 2017, Vol. 37
2017, Vol. 37文章信息
- 安乐生, 周葆华, 赵全升, 王磊
- AN Lesheng, ZHOU Baohua, ZHAO Quansheng, WANG Lei.
- 黄河三角洲植被空间分布特征及其环境解释
- Spatial distribution of vegetation and environmental interpretation in the Yellow River Delta
- 生态学报. 2017, 37(20): 6809-6817
- Acta Ecologica Sinica. 2017, 37(20): 6809-6817
- http://dx.doi.org/10.5846/stxb201607261518
-
文章历史
- 收稿日期: 2016-07-26
- 修订日期: 2017-02-23





2. 中国海洋大学环境工程系, 青岛 266100;
3. 青岛大学环境科学系, 青岛 266071
2. Department of Environmental Engineering, Ocean University of China, Qingdao 266100, China;
3. Department of Environmental Science, Qingdao University, Qingdao 266071, China
植被空间分布与环境因子之间的关系一直是生态学研究的热点问题[1-3], 其研究可为退化生态系统的恢复与重建提供科学依据[4]。植物群落分布受气候、降水、地形、土壤、地下水等多种自然因素的驱动影响与共同作用。在大尺度上气候对植被分布起着决定性作用, 在小尺度上地形、土壤、生物等因素及其相互作用也影响着植物群落的分布[5]。植被(物种组成数据)-环境(实测或潜在环境因子)关系的梯度分析法(排序)可以半定量地确定影响植物群落类型变化及其分布的关键小尺度环境因子[6]。这种生态统计学上的梯度分析法较为常见的有除趋势对应分析(Detrended Correspondence Analysis, DCA)、典范对应分析(Canonical Correspondence Analysis, CCA)和除趋势典范对应分析(Detrended Canonical Correspondence Analysis, DCCA)等。其中, DCA为非约束性排序(间接梯度分析法), 而CCA和DCCA均为约束性排序(直接梯度分析法), 但DCCA克服了CCA的“弓形效应”[7-8]。
植物的生活与分布深受所在地环境条件的制约。陆生植物植根于土壤, 受土壤中水分和盐分条件的影响尤为显著。滨海地区因海水入侵及沿河道上溯顶托导致地下水咸化、土壤盐渍化引起土壤退化演变, 促进了生物群落的改变及植物群落的演替。黄河三角洲地下水埋藏深度普遍较浅且盐分含量高, 在浅埋条件下, 地下水可通过“饱和带-包气带-植被”间水分的垂向联系由点及面产生极为重要的生态效应[9]。当前, 黄河三角洲地区植被与环境因子关系的研究侧重于东部自然保护区河口湿地植物受地表水深和土壤养分含量的影响[10-11]。本研究基于植被群落空间分布(局地植物样方调查)与植被覆盖(区域遥感影像采集)两个角度综合分析该地区植物空间分布格局与环境因子之间的生态关系, 以期为合理开展黄河下游生态调度、积极维护黄河三角洲滨海湿地生态系统健康提供科学依据。
1 材料与方法 1.1 资料收集与采样布置本研究东、北以海岸线为界, 西、南分别延伸到黄河三角洲北部自然保护区西端和东部自然保护区的南端(118°34′—119°15′E, 37°36′—38°09′N), 总面积约2719.8 km2(图 1)。

|
| 图 1 研究区及采样布置示意 Fig. 1 Distribution of sampling stations in the study area |
收集的环境数据主要是2006—2010年黄河三角洲滨海湿地综合地质调查与评价项目151个采样站位的地形高度、地下水位埋深、潜水氯离子质量浓度[c(Cl-)]、潜水总溶解性固体质量浓度[c(TDS)]及表层土壤氯离子含量[w(Cl-)]、土壤全盐量[w(全盐)]、有机质含量[w(OM)]、pH值等。植被数据包括植被样方调查数据、2008年10月7日TM影像数据1景(来源于“对地观测数据共享计划”)所提取的归一化植被指数(NDVI), 以及同步记录的各站位地表覆被类型(包括翅碱蓬、芦苇、柽柳、人工植被、潮滩等)(图 1)。2008年10月在黄河三角洲东、北部自然保护区内各选10个调查样地, 共计20个样地(S1—S20), 其规格为50 m×50 m。植被样地调查时, 对每个样地的植物种类及其单位面积数量、盖度、多度等进行记录。
