, P秋季=0.38, P春季=0.51, P夏季=0.38, P秋季=0.75);夏季黄河流域和淮河流域Shannon-Wiener指数和均匀度指数差异显著(P=0.04, P=0.04), 春季和秋季Shannon-Wiener指数和均匀度指数不显著(P=0.29, P=0.67, P=0.64, P=0.08)。

|
| 图 2 济南地区3个季节大型底栖动物群落结构特征的比较分析4个指标的箱体图(D-物种密度;B-生物量;H′-Shannon-Wiener指数;J -均匀度指数) Fig. 2 Comparison of community structure of macroinvertebrates in spring、summer and autumn (D-Density; B-Biomass; H′-Shannon-Wiener index; J-Pielou index) |
济南地区大型底栖动物功能摄食类群共为5种, 3个季节均为5种功能摄食类群。图 3为济南地区黄河流域与淮河流域底栖动物功能摄食类群各类种类和密度[lg(x+1)转换]比较。就功能摄食类群各类的种类而言, 春季两流域收集者种类均为最高, 分别为13和12种, 夏季两流域刮食者种类均为最多, 分别为11种和10种, 秋季黄河流域刮食者和收集者种类较多, 均为8种, 淮河流域刮食者种类最多为10种。从物种密度来看:黄河流域3个季节中均以收集者(3.92, 3.66, 3.80)密度最高。淮河流域春季滤食者密度最高(4.11), 夏季(3.00)和秋季(3.87)收集者密度最高。功能摄食类群的种类和密度在不同流域, 不同季节均无显著相关性(P>0.05)。

|
| 图 3 济南地区不同季节底栖动物功能摄食类群种类和密度比较 Fig. 3 The compare of number and density of macroinvertebrates functional feeding group in Jinan Region |
基于功能摄食类群的各项参数进行两个流域空间差异分析表明:F4、F7和F9数值在黄河流域明显高于淮河流域;F1、F2、F5和F6数值在淮河流域明显高于黄河流域, F3和F8在两流域数值相差不大。
基于功能摄食类群的各项参数对各个季节进行实践差异分析表明:F1、F5和F6数值表现为春季明显高于夏季与秋季, F2数值表现为秋季明显高于春季与夏季, F3数值表现为各季节相差不大, F4数值表现为春季与秋季明显高于夏季, F7、F8和F9数值表现为夏季和秋季明显高于春季。
(1) 在物质循环方面:底栖动物初级生产力F1在淮河流域明显高于黄河流域, 春季大于其他季节;次级生产力F2在淮河流域明显高于黄河流域, 秋季明显大于其他季节;自养/异养F3在两个流域间无显著差异性, 各季节见无显著差异性;分解能力F4在黄河流域高于淮河流域, 春季明显高于其他季节。(2)物质纵向输送能力:纵向输送能力F5淮河流域高于淮河流域, 春季明显高于其他季节。相对输送能力F6在淮河流域明显高于黄河流域, 春季明显高于其他季节。(3)沿岸物质输入:沿岸物质输入量F7两个流域间无显著差异性, 夏季与秋季明显高于春季;相对输入量F8淮河流域明显高于黄河流域, 春季明显高于其他季节;粗有机颗粒输入量/细有机颗粒输入量F9两个流域之间无显著差异性(P=0.44), 夏季与秋季明显高于春季(图 4)。

|
| 图 4 黄河流域与淮河流域生态系统特征值的季节变化 Fig. 4 The ecosystem attributes of Huang River Basin and Huai River Basin in spring、summer and autumn |
济南地区水环境因子在不同季节和不同流域间相关性分析如表 4所示, 济南地区夏季水体温度最高, 秋季水体温度最低, 水体全年呈弱碱性, pH 7.74—8.21。3个季节中淮河流域电导率、钙离子、氯离子、总硬度、总碱度、总氮、高锰酸钾指数和总磷数值均高于黄河流域, 黄河流域溶解氧含量高于淮河流域。3个季节中黄河流域与淮河流域11个环境指标均无显著相关性(P>0.05)。
| 环境因子 Environmental factors |
春季Spring | 夏季Summer | 秋季Autumn | ||||||||
| 黄河流域 Huang River Basin |
淮河流域 Huai River Basin |
P | 黄河流域 Huang River Basin |
淮河流域 Huai River Basin |
P | 黄河流 Huang River Basin |
淮河流域 Huai River Basin |
P | |||
| 温度Temp(℃) | 19.38±2.06 | 22.30±3.71 | 0.09 | 27.90±1.70 | 26.45±3.31 | 0.70 | 14.37±1.37 | 14.31±1.60 | 0.26 | ||
| pH | 8.21±0.30 | 7.81±0.32 | 0.59 | 8.20±0.30 | 7.74±0.35 | 0.59 | 8.17±0.30 | 7.96±0.39 | 1.00 | ||
