 2017, Vol. 37
2017, Vol. 37文章信息
- 郭磊, 韦博良, 胡金涛, 贾宏汝, 叶永忠, 袁志良
- GUO Lei, WEI Boliang, HU Jintao, JIA Hongru, YE Yongzhong, YUAN Zhiliang.
- 基于两个不同资源轴上苔藓植物生态位分析
- Ecological niche of bryophytes based on two resource axes
- 生态学报. 2017, 37(21): 7266-7276
- Acta Ecologica Sinica. 2017, 37(21): 7266-7276
- http://dx.doi.org/10.5846/stxb201608101638
-
文章历史
- 收稿日期: 2016-08-10
- 网络出版日期: 2017-07-11





2. 河南财政金融学院, 郑州 451464
2. Henan University of Finance & Banking, Zhengzhou 451464, China
生态位理论及应用的研究是国内外学者研究热点, 在探讨群落演替、群落结构以及物种种间关系等科学问题时应用比较广泛[1-3]。在生物群落中, 理论上某一物种能够占据的最大生态位称为基础生态位[4-6]。由于种间竞争等因素的存在, 物种很少能够占据全部基础生态位, 随着群落内物种种内和种间相关性的变化, 物种生态位也会发生变化[7]。
苔藓植物是一类形体较小依靠孢子繁殖的高等植物, 有约23000个种类, 是仅次于被子植物的第二大植物类群, 是生物多样性的重要组成部分[8-9], 也是森林生态系统的重要组成部分, 开展苔藓植物研究有重要的意义。苔藓物种的生长受群落影响很大, 不同群落内苔藓群落结构存在明显差异。郭水良和曹同在对长白山主要生态系统中苔藓植物研究中发现, 在暗针叶林下地面苔藓植物的丰富度最高, 暗针叶林以及红松阔叶混交林与暗针叶林间过渡区树附生苔藓植物的丰富度和多样性最高[10]。Rambo T R和Muir P S在对花旗松林内地面生苔藓植物的研究中发现, 林内的阔叶树种越多, 林间隙越大, 苔藓植物的多样性越高[11]。Király I等[12]、郭水良和曹同[10]研究分别指出苔藓的丰富度还受到灌木层和草本植物的影响。苔藓物种的生长受小生境的影响也很大, 部分苔藓物种具有专一的生境偏好。曹同和郭水良在对长白山地区不同生境类型中苔藓植物研究中发现石生苔藓种类最为丰富, 腐木生和树附生种类次之, 土生种类较少, 沼水生种类最少[13];白学良等在对贺兰山地区的苔藓植物进行调查研究发现石生藓类的种类最多, 土生藓类次之, 木生藓类最少[14]。
关于维管植物尤其是木本植物的生态位研究已经趋于成熟, 但关于苔藓植物的研究还处于初步探索阶段, 与维管植物不同, 苔藓植物不仅对自身生长的小环境反应敏感且大的群落环境对苔藓植物也会产生一定的影响[15], 因此对于苔藓植物种群生态位的研究, 可以使我们更深入地认识苔藓植物种群内和种间的竞争, 对人们深入理解苔藓植物种群在整个森林群落中的地位和作用具有一定的帮助。
对苔藓植物的生态学研究已经做了大量工作, 但是关于苔藓植物生态位研究多是在树附生、土生等单一生境中进行[16-17], 不同生境基质中(树生、石生、土生)苔藓物种生态位特征差异以及森林群落大环境与苔藓自身生长基质小环境对苔藓影响的相关研究还未见报道, 为此本研究通过对小秦岭地区苔藓植物在不同群丛类型下和不同生长基质类型(树生、石生、土生)下的生态位研究, 以期揭示在不同资源轴上苔藓植物生态位特点以及对资源环境的利用能力, 为进一步对该地区苔藓植物资源物种多样性研究和保护提供科学的理论依据。
1 研究区概况小秦岭国家级自然保护区位于豫陕交界河南省灵宝市西部, 地理坐标为34°23′—34°31′N, 110°23′—110°44′E, 保护区东西长31km, 南北宽12km, 总面积15160hm2, 小秦岭山脉北麓主峰老鸦岔垴海拔2413.8m, 属森林生态类型自然保护区, 区内植被良好, 森林覆盖率81.2%[18-20]。本区气候属暖温带大陆性季风型半干旱气候, 四季分明, 年均温11.2—14.2℃, 极端低温-17℃, 极端高温42.7℃, 全年无霜期210d, 年降水量612mm, 且多集中在夏秋季[21]。保护区地层属太古界太华群, 岩石为岩浆岩类, 土壤为薄层酸性岩褐土、薄层酸性岩粗棕壤, pH值为6.2—7.0[22]。
2 研究方法 2.1 样方设置调查与苔藓采集2012年8月在研究区内沿海拔梯度选择代表性群落设置样方, 从海拔1020m开始, 海拔每升高50m设置2个20m×20m的乔木样方, 在每个乔木样方中选择代表性群落设置1个5m×5m的灌木样方, 并在每个乔木样方的4个角设置4个1m×1m的草本样方, 共设置56个样方。对样方内所有胸径≥1cm(DBH≥1cm)乔木, 灌木和草本植物进行调查记录, 共调查维管植物6686株, 隶属143科330属405种。其中乔木以松科(Pinaceae)、漆树科(Anacardiaceae)、杨柳科(Salicaceae)比较多, 灌木以木犀科(Oleaceae)、蔷薇科(Rosaceae)、卫矛科(Celastraceae)较多, 草本以莎草科(Cyperaceae)、禾本科(Gramineae)、菊科(Compositae)较多。