1.2 数据处理与分析植被与环境数据排序分析中, 植被数据为20个样地的物种多度值, 环境数据主要包括各样地对应采样站位的地形高度、地下水位埋深、潜水c(Cl-)、潜水c(TDS)及表层土壤w(Cl-)、w(全盐)、w(OM)、pH值等。综合植被与环境数据进行样地的除趋势对应分析(DCA), 根据DCA排序结果, 分析样地主要群落类型(群丛), 判断选择线性还是单峰模型, 再进行典范对应分析(CCA)或除趋势典范对应分析(DCCA), 并绘制二维排序图(最终采用的是DCCA排序)。DCA和DCCA分析均采用Canoco 4.5软件。
NDVI与环境因子回归分析中, 首先通过ENVI 4.8软件提取上述TM影像的NDVI, 之后将其导入ArcGIS 10.2软件中生成NDVI分级分布图, 再利用ArcMap中Extract values to points命令提取其中151个点的NDVI, 最后利用SPSS 17.0软件对相关数据进行单因子相关性分析和多元逐步回归分析[12]。
2 结果与讨论 2.1 典型植被及其空间分布研究区主要植物种有芦苇(Phragmites australis)、柽柳(Tamarix chinensis)、碱蓬(Suaeda glauca)、白茅(Imperata cylindrical Var.major)、獐茅(Aeluropus littoralis var.sinensis)、罗布麻(Apocynum venetum)、香蒲(Typha angustifolia)、荻(Miscanthus sacchari)、野大豆(Glycine soja)、杞柳(Salix integra)、补血草(Limonium sinense)及狗尾草(Setaria viridis)等。黄河三角洲自然保护区共有种子植物42科、390多种, 以禾本科、豆科、菊科、藜科居多, 植被覆盖率约为55%, 其中典型植被翅碱蓬、柽柳和芦苇分布较广。
基于植被群落样方调查及野外采样记录, 结合遥感影像资料, 并参考崔保山和杨志峰等[13]对黄河三角洲湿地植被分布的研究可知, 区内典型植被主要分布在保护区内, 在东部自然保护区, 芦苇、柽柳、盐地碱蓬(翅碱蓬)沿黄河河道呈条带状分布, 沿河道向外至高潮线附近依次是芦苇、柽柳和翅碱蓬, 各植被带宽度约0.5—4.0 km(翅碱蓬偏窄)、长度从入海口以上5.0 km沿河道至大汶流管理站约45.0 km;在北部自然保护区, 翅碱蓬、柽柳、芦苇自南向北呈斑块状集中分布于一千二管理站东侧和东北侧, 自孤北水库至防潮坝依次是芦苇、柽柳和翅碱蓬, 各植被带宽度约1.0—7.5 km(芦苇偏窄)、长度约10.5—22.0 km[9]。区域地表覆被呈现2条较为明显的生态演替序列, 一是横向上由河道向两侧发育的“芦苇/香蒲/河道防护林-蒿草/獐茅-杞柳-耕地植被”, 二是纵向上沿黄河现行流路由海向陆发育的“盐地碱蓬-柽柳-蒿草/獐茅-耕地植被”, 2条生态序列时空交错, 且在东部保护区表现得更为明显[14]。
2.2 植被群落分布格局与关键环境因子的关系 2.2.1 样地的DCA排序与分类采用DCA对20个样地进行分析, 结果见图 2。DCA前4个排序轴的特征值(表示物种数据异质性)分别为0.772、0.361、0.191、0.084。第一排序轴特征值最大, 包含的生态信息量最多, 第二、三、四排序轴依次减小。一般DCA前2个轴累积贡献率若达到70%, 可反映植被的基本面貌。本研究DCA前2个轴的累积贡献率为75.1%, 即可以解释75.1%的物种数据信息(物种总数为14), 表明DCA排序结果比较理想。

|
| 图 2 20个样地的DCA二维排序 Fig. 2 DCA ordination of 20 sample plots Ⅰ—Ⅳ:群丛;1—20:样地 |
参考宋创业等[15]、贺强等[16]利用双向指示种分析法(TWINSPAN)对黄河三角洲湿地植被群落分类的结果, 结合本次DCA分析, 可将20个样地划分为4组, 代表 4个主要群落类型(群丛)(表 1)。
| 序号 Sequence number |
名称 Name |
包含样地 Sample plots |
分布特征 Distribution characteristics |
盖度 Coverage |
主要伴生种 Companion species |
| 群丛Ⅰ Association Ⅰ |
翅碱蓬群落(Ass. Suaeda heteroptera) | 样地3、6、7、8、9、10、11 | 陆地向滩涂延伸的先锋植物群落, 主要分布在平均海潮线以上的近海滩地, 尤以黄河入海口最为集中 | 翅碱蓬:15%—82% | 柽柳和芦苇, 偶见补血草和獐茅, 其它物种寥寥 |
| 群丛Ⅱ Association Ⅱ |