| 电导率Cond/(ms/m) | 654.25±497.27 | 1222.67±925.47 | 0.23 | 550.07±395.45 | 1151.08±744.14 | 0.19 | 563.52±436.54 | 1194.39±746.45 | 0.32 | ||
| ρ(Ca2+)/(mg/L) | 68.54±44.40 | 111.89±47.88 | 0.06 | 60.23±38.05 | 102.73±34.61 | 0.73 | 61.27±43.62 | 111.80±21.86 | 0.93 | ||
| ρ(Cl-)/(mg/L) | 55.64±84.16 | 124.06±149.62 | 0.35 | 55.19±50.83 | 124.32±144.91 | 0.25 | 53.02±58.60 | 130.57±150.36 | 0.41 | ||
| ρ(ALK)/(mg/L) | 145.16±58.81 | 221.34±50.29 | 0.37 | 120.43±47.61 | 187.67±43.86 | 0.41 | 123.01±54.20 | 205.64±51.75 | 0.62 | ||
| ρ(TD)/(mg/L) | 255.15±124.30 | 430.89±203.23 | 0.17 | 222.88±114.44 | 389.52±136.42 | 0.37 | 226.10±126.72 | 400.64±128.50 | 0.37 | ||
| ρ(DO)/(mg/L) | 8.45±1.96 | 7.90±2.69 | 0.13 | 8.21±1.11 | 6.79±2.50 | 0.40 | 8.17±1.28 | 7.54±1.85 | 0.70 | ||
| ρ(TN)/(mg/L) | 3.85±2.57 | 6.61±6.25 | 0.16 | 3.63±2.44 | 6.34±4.64 | 0.24 | 3.68±2.57 | 6.54±4.90 | 0.52 | ||
| ρ(CODMn)/(mg/L) | 3.89±2.70 | 5.01±3.03 | 0.79 | 3.75±1.11 | 5.36±3.69 | 0.26 | 3.59±1.23 | 4.61±2.37 | 0.37 | ||
| ρ(TP)/(mg/L) | 0.37±1.03 | 0.39±0.42 | 0.42 | 0.14±0.19 | 0.31±0.40 | 0.56 | 0.17±0.20 | 0.49±1.01 | 0.65 | ||
选择各类大型底栖动物功能摄食类群密度和环境因子做CCA, 结果显示(图 5):3个季节大型底栖动物功能摄食类群与环境因子对应关系有所差异, 春季:ρ(TP)(P=0.04)是主要影响黄河流域大型底栖动物功能摄食类群的环境因子, 并对第二轴影响最大, 其主要影响该流域下游区域。ρ(TN)(P=0.03)和ρ(TP)(P=0.04)是主要影响淮河流域大型底栖动物功能摄食类群的环境因子;夏季:pH(P=0.03)是对黄河流域大型底栖动物功能摄食类群起主要影响的环境因子, ρ(DO)(P=0.01)是对淮河流域大型底栖动物功能摄食类群起主要影响的环境因子;秋季:ρ(DO)(P=0.03)是对黄河流域大型底栖动物功能摄食类群起主要影响的环境因子, 对淮河流域大型底栖动物功能摄食类群起主要影响的环境因子是pH(P=0.04)。

|
| 图 5 大型底栖动物功能摄食类群与环境因子的典范对应性分析(CCA) Fig. 5 Canonical correspondence analysis of species-environmental relationship in different seasons G1:刮食者;G2:滤食者;G3:撕食者;G4:收集者;G5:捕食者;电导率:Cond; 钙离子:Ca2+;总碱度:ALK;总硬度:TD;溶解氧:DO;总氮:TN;氨氮:NH3-N; 高锰酸钾指数:CODMn;总磷:TP;氯离子:Cl- |
济南地区此次调查共鉴定出大型底栖动物57种, 3个季节均以节肢动物为主, 春季种类最多为45种。鉴定出的大型底栖动物物种以双翅目摇蚊幼虫为主, 其大多数习惯生活在污水或淤泥或有机质丰富的水体中, 这说明济南部分水体受到了一定程度污染。春季优势种是霍甫水丝蚓和豆螺, 夏季的优势种是溪流摇蚊和豆螺, 秋季的优势种是喜盐摇蚊和豆螺, 豆螺在3个季节中均有一定优势性。从优势种季节变化来看, 济南大型底栖动物群落存在一定季节性变化。
3个季节中大型底栖动物主体群落结构均以摇蚊幼虫和粘附者螺类等为主导。春季、夏季和秋季密度最高的点位位于J16(17.33×103个体/m2)、J8(4.02×103个体/m2)和J22(6.54×103个体/m2)。J16位于淮河流域, 以水中双翅目幼虫为主, 尤其是春季为摇蚊科幼虫数量处于繁殖旺盛时期, 使得生物总密度呈现大幅升高。J8位于黄河流域, 水流速缓慢, 种类多为粘附者, 依靠有力的附肢、吸盘和粘附性虫巢得以生存, 使得生物总密度呈现大幅升高[22]。J22位于淮河流域, 底质大多以基石和砂石为主, 由于水流速缓慢, 致使附着性大型底栖动物及双翅目种类生活在基石与泥沙中, 使得生物总密度呈现大幅升高。