对56个样方中所有苔藓(包括石生苔藓、土生苔藓以及所有木本植物上的树附生苔藓)进行采集, 采集时样方中同一藓丛的每种苔藓均采集装入同一纸袋内, 不同藓丛分别装袋, 如有多种苔藓交错生长则对每种苔藓都进行采集。装袋后记录所采苔藓的样方号及生境基质, 带回实验室, 共采得苔藓标本1378份。根据中国苔藓志(共10卷)、中国苔纲和角苔纲植物属志和中国生物物种名录对所有苔藓植物进行鉴定[23-24], 共鉴定得苔藓植物33科89属240种, 其中苔类9科10属14种, 藓类24科79属226种。苔藓标本保存于河南农业大学标本馆。
为了更好的研究苔藓物种生态位特征, 探寻不同群丛类型与不同生长基质类型对苔藓物种生态位特征的影响, 本研究采用两种资源位的划分方式对苔藓物种生态位进行研究。在群丛类型下, 采用MRT(多元回归树)方法以56个样方海拔、坡向和坡度为自变量, 405种植物在56个样方内的重要值为因变量进行群落分类(具体方法见陈云等人的研究[22]), 将56个样方分为5类不同群丛类型, 作为不同资源位。这5类不同群落组成类型分别为:Ⅰ、领春木-葛萝槭-活血丹+白顶早熟禾(Ass.Euptelea pleiospermum-Acer grosseri-Glechoma longituba+Poa acroleuca)群落, 包括样方14—41, 共28个样方。Ⅱ、华山松+糙皮桦-东陵山柳-宽叶薹草(Ass.Pinus armandii+Betulautilis-Salix phylicifolia-Carex siderosticta)群落, 包括样方44—56, 共13个样方。Ⅲ、蒙古栎-美蔷薇-莎草(Ass.Quercus mongolica-Rosa bella-Cayerus compressus)群落, 包括样方42、43, 共2个样方。Ⅳ、华山松+橿子栎+黄栌-连翘-莎草(Ass.Pinus armandii+Quercus baronii var. Baronii+Cotinus coggygria-Forsythia suspensa-Cayerus compressus)群落, 包括样方1、2、3、5、6、9、12, 共7个样方。Ⅴ、华山松+油松-葛萝槭-求米草(Ass.Pinus armandii+Pinus tabulaeformis-Acer grosseri-Oplismenus undulatifolius)群落, 包括样方4、7、8、10、11、13, 共6个样方。
在生长基质类型下, 依据所采苔藓生长的基质划分为石生、土生和树附生3种, 将这3种生长基质类型作为资源位。每个资源位中包含在调查时生长在该基质上所有苔藓植物。对于生长在不同基质交界处的苔藓物种, 根据假根着生位置的基质类型将其划分到相应资源位中。
2.2 计算方法 2.2.1 重要值重要值是表示某个种在群落中重要程度或者优势度的指标, 其数值大小可作为群落中植物种优势度的一个度量标志, 指示群落中每种植物的相对重要性和植物最适生境[25-27]。
苔藓植物形态较小, 多数苔藓群落分布范围较小, 而使用传统分样点取样时, 多为面积50cm×50cm的小样方, 这种取样方式会降低苔藓植物的物种多样性[13, 28], 且由于苔藓物种孢子传播特殊性, 在有水的地方可能传播较远, 在较大尺度对苔藓物种生态位特征的研究具有重大意义, 因此本研究没有采用传统取样, 而是在20m×20m的样方中进行全部采集调查。鉴于苔藓植物的生物学特点以及本实验调查方法, 对于苔藓植物多度参考贾宏汝[29]在文中的处理方法, 以该种苔藓在一块样方内出现次数为该种苔藓在该样方内的多度, 该种苔藓在该样方内出现的次数与所有样方内出现的次数的比值即为该物种的频度。苔藓植物重要值计算中, 以相对多度和相对频度作为计算重要值参数[30]。
重要值= (相对多度+相对频度) / 2×100%
相对多度=(某种苔藓植物在样方内多度/样方内所有苔藓多度总和)×100%
相对频度=(某种苔藓在所有样方中出现频度/所有苔藓物种频度总和)×100%
2.2.2 生态位宽度研究中一方面将5个群丛综合看作一维资源状态进行分析, 以群丛类型作为资源轴测定物种生态位特征。另一方面将苔藓物种生长基质(树生、石生、土生)作为一维资源状态进行分析, 以不同生长基质类型作为资源轴测定物种生态位特征。以群丛类型作为资源轴时, 反应了苔藓物种在森林群落类型内这个大环境对资源利用程度;以生长基质类型作为资源轴时, 反应了苔藓物种对自身生长基质小环境中对资源的利用程度。本研究采用Shannon-Wiener生态位指数计算生态位宽度[31], 公式如下:

|
(1) |
式中, B为物种i的Shannon-Wiener生态位宽度;Pij是物种i利用第j资源占它利用全部资源位的比例;r为资源位数。其中:
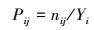
|
(2) |

|
(3) |
式中, nij为物种i在第j资源Yi位的重要值, 为物种i所在利用全部资源位重要值之和。
2.2.3 生态位重叠生态位重叠是指两个种对一定资源位的共同利用程度[7, 32], 本文根据数据特点采用Levins公式[33-34], 其计算公式为:

|
(4) |

|
(5) |
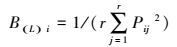
|
(6) |
式中:Lih表示物种i对物种h的生态位重叠值;Lhi表示物种h对物种i的生态位重叠值, 其域值为[0, 1]。BL表示Levins生态位宽度指数。Lih、Lhi的域值[0, 1], BL具域值[1/r, 1]。