柽柳-翅碱蓬群落(Ass. Tamarix chinensis-Suaeda heteroptera) | 样地4、5 | 潮间带中上部分的盐碱荒地, 土壤盐分含量较高(低于翅碱蓬群落) | 柽柳:20%—50% 翅碱蓬:10%—48% |
芦苇和獐茅, 散布有少量的补血草、茵陈蒿和苣荬菜等 |
| 群丛Ⅲ Association Ⅲ |
芦苇-柽柳群落(Ass. Phragmites australis-Tamarix chinensis) | 样地2、12、14、15、17、18 | 潮间带上部分的高地及潮上带, 土壤盐分含量较柽柳-翅碱蓬群落低 | 芦苇:8%—90% 柽柳:5%—40% |
翅碱蓬, 白茅、罗布麻和茵陈蒿也较为常见 |
| 群丛Ⅳ Association Ⅳ |
芦苇群落(Ass. Phragmites australis) | 样地1、13、16、19、20 | 潮上带地区, 土壤盐分含量较低 | 芦苇:60%—90% | 柽柳和翅碱蓬, 白茅、香蒲、荻等出现频率也较高 |
| 变量 Variables |
归一化植被指数NDVI | 地下水位Groundwater table | 地形高度Terrain height | 潜水c(Cl-)Cl- concentration in phreatic aquifer | 表层土壤w(Cl-)Cl- content in surface soil | 表层土壤w(OM)OM content in surface soil | 表层土壤pH值pH value of surface soil |
| 归一化植被指数NDVI | 1 | ||||||
| 地下水位Groundwater table | 0.468** | 1 | |||||
| 地形高度Terrain height | 0.330** | 0.665** | 1 | ||||
| 潜水c(Cl-) Cl- concentration in phreatic aquifer | -0.737** | -0.400** | -0.316** | 1 | |||
| 表层土壤w(Cl-) Cl- content in surface soil | -0.740** | -0.423** | -0.360** | 0.643** | 1 | ||
| 表层土壤w(OM) OM content in surface soil | 0.148 | 0.093 | 0.178* | -0.154 | -0.107 | 1 | |
| 表层土壤pH值pH value of surface soil | 0.000 | -0.040 | -0.070 | 0.024 | -0.160 | -0.441** | 1 |
| *:0.05的显著性水平Significant at the 0.05 level;**:0.01的显著性水平Significant at the 0.05 level | |||||||
DCA排序的横、纵轴显示不同的生态意义。图 2显示, 群丛主要沿着第一轴表现出明显的梯度变化, 自左向右分别对应翅碱蓬、柽柳、芦苇等植被类型, 初步表明第一轴包含潜水c(TDS)、土壤w(全盐)、地形高度和地下水位等信息, 且第一轴正向表示的是地形高度和地下水埋深的增加、潜水c(TDS)和土壤w(全盐)的减小。沿第二排序轴, 群丛Ⅰ—Ⅲ基本位于同一水平线上, 而群丛Ⅳ的样地在第二轴方向上呈条带状分布, 进一步对样地1、13、16、19、20中环境因子进行比较分析发现, pH值相对于其他环境因子变化最为明显, 说明第二轴主要反映土壤pH值的变化。
DCA分析结果中梯度长度(表示物种变化范围)的第一排序轴长度为3.517, 介于3—4之间, 说明梯度分析排序模型采用单峰或线性模型均可。考虑到各样地物种多度统计结果中零值较多, 且DCA排序结果显示物种分布受环境梯度影响显著, 因此物种分布与环境因子关系分析选择基于单峰模型的排序方法, 同时, 为消除第一、二排序轴相关而产生的“弓形效应”,采用非线性的DCCA模型纠偏。
2.2.2 关键环境因子对植物分布格局的影响通过筛选确定了地形高度、地下水位埋深、潜水c(Cl-)及表层土壤w(Cl-)、w(OM)、pH值共6个环境因子作为环境信息的代理变量, 应用DCCA排序对黄河三角洲地区20个样地的物种和环境数据进行分析, 得到物种多度与环境因子的排序图(图 3)。

|