Shannon-Wiener指数常用来监测大型底栖动物群落结构的变化, 是一个评价水域有机污染程度的有效工具[23-24]。国内外许多学者都应用大型底栖动物多样性指数进行了水质评价。香农维纳指数在区间0—1为重度污染、1—2为中度污染、2—2.5为轻度污染, 2.5—3.5为清洁, 大于3.5为最清洁[25]。济南地区3个季节香农维纳指数平均值均在1—2之间, 从而判断济南地区水体为中度污染。污染原因可能是由于济南地区人口数量较多, 人类活动对水体及环境影响较大, 生活污水排放及工业污染严重影响了济南地区水质。
济南地区共划分出5类功能摄食类群, 其中收集者密度明显高于其他类群。收集者以有机碎屑为食, 流速低的区域有机碎屑较易沉积, 因此收集者偏爱流速较低的生境[26], 3个季节中黄河流域收集者密度均要高于淮河流域密度, 主要由于黄河流域下游点位底质以淤泥为主, 水面宽度较大, 有机营养物质较多, 流速较慢, 满足了收集者摄食要求, 为收集者提供了丰富的食物来源, 而淮河流域水流速相对较快, 食物较少, 不适合收集者生活;撕食者主要以沿岸植被[27-28]和堆积在河道底的树叶为食[29-30], 水流加快会导致撕食者的捕食能力下降。春季和秋季撕食者密度要高于夏季, 黄河流域撕食者密度高于淮河流域, 这是由于春季和秋季黄河流域河岸带农田、植被覆盖率较高, 枯枝树叶堆积较多, 为撕食者提供了大量食物来源;刮食者密度表现为秋季高于春季和夏季, 主要是因为树木的落叶期主要集中与秋季, 为刮食者提供了丰富的食物来源。底质类型对大型底栖动物食物的获取也有较大影响, 卵石具有较大的表面积, 可以滞留较多的粗颗粒有机物, 附生更多的藻类和水生植物, 为刮食者提供丰富的食物来源[26-27]。淮河流域刮食者密度明显高于黄河流域, 主要由于淮河流域水体底质中卵石数量较多, 占具面积较大, 而黄河流域底质为泥沙底质, 致使刮食者密度相对较低。卵石能够支持以刮食者为食的更高营养级物种的生存繁殖, 形成复杂的食物链, 进而提高大型底栖动物群落结构多样性[31];滤食者以细有机颗粒物为食, 一般都有一个最适滤食流速[12];滤食者密度主要受到水流速及食物影响, 淮河流域密度较高, 主要由于淮河流域食物及水流速等环境因子更适合滤食者摄食, 夏季滤食者密度高于春季及秋季, 是由于夏季能提供更多滤食者生存的食物及适合其摄食的水文条件;流速较大的区域适合捕食者生活, 大部分捕食者均适合缓流生境;捕食者密度在两个流域并没有明显差异性, 黄河流域捕食者密度高于淮河流域, 主要是由于黄河流域水流速较慢, 适合捕食者捕食和生活。春季捕食者密度高于其他季节, 是因为春季其他类群密度较大, 为捕食者提供了丰富的食物。
3.2 大型底栖动物功能摄食类群与环境因子的关系评价水生生物对某一环境因子的适宜范围在河流生态修复中有着重要作用[27], 这是研究水生生物与环境因子关系的主要方式。许多研究表明, 不同季节影响大型底栖动物分布的环境因子存在显著差异[32-33]。通过CCA对济南地区3个季节大型底栖动物功能摄食类群和水环境因子的相关性分析, 结果显示:3个季节中大型底栖动物功能摄食类群与水生态因子对应关系有所差异。总磷和总氮是衡量水体受营养物质污染程度的重要指标[34], 春季:黄河流域内水体含沙量极大, 水体浊度较高, 透明度低, 总磷含量较高, 总磷对滤食者和刮食者起主要影响作用, 与滤食者呈正相关性, 与刮食者呈负相关性, 磷含量的增加影响了滤食者和刮食者的生长繁殖。淮河流域总氮对刮食者和捕食者起到主要影响作用, 与刮食者和捕食者呈负相关性, 总氮含量较高, 导致刮食者和捕食者的生活环境发生改变[35];大型底栖动物喜欢生活在酸碱度适宜的环境下, 夏季:黄河流域水体偏碱性, pH对是主要影响撕食者的环境因子, 与撕食者呈正相关性, 夏季pH最高, 为8.21, 撕食者的数量在夏季达到高峰。水中溶解氧含量的多少直接影响大型底栖动物的生活与繁殖, 溶解氧含量下降可能会引起水中大型底栖动物的死亡[36]。淮河流域溶解氧对刮食者与撕食者起影响作用, 溶解氧与刮食者与撕食者呈负相关性, 淮河流域溶氧含量较低, 限制了撕食者的生存, 而耐污能力较强的刮食者则大量繁殖, 成为绝对优势种;秋季:黄河流域溶解氧对滤食者起主要影响作用, 与滤食者呈负相关性, 淮河流域pH对捕食者起主要影响作用, 与捕食者呈正相关性。通过分析表明, 不同时空尺度对大型底栖动物功能类群群落起主要影响的环境因子并不相同, 各类环境因子基本上可归纳为3类:1)物理因素:包括水温、流速和电导率等;2)富营养化因素:包括氮、磷等元素以及各类离子含量、有机物和营养盐等;3)底质类型, 如泥沙、卵石等。
| 目 | 种 |
| 蜉蝣目 | 黑翅四节蜉Baetis vaillanti |
| 东方蜉Ephemera orientalis | |
| 毛翅目 | 虎皮纹石蛾Hydropsyche sp. |
| 双翅目 | 费塔无突摇蚊Ablabesmyia phatta |
| 刺铗长足摇蚊Tanypus punctipennis | |
| 浪突摇蚊Zalutschia sp | |