3 结果与分析 3.1 苔藓属的重要值分析在以不同群丛类型和以不同生长基质类型作为资源轴的两种情况下分别求重要值, 取两者重要值之和最大的15个属进行分析(表 1, 表 2), 其中叶苔属为苔纲植物其余均为藓纲植物;且在藓纲植物中不仅含有侧蒴藓植物, 还有一定数量顶蒴藓植物。在不同资源位划分情况下, 青藓属和匐灯藓属的重要值均明显高于其他属, 说明这两个属在本区的苔藓群落中占优势地位。
| 序号No. | 属名Genus | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | 总和Total |
| 1 | 青藓属Brachythecium | 15.48 | 22.59 | 24.16 | 15.35 | 23.36 | 100.94 |
| 2 | 匐灯藓属Plagiomnium | 14.49 | 18.61 | 9.75 | 7.51 | 9.08 | 59.44 |
| 3 | 绢藓属Entodon | 5.4 | 11.93 | 5.64 | 5.66 | 2.31 | 30.94 |
| 4 | 鳞叶藓属Taxiphyllum | 3.63 | 10.37 | 6.14 | 3.1 | 2.31 | 25.55 |
| 5 | 羽藓属Thuidium | 4.23 | 2.13 | 3.59 | 5.01 | 7.22 | 22.18 |
| 6 | 提灯藓属Minum | 4.98 | 2.7 | 1.54 | 4.35 | 3.87 | 17.43 |
| 7 | 毛锦藓属Pylaisiadelpha | 2.23 | 4.26 | 3.07 | 0.77 | 2.83 | 13.17 |
| 8 | 美喙藓属Eurhynchium | 3.59 | 0 | 3.07 | 3.28 | 2.31 | 12.24 |
| 9 | 牛舌藓属Anomodon | 4.19 | 0 | 0 | 4.47 | 2.31 | 10.97 |
| 10 | 叶苔属Jungermania | 1.65 | 0 | 0 | 4.41 | 3.87 | 9.93 |
| 11 | 金灰藓属Pylaisiella | 0.75 | 2.7 | 4.61 | 1.37 | 0 | 9.43 |
| 12 | 仙鹤藓属Atrichum | 1.52 | 2.13 | 3.07 | 0 | 2.31 | 9.03 |
| 13 | 凤尾藓属Fissidens | 2.5 | 0 | 4.1 | 0 | 2.31 | 8.91 |
| 14 | 树平藓属Homaliodendron | 1.17 | 0 | 0 | 3.16 | 3.87 | 8.2 |
| 15 | 酸土藓属Oxystegus | 1.62 | 0 | 4.11 | 2.45 | 0 | 8.19 |
| Ⅰ:领春木-葛萝槭-活血丹+白顶早熟禾(Ass.Euptelea pleiospermum-Acer grosseri-Glechoma longituba+Poa acroleuca)群落;Ⅱ:华山松+糙皮桦-东陵山柳-宽叶薹草(Ass.Pinus armandii+Betulautilis-Salix phylicifolia-Carex siderosticta)群落;Ⅲ:蒙古栎-美蔷薇-莎草(Ass.Quercus mongolica-Rosa bella-Cayerus compressus)群落;Ⅳ:华山松+橿子栎+黄栌-连翘-莎草(Ass.Pinus armandii+Quercus baronii var. Baronii+Cotinus coggygria-Forsythia suspensa-Cayerus compressus)群落;Ⅴ:华山松+油松-葛萝槭-求米草(Ass.Pinus armandii+Pinus tabulaeformis-Acer grosseri-Oplismenus undulatifolius)群落 | |||||||
| 序号No. | 属名Genus | X | Y | Z | 总和Total |
| 1 | 青藓属Brachythecium | 16.6459 | 18.8097 | 19.8838 | 55.3394 |
| 2 | 匐灯藓属Plagiomnium | 9.5883 | 12.8489 | 15.9755 | 38.4127 |
| 3 | 绢藓属Entodon | 6.9720 | 4.7390 | 5.1267 | 16.8378 |
| 6 | 提灯藓属Minum | 3.7011 | 7.0971 | 4.6382 | 15.4364 |
| 8 | 美喙藓属Eurhynchium | 3.3396 | 8.6742 | 1.3644 | 13.3783 |
| 5 | 羽藓属Thuidium | 3.3740 | 4.7390 | 4.9905 | 13.1035 |
| 4 | 鳞叶藓属Taxiphyllum | 2.7372 | 6.4245 | 3.9219 | 13.0836 |
| 9 | 牛舌藓属Anomodon | 3.2192 | 0.5644 | 5.1598 | 8.9433 |
| 10 | 叶苔属Jungermannia | 2.7716 | 3.2702 | 1.7848 | 7.8266 |
| 7 | 毛锦藓属Pylaisiadelpha | 3.8216 | 1.6931 | 1.0004 | 6.5151 |
| 13 | 凤尾藓属Fissidens | 0.7747 | 2.8218 | 2.2286 | 5.8250 |