| 图 3 物种多度与环境因子的DCCA二维排序 Fig. 3 DCCA ordination of species abundance and environmental factors 排序轴与环境因子间的相关系数用箭头线的长度和方向来表示; GE:地形高度(地面标高) Ground elevation;GWD:地下水位埋深Depth of groundwater table;GWCl:潜水c(Cl-) Cl- concentration in phreatic aquifer;SCl:表层土壤w(Cl-) Cl- content in surface soil;SOM:表层土壤w(OM) OM content in surface soil;pH:表层土壤pH值pH value of surface soil; △:群丛Ⅰ样地Association Ⅰ plots;✍:群丛Ⅱ样地Association Ⅱ plots;□:群丛Ⅲ样地Association Ⅲ plots;○:群丛Ⅳ样地Association Ⅳ plots (符号的大小分别代表物种多度值);+:物种Plant species;Phragmit:芦苇Phragmites australis;Suaeda:翅碱蓬Suaeda heteroptera;Tamarix:柽柳Tamarix chinensis;Aelurop:獐茅Aeluropus littoralis var.sinensis;Limon:补血草Limonium sinense;Sonchus:苣荬菜Sonchus brachyotu;Miscan:荻Miscanthus sacchari;Imperata:白茅Imperata cylindrical Var.major;Typha:香蒲Typha orientalis Presl;Apocynum:罗布麻Apocynum venetum;Artemis:茵陈蒿Artemisla capillaris;Glycine:野大豆Glycine soja;Melilot:草木樨Melilotus officinalis;Setaria:狗尾草Setaria viridis |
结合表 2可看出, DCCA排序4个轴的特征值分别为0.727、0.242、0.137、0.086, 前两轴的物种与环境相关性较高(R分别为0.972和0.842), 共解释了物种-环境关系总方差的72.9%, 说明排序效果较好[16]。
| 项目Items | 排序轴Ordination axes | |||
| 1 | 2 | 3 | 4 | |
| 特征值Eigenvalues of each ordering axis | 0.727 | 0.242 | 0.137 | 0.086 |
| 物种-环境相关性 Correlation between plant species and its main environmental factors |
0.972 | 0.842 | 0.544 | 0.742 |
| 物种-环境关系累积百分比方差 Cumulative percentage variance of relationship between species and environment variables |
54.7 | 72.9 | 83.2 | 89.7 |
DCCA排序图中环境因子用带箭头的连线表示, 线条的长度及其与排序轴的夹角反映某个环境因子与排序轴相关性的大小, 且长度越长、夹角越小, 相关系数的绝对值越大[17-19]。图 3显示, 代表潜水c(Cl-)和表层土壤w(Cl-)的线条长度较长, 且与第一轴夹角很小, 地形高度和地下水位埋深次之, 表明潜水c(Cl-)和表层土壤w(Cl-)与第一轴相关性最高(R分别为-0.800和-0.938, 表 3), 其次是地形高度和地下水位埋深(R分别为-0.623和-0.606)。表层土壤w(OM)虽然与第一轴夹角较小, 但其连线较短, 故其与第一轴的相关性并不高(R为0.270)。同时, 第二轴只有代表pH值的连线向其靠近, 且相关性较低(R为0.287), 表明第二轴包含的环境信息很少, 且反映程度较弱。从图 3和表 3不难看出, DCCA排序第一轴侧重反映的是表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深等关键环境因子, 而第二轴在一定程度上代表着表层土壤pH值。
| 排序轴 Ordination axes |
地形高度 Terrain height |
地下水位 Groundwater table |
潜水c(Cl-) Cl- concentration in phreatic aquifer |
表层土壤w(Cl-) Cl- content in surface soil |
表层土壤w(OM) OM content in surface soil |
表层土壤pH值 pH value of surface soil |
| 1 | 0.623** | 0.606** | -0.800** | -0.938** | 0.270** | -0.242* |