| 石栖直突摇蚊Orthocladius saxicola | |
| 塔马拟环足摇蚊paracricotopus tamabrevis | |
| 三带环足摇蚊Cricotopus trifasciatus | |
| 特式直突摇蚊Orthocladius thienemanni | |
| 三轮环足摇蚊C. over | |
| 长跗摇蚊Tanytarsus gracilistylus | |
| 若西摇蚊Chironomus yoshimatusi | |
| 喙隐摇蚊Crytochironomus rostratus | |
| 溪流摇蚊C. riparius | |
| 喜盐摇蚊C. salinarius | |
| 猛摇蚊C. acerbiphilus | |
| 软铗小摇蚊Microchironomus tener | |
| 等叶裸须摇蚊Pparadoxus | |
| 苍白摇蚊C. pallidivittatus | |
| 分齿恩非摇蚊Einfeldia dissidens | |
| 德永雕翅摇蚊Glyptotendipes tokunagai | |
| 小云多足摇蚊Polypedilum nubeculosum | |
| 云集多足摇蚊P. nubifer | |
| 梯形多足摇蚊P. scalaenum | |
| 俊才齿斑摇蚊Stictochironomus juncaii | |
| 双叉巨吻沼蚊Antocha bifida Alexander | |
| 大蚊Tipula sp. | |
| 广翅目 | 日本星齿蛉Parachauliodes japonicas |
| 蜻蜓目 | 箭蜓Gomphs sp. |
| 亚洲瘦蟌Ischnura asiatica | |
| 蜓Aeschna | |
| 江鸡稚虫M.clio | |
| 十足目 | 克氏原鳌虾Procambarus clarkii |
| 秀丽白虾Palaemon modestus | |
| 中华小长臂虾Palaemonetes sinensis | |
| 基眼目 | 直缘萝卜螺Radix clessini |
| 大耳萝卜螺R.auricularia | |
| 狭耳萝卜螺R.tagotis | |
| 卵萝卜螺R. ovata | |
| 大脐圆扁螺physa acuta cf. | |
| 中国圆田螺Cipangopaludina chinensis | |
| 铜锈环棱螺Bellamya aeruginosa | |
| 梨形环棱螺B.quadrata | |
| 方形环棱螺B.quadrata | |
| 短沟蜷Semisulcospira | |
| 拟沼螺Assimineidae.sp | |
| 豆螺B.fuchsiana | |
| 真瓣鳃目 | 钳形无齿蚌A.arcaeformis |
| 圆顶珠蚌Eulamellibranchia | |
| 河蚬Corbicula fluminea | |
| 颤蚓目 | 霍甫水丝蚓Limnodrilus hoffmeisteri |
| 苏式尾鳃蚓Branchiura soJerbyi | |
| 克拉伯水丝蚓L.claparedianus | |
| 颚蛭目 | 宽体金线蛭Jhitmania pigra |
| 无吻蛭目 | 葦氏白勃石蛭Barbronia Jeberi |
| 扁蛭Barbronia flat |
| [1] | Rosi-Marshall E J, Wallace J B. Invertebrate food webs along a stream resource gradient. Freshwater Biology, 2002, 47(1): 129–141. DOI:10.1046/j.1365-2427.2002.00786.x |
| [2] | Heino J, Muotka H, Mykrä R, Paavola R, Hämäläinen H, Koskenniemi E. Defining macroinvertebrate assemblage types of headwater streams: implications for bioassessment and conservation. Ecological Applications, 2003, 13(3): 842–852. DOI:10.1890/1051-0761(2003)013[0842:DMATOH]2.0.CO;2 |
| [3] | Cummins K W. Structure and function of stream ecosystems. BioScience, 1974, 24(11): 631–641. DOI:10.2307/1296676 |
| [4] | 段学花, 王兆印, 徐梦珍. 底栖动物与河流生态评价. 北京: 清华大学出版社, 2010: 47–91. |
| [5] | KoskenniemiE. The role of chironomids(Diptera)in the profundalmacrozoobenthos in Finnish reservoirs. Netherlands Journal of Aquatic Ecology, 1992, 26(2/4): 503–508. |