| 15 | 酸土藓属Oxystegus | 1.5493 | 2.4816 | 1.4890 | 5.5199 |
| 12 | 仙鹤藓属Atrichum | 1.7559 | 2.8218 | 0.7163 | 5.2940 |
| 14 | 树平藓属Homaliodendron | 2.1174 | 0 | 1.4208 | 3.5382 |
| 11 | 金灰藓属Pylaisioideae | 2.0830 | 0 | 1.1484 | 3.2314 |
| X:石生基质类型the substrate of rock;Y:土生基质类型the substrate of soil;Z:树生基质类型the substrate of tree | |||||
生态位宽度主要反映出物种对周围资源的利用程度。在以不同群丛类型作为资源轴时(图 1), 生态位宽度大小排序为青藓属、匐灯藓属、提灯藓属、羽藓属、毛锦藓属、绢藓属、鳞叶藓属、美喙藓属、仙鹤藓属、金灰藓属、凤尾藓属、牛舌藓属、酸土藓属、叶苔属、树平藓属依次排列。在以不同生长基质类型作为资源轴时(图 1), 生态位宽度大小排序为青藓属、羽藓属、绢藓属、匐灯藓属、酸土藓属、叶苔属、提灯藓属、鳞叶藓属、凤尾藓属、仙鹤藓属、毛锦藓属、美喙藓属、牛舌藓属、树平藓属、金灰藓属。

|
| 图 1 两种资源轴下苔藓植物生态位宽度 Fig. 1 Niche breadth of bryophytes in two resource axes A:青藓属Brachythecium; B:匐灯藓属Plagiomnium; C:绢藓属Entodon; D:鳞叶藓属Taxiphyllum; E:羽藓属Thuidium; F:提灯藓属Minum; G:毛锦藓属Pylaisiadelpha; H:美喙藓属Eurhynchium; I:牛舌藓属Anomodon; J:叶苔属Jungermannia; K:金灰藓属Pylaisioideae; L:仙鹤藓属Atrichum; M:凤尾藓属Fissidens; N:树平藓属Homaliodendron; O:酸土藓属Oxystegus |
两种资源轴下青藓属生态位宽度最大, 表明该属具有最宽生态适应范围, 对环境资源的利用能力也比较强。树平藓属生态位宽度较小, 说明其生态适应性范围较窄, 适应能力较弱。通过两种不同资源轴方法计算苔藓生态位宽度在排列顺序上存在一定差异, 其中以酸土藓属和叶苔属排列次序尤为明显, 这说明酸土藓属和叶苔属对自身生长小环境的适应性较强, 在不同生长基质类型下均有分布, 但对不同森林群落的环境的适应能力不同, 在某些群丛类型中几乎没有这两个属苔藓植物生长。
3.3 生态位重叠分析不同群丛类型作为资源轴时(表 3, 图 2, 图 3), 生态位重叠值计算发现, 有20对物种对生态位重叠值L>0.20, 约占百分比9.52%, 有140对物种对生态位重叠值0.20>L>0.10, 约占百分比66.67%, 有50对物种对生态位重叠值L<0.10, 约占百分比23.81%。以不同生长基质类型作为资源轴时(表 3), 有101对物种对生态位重叠值L>0.30, 约占百分比48.10%, 有82对物种对生态位重叠值0.30>L>0.20, 约占百分比39.05%, 有27对物种对生态位重叠值L<0.20, 约占百分比12.86%。
| 种对号Species pair | 不同群丛类型Different cluster types | 不同生长基质类型Different growth matrix types | ||
| Lih | Lhi | Lih | Lhi | |
| 1-2 | 0.1939 | 0.1800 | 0.3364 | 0.3247 |
| 1-3 | 0.1944 | 0.1605 | 0.3279 | 0.3200 |
| 1-4 | 0.2019 | 0.1578 | 0.3357 | 0.3002 |
| 1-5 | 0.1919 | 0.1741 | 0.3354 | 0.3285 |
| 1-6 | 0.1821 | 0.1680 | 0.3346 | 0.3122 |
| 1-7 | 0.2050 | 0.1794 | 0.3182 | 0.2451 |
| 1-8 | 0.1825 | 0.1478 | 0.3304 | 0.2245 |
| 1-9 | 0.1631 | 0.0950 | 0.3352 | 0.2409 |
| 1-10 | 0.1765 | 0.0973 | 0.3287 | 0.3130 |
| 1-11 | 0.2075 | 0.1236 | 0.3199 | 0.1978 |
| 1-12 | 0.2112 | 0.1654 | 0.3278 | 0.2664 |
| 1-13 | 0.2054 | 0.1192 | 0.3403 | 0.2860 |
| 1-14 | 0.1828 | 0.0969 | 0.3226 | 0.2081 |
| 1-15 | 0.1888 | 0.1030 | 0.3324 | 0.3150 |
| 2-3 | 0.2036 | 0.1811 | 0.3112 | 0.3147 |
| 2-4 | 0.2059 | 0.1734 | 0.3276 | 0.3034 |
| 2-5 | 0.1622 | 0.1586 | 0.3300 | 0.3348 |
| 2-6 | 0.1771 | 0.1761 | 0.3251 | 0.3143 |