| 2 | -0.104 | 0.015 | -0.211 | -0.064 | 0.023 | 0.287** |
| *:0.05的显著性水平Significant at the 0.05 level;**:0.01的显著性水平Significant at the 0.01 level | ||||||
DCCA二维点图中各群落类型主要沿第一排序轴变化, 且排列次序与DCA一致, 虽然DCCA排序图中群丛Ⅲ和Ⅳ略有交错, 但其分布仍较集中且相对位置明确, 表明DCCA和DCA排序图总体上具有相似性, 研究区主要群落类型在2种排序图上的分布均反映了植物群落在环境梯度上的变化趋势。与DCA相比, DCCA清晰地表明其第一排序轴主要代表的是潜水c(Cl-)等关键水盐因子, 且随着潜水c(Cl-)和表层土壤w(Cl-)的减小, 群丛由翅碱蓬群落逐渐向芦苇群落演变。
与样地相比, 物种在DCCA二维排序空间中的布局较为分散, 这主要是由于样地中存在一些物种出现频度低、多度值小, 在排序时向排序轴边缘靠近。总体来看, 优势种和主要伴生种分布相对集中, 且沿第一排序轴有明显变化, 自左向右依次为翅碱蓬、补血草、柽柳、獐茅、荻、芦苇、白茅、香蒲、野大豆等。翅碱蓬、柽柳和芦苇在二维空间中的具体位置表明, 各群丛类型与其优势种的分布格局基本相似, 如以翅碱蓬为优势种的群丛Ⅰ(翅碱蓬群落), 同翅碱蓬一样位于DCCA排序图的最左端, 这也说明优势种的分布格局在一定程度上决定着植物群落类型的分布格局[20]。
总之, DCCA第一排序轴集中了排序的大部分信息, 突出反映了表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深等水盐环境梯度。样地、物种主要沿第一排序轴有序变化充分表明, 表层土壤w(Cl-)、潜水c(Cl-)和地下水位是决定区域典型植被群落分布格局的关键因子[12]。
2.3 NDVI与关键环境因子的关系前已述及, 研究区典型植被群落分布格局受表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深等关键水盐因子影响显著。将这3个关键因子空间分布与区域NDVI空间分布进行比较发现, NDVI沿地下水径流方向总体呈递减趋势, 与3个关键因子在空间分布上具有较高的相似性[9]。由此可见, NDVI和3个关键水盐因子之间存在较高的相关性。
结合151个采样站位表层土壤w(Cl-)等6个环境因子对NDVI的散点图, 经正态检验后剔除12个离群值, 最终对139个点的相应数据进行单因子相关性分析和多元逐步回归分析, 结果见公式1—3、表 4和图 4。
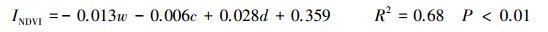
|
(1) |

|
(2) |
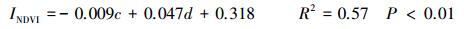
|
(3) |

|
| 图 4 NDVI与主要环境因子关系 Fig. 4 Relationship between NDVI and main environmental factors |
式中, INDVI为归一化植被指数;w为表层土壤w(Cl-), g/kg;c为潜水c(Cl-), g/L;d为地下水位埋深(m)。
从NDVI与6个环境因子的相关性可看出, NDVI与地下水位埋深、地形高度、表层土壤w(Cl-)、潜水c(Cl-)间具有极显著的相关性, 且与地下水位埋深、地形高度呈正相关, R分别为0.468、0.330;与表层土壤w(Cl-)、潜水c(Cl-)呈负相关, R分别为-0.740、-0.737。通过数据拟合发现, NDVI与表层土壤w(Cl-)、潜水c(Cl-)间比较适合用Logistic曲线来拟合, R2分别达到0.65、0.60, 而NDVI与地下水位埋深、地形高度间用线性模型拟合即可, R2分别达到0.22、0.11(图 4)。上述结果表明, 研究区NDVI主要受表层土壤w(Cl-)、潜水c(Cl-)显著影响。
多元逐步回归分析通过3步运行依次将表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深引入回归模型, 而剔除了地形高度、表层土壤w(OM)及pH值3个对NDVI影响较弱的环境因子, 最终建立了三元线性回归方程(式1), 且R2达到0.66的较高水平。再次表明, 表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深3个水盐因子是影响区域NDVI的关键环境因子。同时显示, 仅引入表层土壤w(Cl-)、潜水c(Cl-)2个变量的情况下, 得到的二元回归方程R2与三元回归方程相比只差0.02, 回归效果依然显著, 这突出反映了水土环境系统中盐分因子对NDVI的决定作用[12]。