| [6] | Barbour M T, GerritsenJ, Snyder B D, Stribling J B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates, and Fish. 2nd ed. Washington, DC: US EPA, Office of Water, 1999. |
| [7] | Wright J F. Development and use of a system for predicting the macroinvertebrate fauna in flowing waters. Australian Journal of Ecology, 1995, 20(1): 181–197. DOI:10.1111/aec.1995.20.issue-1 |
| [8] | Hughes D L, Gore J A, Brossett M P, Olson J R. Rapid Bioassessment of Stream Health. New York: CRC Press, 2010. |
| [9] | Zhang Y, Zhao R, Kong W J, Geng S W, Bentsen C N, Qu X D. Relationships between macroinvertebrate communities and land use types within different riparian widths in three headwater streams of Taizi River, China. Journal of Freshwater Ecology, 2013, 28(3): 307–328. DOI:10.1080/02705060.2013.779941 |
| [10] | Barbour M T, Gerritsen J, SnyderB D, Stribling J B. RapidBioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish.2nded. Washington, DC: U.S. Environmental Protection Agency, Office of Water, 1999. |
| [11] | Bode R W, Novak M A, Abele L E, Heitzman DL, Smith A J. Quality Assurance Work Plan for Biological Stream Monitoring in New York state. Albany, NY:NYS Department of Environmental Conservation, 2002. |
| [12] | 蒋万祥, 蔡庆华, 唐涛, 渠晓东. 香溪河水系大型底栖动物功能摄食类群生态学. 生态学报, 2009, 29(10): 5207–5218. DOI:10.3321/j.issn:1000-0933.2009.10.005 |
| [13] | Thorp J H, Covich A P. Ecology and Classification of North American Freshwater Invertebrates.2nd ed. New York: Academic Press, 2001. |
| [14] | 王桢瑞. 中国动物志-软体动物门-双壳纲. 北京: 科学出版社, 2002. |
| [15] | 王艳杰. 辽河流域大型底栖动物群落特征与生境指标关系[D]. 沈阳: 辽宁大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10140-1013103651.htm |
| [16] | Merritt R W, Cummins K W, Berg M B. An Introduction to the Aquatic Insects of North America.4th ed. Dubuque: KendallHunt Publishing, 2008. |
| [17] | 殷旭旺, 徐宗学, 高欣, 白海锋, 武玮, 宋进喜. 渭河流域大型底栖动物群落结构及其与环境因子的关系. 应用生态学报, 2013, 24(1): 218–226. |
| [18] | 殷旭旺, 徐宗学, 鄢娜, 武玮, 宋进喜. 渭河流域河流着生藻类的群落结构与生物完整性研究. 环境科学学报, 2013, 33(2): 518–527. |
| [19] | 国家环境保护总局. 水和废水监测分析方法(第四版). 北京: 中国环境科学出版社, 2002. |
| [20] | Yoshinura C, Tockner K, Omura T, Moog O. Species diversity and functional assessment of macroinvertebrate communities in Austrianrivers. Limnology, 2006, 7(2): 63–74. DOI:10.1007/s10201-006-0170-4 |