| 2-7 | 0.1976 | 0.1863 | 0.2854 | 0.2276 |
| 2-8 | 0.1568 | 0.1368 | 0.3088 | 0.2174 |
| 2-9 | 0.1581 | 0.0992 | 0.3369 | 0.2508 |
| 2-10 | 0.1396 | 0.0829 | 0.3101 | 0.3059 |
| 2-11 | 0.1856 | 0.1192 | 0.2964 | 0.1899 |
| 2-12 | 0.1875 | 0.1583 | 0.3047 | 0.2565 |
| 2-13 | 0.1641 | 0.1026 | 0.3402 | 0.2962 |
| 2-14 | 0.1391 | 0.0795 | 0.3038 | 0.2031 |
| 2-15 | 0.1505 | 0.0885 | 0.3194 | 0.3135 |
| 3-4 | 0.2021 | 0.1913 | 0.3069 | 0.2812 |
| 3-5 | 0.1316 | 0.1446 | 0.3149 | 0.3160 |
| 3-6 | 0.1495 | 0.1670 | 0.3108 | 0.2972 |
| 3-7 | 0.1778 | 0.1884 | 0.3522 | 0.2779 |
| 3-8 | 0.1273 | 0.1248 | 0.3076 | 0.2142 |
| 3-9 | 0.1248 | 0.0881 | 0.3325 | 0.2448 |
| 3-10 | 0.1108 | 0.0740 | 0.3239 | 0.3160 |
| 3-11 | 0.1909 | 0.1378 | 0.3642 | 0.2308 |
| 3-12 | 0.1602 | 0.1520 | 0.3189 | 0.2655 |
| 3-13 | 0.1211 | 0.0851 | 0.2989 | 0.2574 |
| 3-14 | 0.1039 | 0.0667 | 0.3592 | 0.2375 |
| 3-15 | 0.1438 | 0.0950 | 0.3154 | 0.3062 |
| 4-5 | 0.1219 | 0.1415 | 0.3074 | 0.3367 |
| 4-6 | 0.1318 | 0.1556 | 0.3256 | 0.3397 |
| 4-7 | 0.1792 | 0.2005 | 0.2636 | 0.2270 |
| 4-8 | 0.1141 | 0.1182 | 0.3568 | 0.2712 |
| 4-9 | 0.0924 | 0.0689 | 0.2484 | 0.1996 |
| 4-10 | 0.0849 | 0.0598 | 0.3092 | 0.3292 |
| 4-11 | 0.1978 | 0.1508 | 0.2147 | 0.1485 |
| 4-12 | 0.1691 | 0.1694 | 0.3306 | 0.3004 |
| 4-13 | 0.1310 | 0.0972 | 0.3383 | 0.3180 |
| 4-14 | 0.0826 | 0.0560 | 0.2184 | 0.1576 |
| 4-15 | 0.1393 | 0.0973 | 0.3205 | 0.3396 |
| 5-6 | 0.1856 | 0.1887 | 0.3336 | 0.3178 |
| 5-7 | 0.1611 | 0.1553 | 0.2957 | 0.2325 |
| 5-8 | 0.1910 | 0.1705 | 0.3289 | 0.2282 |
| 5-9 | 0.2041 | 0.1310 | 0.3266 | 0.2396 |
| 5-10 | 0.2263 | 0.1374 | 0.3207 | 0.3117 |
| 5-11 | 0.1352 | 0.0888 | 0.2936 | 0.1854 |
| 5-12 | 0.1688 | 0.1457 | 0.3212 | 0.2665 |
| 5-13 | 0.1857 | 0.1188 | 0.3460 | 0.2969 |
| 5-14 | 0.2342 | 0.1368 | 0.2991 | 0.1970 |
| 5-15 | 0.1631 | 0.0981 | 0.3289 | 0.3182 |
| 6-7 | 0.1612 | 0.1529 | 0.2842 | 0.2346 |
| 6-8 | 0.1909 | 0.1675 | 0.3607 | 0.2627 |
| 6-9 | 0.2289 | 0.1446 | 0.2679 | 0.2063 |
| 6-10 | 0.2176 | 0.1300 | 0.3207 | 0.3272 |
| 6-11 | 0.1302 | 0.0841 | 0.2426 | 0.1608 |
| 6-12 | 0.1524 | 0.1294 | 0.3390 | 0.2952 |
| 6-13 | 0.1586 | 0.0997 | 0.3430 | 0.3090 |
| 6-14 | 0.2149 | 0.1235 | 0.2452 | 0.1695 |
| 6-15 | 0.1560 | 0.0923 | 0.3295 | 0.3346 |
| 7-8 | 0.1386 | 0.1282 | 0.2532 | 0.2586 |
| 7-9 | 0.1128 | 0.0751 | 0.2421 | 0.2259 |