对表 4和图 4进一步分析发现, 表层土壤w(Cl-)与潜水c(Cl-)相关性较高, R为0.643, 此外, 图 4显示NDVI与潜水c(Cl-)间Logistic模型与图 4中NDVI与表层土壤w(Cl-)间Logistic模型的R2相差无几(二者R2分别为0.66和0.60)。研究区潜水借助包气带自身的水势梯度或植物蒸腾作用向土壤输送水分, 进而控制着土壤Cl-的空间分布与变异[21]。因而, 植被群落的空间分布和NDVI的空间变异实质上受潜水c(Cl-)控制。值得注意的是, 研究区地下水埋深集中在0.5—2.5 m以下, 绝大部分区域地下水位已超过临界埋深, 致使区内潜水Cl-向上迁移与累计成为普遍现象, 地下水位埋深直接关系到潜水Cl-向上迁移与否, 因而地下水位埋深在影响“饱和带-包气带-植被”系统水盐迁移等方面的重要生态作用决不能忽视。可见, 地下水位埋深和潜水c(Cl-)是影响黄河三角洲植被生长与分布的主导环境因子, 尤其是潜水c(Cl-)由海向陆、由河道向两侧的梯度变化对翅碱蓬、柽柳、芦苇等天然植被分布格局起重要的控制作用。
3 结论(1) DCA二维排序表明黄河三角洲植物群落主要沿着能反映潜水c(TDS)、土壤w(全盐)和地下水位变化等信息的第一排序轴变化, 且按梯度顺序可划分为翅碱蓬、柽柳-翅碱蓬、芦苇-柽柳、芦苇4个主要群落类型(群丛)。DCCA排序二维点图和DCA排序图总体相似, 但与DCA相比, DCCA清晰地表明其第一轴主要代表的是潜水c(Cl-)等关键水盐因子, 且随着潜水c(Cl-)和表层土壤w(Cl-)的减小, 群丛由翅碱蓬群落逐渐向芦苇群落演变。DCCA排序图中环境因子线条长度及其与第一轴的夹角大小也进一步揭示地下水位、潜水c(Cl-)及表层土壤w(Cl-)对该地区典型植被群落分布格局影响最为显著。
(2) 环境因子与植被覆盖度(NDVI)的单因子相关性分析显示黄河三角洲NDVI主要受表层土壤w(Cl-)、潜水c(Cl-)显著影响, 其次是地下水位埋深、地形高度, 而受表层土壤w(OM)及pH值影响比较小。多元回归分析表明表层土壤w(Cl-)、潜水c(Cl-)和地下水位埋深3个水盐因子主导区域NDVI的空间分布。但黄河三角洲地下水埋藏较浅, 区域土壤w(Cl-)空间分布与变异受地下水位埋深和潜水c(Cl-)二者共同作用, 因此, 土壤w(Cl-)的植被效应实际上还是受地下水位埋深和潜水c(Cl-)的影响。
(3) 黄河三角洲地区地下水是影响植被生长与分布的敏感要素, 而潜水c(Cl-)和地下水位埋深是地下水这一生态环境敏感要素中的2个关键因子, 尤其是潜水c(Cl-)梯度变化对天然植被分布格局起重要的控制作用。在加强黄河三角洲滨海湿地保护、维护湿地生态系统健康过程中, 应重视河口与近岸地区故道断流、岸线蚀退、海水顶托等导致潜水c(Cl-)升高对植被的不利影响, 充分发挥黄河下游生态调度的“压咸补淡”作用, 促进滨海湿地植被顺向演替发展。
| [1] | Clark C M, Cleland E E, Collins S L, Fargione J E, Gough L, Gross K L, Pennings S C, Suding K N, Grace J B. Environmental and plant community determinants of species loss following nitrogen enrichment. Ecology Letters, 2007, 10(7): 596–607. DOI:10.1111/j.1461-0248.2007.01053.x |
| [2] | 张庆, 牛建明, BuyantuyevA, 丁勇, 康萨如拉, 王凤兰, 张艳楠, 杨艳, 韩砚君. 内蒙古短花针茅群落数量分类及环境解释. 草业学报, 2012, 21(1): 83–92. DOI:10.11686/cyxb20120111 |
| [3] | 李思阳, 张娟, 姚玲爱, 胡芳, 许振成, 赵学敏. 西枝江流域浮游植物群落结构特征与主要环境因子的关系研究. 环境科学学报, 2016, 36(6): 1939–1947. |
| [4] | Zuo X A, Zhao X Y, Zhao H L, Zhang T H, Li Y L, Wang S K, Li W J, Powers R. Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China. Geoderma, 2012, 173-174(3): 1–9. |
| [5] | 刘瑞雪, 陈龙清, 史志华. 丹江口水库水滨带植物群落空间分布及环境解释. 生态学报, 2015, 35(4): 1208–1216. |