| [21] | 殷旭旺, 渠晓东, 李庆南, 刘颖, 张远, 孟伟. 基于着生藻类的太子河流域水生态系统健康评价. 生态学报, 2012, 32(6): 1677–1691. |
| [22] | 殷旭旺, 李庆南, 朱美桦, 宋佳, 武玮, 徐宗学. 渭河丰、枯水期大型底栖动物群落特征及综合健康评价. 生态学报, 2015, 35(14): 4784–4796. |
| [23] | 郝卫民, 王士达, 王德铭. 洪湖底栖动物群落结构及其对水质的初步评价. 水生生物学报, 1995, 19(2): 124–134. |
| [24] | Van Dolah R F, Hyland J L, Holland A F, Rosen J S, Snoots T R. Abenthic index of biological integrity for assessing habitat quality inestuaries of the southeastern USA. Marine Environmental Research, 1999, 48(4/5): 269–283. |
| [25] | 吴召仕, 蔡永久, 陈宇炜, 邵晓阳, 高俊峰. 太湖流域主要河流大型底栖动物群落结构及水质生物学评价. 湖泊科学, 2011, 23(5): 686–694. DOI:10.18307/2011.0504 |
| [26] | Mérigoux, OlédecS. Hydraulic requirements of stream communities: a case study on invertebrates. Freshwater Biology, 2004, 49(5): 600–613. DOI:10.1111/fwb.2004.49.issue-5 |
| [27] | MollesM C Jr. Trichopteran communities of streams associated with aspen and conifer forests: long-term structural change. Ecology, 1982, 63(1): 1–6. DOI:10.2307/1937023 |
| [28] | Cummins K W, WilzbachM A, Gates D M, Perry J B, Taliaferro WB. Shredders and riparian vegetation. BioScience, 1989, 39(1): 24–30. DOI:10.2307/1310804 |
| [29] | Dobson M. Microhabitat as a determinant of diversity: stream invertebrates colonizing leaf packs. Freshwater Biology, 1994, 32(3): 565–572. DOI:10.1111/fwb.1994.32.issue-3 |
| [30] | HaapalaA, MuotkaT, LaasonenP. Distribution of benthic macroinvertebrates and leaf litter in relation to streambed retentivity: implications for headwater stream restoration. Boreal Environment Research, 2003, 8(1): 19–30. |
| [31] | 王强, 袁兴中, 刘红. 西南山地源头溪流附石性水生昆虫群落特征及多样性——以重庆鱼肚河为例. 水生生物学报, 2011, 35(5): 887–892. |
| [32] | 赵进勇, 董哲仁, 孙东亚. 河流生物栖息地评估研究进展. 科技导报, 2008, 26(17): 82–88. DOI:10.3321/j.issn:1000-7857.2008.17.014 |
| [33] | 蒋万祥, 贾兴焕, 周淑婵, 李凤清, 唐涛, 蔡庆华. 香溪河大型底栖动物群落结构季节动态. 应用生态学报, 2009, 20(4): 923–928. |
| [34] | 寿鹿, 曾江宁, 廖一波, 赵永强, 江志兵, 陈全震, 高爱根, 杨家新. 瓯江口海域大型底栖动物分布及其与环境的关系. 应用生态学报, 2009, 20(8): 1958–1964. |
| [35] | 吴东浩, 于海燕, 吴海燕, 周斌, 王备新. 基于大型底栖无脊椎动物确定河流营养盐浓度阈值——以西苕溪上游流域为例. 应用生态学报, 2010, 21(2): 483–488. |
| [36] | 王银东, 熊邦喜, 陈才保, 胡宏胜. 环境因子对底栖动物生命活动的影响. 浙江海洋学院学报:自然科学版, 2005, 24(3): 253-257, 280-280. |