| 7-10 | 0.1162 | 0.0732 | 0.2691 | 0.3327 |
| 7-11 | 0.1927 | 0.1312 | 0.3314 | 0.2662 |
| 7-12 | 0.2015 | 0.1804 | 0.2710 | 0.2860 |
| 7-13 | 0.1775 | 0.1177 | 0.2012 | 0.2196 |
| 7-14 | 0.1249 | 0.0756 | 0.3161 | 0.2649 |
| 7-15 | 0.1416 | 0.0883 | 0.2473 | 0.3043 |
| 8-9 | 0.2036 | 0.1465 | 0.1282 | 0.1355 |
| 8-10 | 0.1882 | 0.1281 | 0.2586 | 0.3623 |
| 8-11 | 0.1442 | 0.1061 | 0.1333 | 0.1213 |
| 8-12 | 0.1427 | 0.1381 | 0.2989 | 0.3574 |
| 8-13 | 0.1924 | 0.1379 | 0.2611 | 0.3230 |
| 8-14 | 0.1827 | 0.1196 | 0.1287 | 0.1222 |
| 8-15 | 0.2060 | 0.1388 | 0.2630 | 0.3667 |
| 9-10 | 0.1833 | 0.1733 | 0.2040 | 0.2703 |
| 9-11 | 0.0503 | 0.0514 | 0.3124 | 0.2689 |
| 9-12 | 0.0663 | 0.0892 | 0.1652 | 0.1868 |
| 9-13 | 0.0908 | 0.0904 | 0.2138 | 0.2501 |
| 9-14 | 0.1745 | 0.1588 | 0.3195 | 0.2869 |
| 9-15 | 0.1108 | 0.1038 | 0.2037 | 0.2686 |
| 10-11 | 0.0413 | 0.0446 | 0.2929 | 0.1903 |
| 10-12 | 0.0678 | 0.0964 | 0.3514 | 0.2999 |
| 10-13 | 0.0784 | 0.0826 | 0.3189 | 0.2815 |
| 10-14 | 0.2011 | 0.1935 | 0.2874 | 0.1948 |
| 10-15 | 0.0880 | 0.0871 | 0.3303 | 0.3287 |
| 11-12 | 0.1419 | 0.1865 | 0.1611 | 0.2117 |
| 11-13 | 0.1419 | 0.1383 | 0.1364 | 0.1853 |
| 11-14 | 0.0387 | 0.0344 | 0.3251 | 0.3392 |
| 11-15 | 0.1748 | 0.1601 | 0.1703 | 0.2609 |
| 12-13 | 0.2038 | 0.1510 | 0.2862 | 0.2960 |
| 12-14 | 0.1092 | 0.0739 | 0.2043 | 0.1623 |
| 12-15 | 0.1539 | 0.1072 | 0.2984 | 0.3480 |
| 13-14 | 0.0908 | 0.0829 | 0.1950 | 0.1497 |
| 13-15 | 0.1602 | 0.1506 | 0.2995 | 0.3377 |
| 14-15 | 0.0733 | 0.0754 | 0.1773 | 0.2604 |

|
| 图 2 群丛类型下生态位重叠值 Fig. 2 Niche overlap value of cluster type |

|
| 图 3 生境基质类型下生态位重叠值 Fig. 3 Niche overlap value of substrate |
两种研究方法中苔藓植物群落生态位宽度较大物种对之间生态位重叠值比较高, 表明生态位宽度大的种群在利用相同资源位时更容易因对资源利用的相似性而发生生态位重叠。但生态位宽度小物种也可能有较大生态位重叠, 如仙鹤藓属与酸土藓属的生态位重叠值均比较大。此外, 生态位宽度大与生态位宽度较小苔藓种群间生态位重叠值Lih与Lhi具有一定的差异, 这主要是由于生态位宽度较大的苔藓植物种群分布较广, 对环境资源利用能力较强所致。
两种研究方法中苔藓植物群落生态位重叠值也具有一定差别。在以不同群丛类型作为资源轴时, 生态位重叠值L>0.15的物种对为46.66%, 在以不同生长基质类型作为资源轴时, L>0.15的物种对为71.43%。多数苔藓物种在相同小环境下生长, 它们对自身生长小环境的资源利用较为一致, 但对森林群落大环境中的资源利用有所差异。另外, 在以不同群丛类型作为资源轴时, 树平藓属和金灰藓属的生态位重叠值Lih与Lhi分别为0.034和0.038, 与相同资源轴下与其他种对相比均为最小;但是在以不同生长基质类型作为资源轴时, 树平藓属和金灰藓属的生态位重叠值Lih与Lhi分别为0.34和0.33, 与相同资源轴下其他种对相比两者均为最大。然而, 与树平藓属和金灰藓属相反, 在以不同群丛类型作为资源轴时, 鳞叶藓属和金灰藓属的生态位重叠值分Lih与Lhi分别为0.20和0.15, 与相同资源轴下其他种对相比均较大;在以不同生长基质类型作为资源轴时, 两者生态位重叠值Lih与Lhi分别为0.21和0.15, 与相同资源轴下其他种对相比均较小。树平藓属和金灰藓属、鳞叶藓属和金灰藓属两个种对在两种不同资源轴下生态位重叠值排序具有完全相反的表现, 出现这一情况的原因可能是因为不同苔藓物种生物学特性存在差异, 对不同尺度上环境中的资源利用有所不同所致, 这种差异有利于物种共存。