| [6] | Hopfensperger K N, Burgin A J, Schoepfer V A, Helton A M. Impacts of saltwater incursion on plant communities, anaerobic microbial metabolism, and resulting relationships in a restored freshwater wetland. Ecosystems, 2014, 17(5): 792–807. DOI:10.1007/s10021-014-9760-x |
| [7] | 胡芳, 许振成, 姚玲爱, 陈晓燕, 马千里, 张杏杏, 王丽, 赵学敏. 剑潭水库浮游植物群落特征与水环境因子关系研究. 环境科学学报, 2014, 34(4): 950–958. |
| [8] | 周欣, 左小安, 赵学勇, 王少昆, 刘川, 张婧, 吕朋, 张建鹏. 科尔沁沙地植物群落分布与土壤特性关系的DCA、CCA及DCCA分析. 生态学杂志, 2015, 34(4): 947–954. |
| [9] | 安乐生, 赵全升, 叶思源, 刘贯群, 丁喜桂. 黄河三角洲地下水关键水盐因子及其植被效应. 水科学进展, 2011, 22(5): 689–695. |
| [10] | Cui B S, Yang Q C, Yang Z F, Zhang K J. Evaluating the ecological performance of wetland restoration in the Yellow River Delta, China. Ecological Engineering, 2009, 35(7): 1090–1103. DOI:10.1016/j.ecoleng.2009.03.022 |
| [11] | 赵欣胜, 吕卷章, 孙涛. 黄河三角洲植被分布环境解释及柽柳空间分布点格局分析. 北京林业大学学报, 2009, 31(3): 29–36. |
| [12] | 安乐生. 黄河三角洲地下水水盐特征及其生态效应. 青岛:中国海洋大学, 2012: 75–90. |
| [13] | 崔保山, 杨志峰. 湿地学. 北京:北京师范大学出版社, 2006: 85–90. |
| [14] | 叶庆华, 田国良, 刘高焕, 叶景敏, 娄维国. 黄河三角洲新生湿地土地覆被演替图谱. 地理研究, 2004, 23(2): 257–264. |
| [15] | 宋创业, 刘高焕, 刘庆生, 曹铭昌, 黄翀. 黄河三角洲植物群落分布格局及其影响因素. 生态学杂志, 2008, 27(12): 2042–2048. |
| [16] | 贺强, 崔保山, 赵欣胜, 付华龄, 廖晓琳. 黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系. 生态学报, 2009, 29(2): 676–687. |
| [17] | Franklin J, Wiser S K, Drake D R, Burrows L E, Sykes W R. Environment, disturbance history and rain forest composition across the islands of Tonga, Western Polynesia. Journal of Vegetation Science, 2006, 17(2): 233–244. DOI:10.1111/j.1654-1103.2006.tb02442.x |
| [18] | 李新荣, 谭会娟, 何明珠, 王新平, 李小军. 阿拉善高原灌木种的丰富度和多度格局对环境因子的变化响应——极端干旱荒漠区灌木多样性保育的前提. 中国科学D辑:地球科学, 2009, 39(4): 504–515. |
| [19] | Lai J S, Mi X C, Ren H B, Ma K P. Species-habitat associations change in a subtropical forest of China. Journal of Vegetation Science, 2009, 20(3): 415–423. DOI:10.1111/jvs.2009.20.issue-3 |
| [20] | 朱军涛, 于静洁, 王平, 王志勇. 额济纳荒漠绿洲植物群落的数量分类及其与地下水环境的关系分析. 植物生态学报, 2011, 35(5): 480–489. |
| [21] | 安乐生, 周葆华, 赵全升, 刘贯群, 朱磊. 黄河三角洲土壤氯离子空间变异特征及其控制因素. 地理科学, 2015, 35(3): 358–364. |