4 讨论物种生态位宽度反映了物种对环境资源利用程度和对环境的适应状况, 也表征了物种生态适应性和分布幅度, 即生态位宽度越大, 对环境适应能力越强[35]。以Shannon-Wiener生态位宽度公式对群丛类型和生长基质类型两个资源轴的苔藓植物类群研究, 所得生态位宽度表现出一定差异。两种不同的资源轴分别代表着苔藓植物生长的不同资源维度, 两种资源轴下青藓属生态位宽度最大, 表明该属具有最宽生态适应范围。其次, 匐灯藓属, 绢藓属, 羽藓属也具有较大生态位宽度, 对环境适应性较强。而酸土藓属, 叶苔属在两种资源轴上表现差异很大, 出现这种情况可能由于不同群丛类型下酸土藓属和叶苔属的适应性较弱, 例如在群丛Ⅱ中均没见到这两个属苔藓植物出现, 而在不同生长基质类型下这两个属表现出较强适应性, 这与林思祖等[36]在研究杉阔混交林主要种群在不同资源空间中生态适应性不同这一结论相一致。
生态位重叠是指两个或多个物种共同利用或占有同一资源因素的现象[37]。两种群间较大的生态位重叠值表明两种群对某一资源利用的情况相似, 或者在某些生态因子联系上的相似性。两种资源轴上苔藓植物种对间生态位重叠值差异较大, 生境类型的生态位重叠值明显高于群丛类型, 表明多数苔藓物种在生境类型这一生长小环境中具有相似的要求, 生态位分化较小, 这一观点在陈怡等对地面藓类的研究中所得结论一致[38];而在森林群丛类型这个大环境中资源利用差异较大, 生态位分化较大, 这一特征在郭水良和曹同对长白山地区三种林型下地面生苔藓植物研究中也具有一定体现[17]。在苔藓物种中, 生态位宽度大的物种间生态位重叠值较大;生态位宽度大的物种与生态位宽度较小的物种间也可能有较大的生态位重叠值, 这一研究结果与维管植物的研究相似。
在生长基质上生态位分化较小不利于苔藓物种的共存, 但是不同群落内适应程度的差异增加了苔藓物种的生态位分化, 从而使苔藓物种整体上生态位的分化程度增大, 减小了苔藓物种对相同资源的竞争, 有利于苔藓物种的共存。由于物种在不同资源轴上或多或少的表现出一定的差异, 随着资源轴的增加, 物种之间的生态位分化可能会逐渐增大, 这可能是资源丰富的环境中能够有更多物种共存的原因之一, 但其具体机制有待进一步研究。
5 结论本文应用生态位理论, 对两种资源轴上小秦岭地区苔藓植物生态位宽度和生态位重叠值进行了研究和比较, 结果表明在两种资源轴上苔藓植物生态位特征有明显差异。两种资源轴上青藓属的生态位宽度均表现较大, 而其他苔藓植物则表现具有或多或少的差异, 以酸土藓属和叶苔属生态位宽度差异较大。生态位宽度较大的物种生态位重叠值也较大这一理论在两种资源轴上的苔藓植物种群间同样适用。群丛类型和生境类型下苔藓植物生态位的研究对揭示苔藓植物的种间关系和生态适应性具有重要的意义, 该结果对于揭示森林环境中苔藓植物的存在机制具有积极的意义, 也为苔藓植物生物学特性的研究积累资料。
| [1] | 尚玉昌. 现代生态学中的生态位理论. 生态学进展, 1988, 5(2): 77–84. |
| [2] | Elton C S. Animal Ecology. Chicago: University of Chicago Press, 2001. |
| [3] | Leibold M A. The niche concept revisited mechanistic models and community context. Ecology, 1995, 76(5): 1371–1382. DOI:10.2307/1938141 |
| [4] | 朱春全. 生态位态势理论与扩充假说. 生态学报, 1997, 17(3): 324–332. |
| [5] | May R M. 理论生态学. 孙儒泳, 译. 北京: 科学出版社, 1980: 58-64. |
| [6] | Curtis J T, McIntosh R P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology, 1951, 32(3): 476–496. DOI:10.2307/1931725 |
| [7] | 王刚, 赵松岭, 张鹏云, 陈庆诚. 关于生态位定义的探讨及生态位重叠计测公式改进的研究. 生态学报, 1984, 4(2): 119–127. |
| [8] | Schofield W B. Introduction to Bryology. New York: Macmillan Publishing Company, 1985. |
| [9] | 汪庆, 吴鹏程. 苔藓植物的多样性研究. 生物多样性, 1999, 7(4): 332–339. |
| [10] | 郭水良, 曹同. 长白山森林生态系统树附生苔藓植物分布与环境关系研究. 生态学报, 2000, 20(6): 922–931. |
| [11] | Rambo T R, Muir P S. Forest floor bryophytes of Pseudotsuga menziesii-Tsuga heterophylla stands in Oregon: influences of substrate and overstory. The Bryologist, 1998, 101(1): 116–130. DOI:10.1639/0007-2745(1998)101[116:FFBOPM]2.0.CO;2 |
| [12] | Király I, Nascimbene J, Tinya F, Ódor P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodiversity and Conservation, 2013, 22(1): 209–223. DOI:10.1007/s10531-012-0415-y |
| [13] | 曹同, 郭水良. 长白山主要生态系统苔藓植物的生物多样性研究. 生物多样性, 2000, 8(1): 50–59. |
| [14] | 白学良, 赵连梅, 孙维, 孙卫国. 贺兰山苔藓植物物种多样性、生物量及生态学作用的研究. 内蒙古大学学报:自然科学版, 1998, 29(1): 118–124. |
| [15] | 吴鹏程. 苔藓植物生物学. 北京: 科学出版社, 1998. |
| [16] | 徐晟翀, 曹同, 于晶, 陈怡, 宋国元. 上海市树附生苔藓植物生态位. 生态学杂志, 2006, 25(11): 1338–1343. DOI:10.3321/j.issn:1000-4890.2006.11.008 |
| [17] | 郭水良, 曹同. 长白山主要生态系统地面藓类植物的生态位研究. 生态学报, 2001, 21(2): 231–236. |
| [18] | 叶永忠, 汪万森, 李合申. 河南小秦岭自然保护区科学考察集. 北京: 科学出版社, 2004. |
| [19] | 张凯, 王海亮, 李东伟. 小秦岭国家级自然保护区鸟类调研. 中国林业, 2011(24): 38–38. |
| [20] | 彭舜磊, 王得祥. 秦岭火地塘林区华山松人工林与天然次生林群落特征比较. 西北植物学报, 2009, 29(11): 2301–2311. DOI:10.3321/j.issn:1000-4025.2009.11.023 |
| [21] | 贾宏汝, 陈云, 张旭, 袁志良, 王海亮, 黄群策. 小秦岭自然保护区秦岭冷杉死亡原因. 生态学报, 2016, 36(7): 1936–1945. |
| [22] | 陈云, 王海亮, 韩军旺, 韦博良, 贾宏汝, 叶永忠, 袁志良. 小秦岭森林群落数量分类、排序及多样性垂直格局. 生态学报, 2014, 34(8): 2068–2075. |
| [23] | 高谦, 吴玉环. 中国苔纲和角苔纲植物属志. 北京: 科学出版社, 2010. |
| [24] | 贾渝, 何思. 中国生物物种名录:第一卷:植物, 苔藓植物. 北京: 科学出版社, 2013. |
| [25] | 高贤明, 马克平, 陈灵芝. 暖温带若干落叶阔叶林群落物种多样性及其与群落动态的关系. 植物生态学报, 2001, 25(3): 283–290. |
| [26] | 王伯荪, 李鸣光. 重要值-面积曲线在热带亚热带森林中的应用. 植物生态学报, 1986, 10(3): 161–170. |
| [27] | 马克平. 生物群落多样性的测度方法//钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994: 141-165. |
| [28] | 沈蕾, 郭水良, 宋洪涛, 娄玉霞, 曹同. 样方大小对苔藓植物生态学指标的影响. 广西植物, 2011, 31(2): 198–203. |
| [29] | 贾宏汝. 环境空间异质性对暖温带-北亚热带过渡区植物群落构建的影响[D]. 郑州: 郑州大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10459-1015302376.htm |
| [30] | 王育松, 上官铁梁. 关于重要值计算方法的若干问题. 山西大学学报:自然科学版, 2010, 33(2): 312–316. |
| [31] | 杨效文, 马继盛. 生态位有关术语的定义及计算公式评述. 生态学杂志, 1992, 11(2): 44-49, 35-35. |
| [32] | Colwell R K, Futuyma D J. On the measurement of niche breadth and overlap. Ecology, 1971, 52(4): 567–576. DOI:10.2307/1934144 |
| [33] | 黄英姿. 生态位理论研究中的数学方法. 应用生态学报, 1994, 5(3): 331–337. |
| [34] | Hurlbert S H. The measurement of niche overlap and some relatives. Ecology, 1978, 59(1): 67–77. DOI:10.2307/1936632 |
| [35] | 张桂莲, 张金屯. 关帝山神尾沟优势种生态位分析. 武汉植物学研究, 2002, 20(3): 203–208. |
| [36] | 林思祖, 黄世国, 洪伟, 黄宝龙, 俞新妥. 杉阔混交林主要种群多维生态位特征. 生态学报, 2002, 22(6): 962–968. |
| [37] | 余世孝, 奥罗西L. 物种多维生态位宽度测度. 生态学报, 1994, 14(1): 32–39. |
| [38] | 陈怡, 曹同, 宋国元, 于晶. 上海市地面藓类植物生态位研究. 应用生态学报, 2005, 16(1): 39–43. |