 2017, Vol. 37
2017, Vol. 37文章信息
- 覃扬浍, 马姜明, 梅军林, 杨栋林, 庄枫红, 苏静
- QIN Yanghui, MA Jiangming, MEI Junlin, YANG Donglin, ZHUANG Fenghong, SU Jing.
- 漓江流域岩溶区檵木群落不同恢复阶段凋落物分解初期动态
- The initial dynamic of litter decomposition of Loropetalum chinense communities amomg different recovery stages in karst area of Lijiang river watershed
- 生态学报. 2017, 37(20): 6792-6799
- Acta Ecologica Sinica. 2017, 37(20): 6792-6799
- http://dx.doi.org/10.5846/stxb201606291301
-
文章历史
- 收稿日期: 2016-06-29
- 网络出版日期: 2017-06-01





2. 广西珍稀濒危动物生态学重点实验室, 桂林 541004;
3. 岩溶生态与环境变化研究广西高校重点实验室, 桂林 541004
2. Guangxi Key Laboratory of Rare and Endangered Animal Ecology, Guangxi Normal University, Guilin 541004, China;
3. University Key Laboratory of Karst Ecology and Environmental Change of Guangxi Province (Guangxi Normal University), Guilin 541004, China
凋落物分解是陆地生态系统元素转移和C循环的重要过程, 是大气中CO2的重要来源[1-2]。凋落物分解释放的养分对维持土壤肥力具有重要作用, 可反映森林生态系统与环境之间相互作用的强度[3-4]。目前, 国内外学者对凋落物的分解开展了大量的研究, 分解方式包括单一物种和混合凋落叶分解, 研究对象包括植被类型、气候变化、凋落物自身特性、土壤生物等[5]。研究表明, 凋落物分解主要受气候、凋落物基质质量、分解者三方面的影响[6]。国内学者在植物群落恢复演替过程中凋落物分解已开展相应的研究, 如王敏英等[7]在海南丘陵植被恢复凋落物分解研究表明, 凋落物初始N对凋落物分解速率影响较大。另有研究表明, 随着植被恢复正向演替, 凋落物分解速率逐渐增大[8]。森林植被恢复演替不同阶段的土壤状况有所差异, 从而导致凋落物基质质量的差异, 这种差异对凋落物分解的快慢起着决定性的影响[9]。凋落物分解的快慢决定着土壤养分供应能力的大小, 影响着群落内树种间的竞争以及生态系统的生产力, 进而促进或延缓了植被的恢复演替进程[10]。
岩溶(喀斯特)环境是一种脆弱的生态环境, 岩溶生态系统抗干扰能力低, 石漠化是岩溶区土壤荒漠化的表现形式之一[11-12]。岩溶区地表凋落物储量、分解程度和组分等控制着表层土有机质、有效N、P、K、Mn、Zn等有效态的含量, 可以促进碳酸盐岩的成土速率[11], 对喀斯特森林土壤发育和改良有重要意义[13]。目前, 有关岩溶区凋落物方面的研究主要集中在凋落物凋落动态[14]、凋落物养分归还特征[15]和凋落物化学计量[16]等。檵木(Loropetalum chinense)群落为漓江流域岩溶区分布广泛的一种典型次生林类型。随着自然恢复演替的进行, 形成了檵木群落的灌木阶段、乔灌阶段和乔林阶段[17-18]。本研究以漓江流域岩溶区不同恢复阶段檵木群落凋落物分解为切入点, 探讨檵木群落演替过程中凋落物分解基质质量动态变化, 研究结果为探究岩溶区植被恢复演替过程中的物质循环规律提供科学依据。
1 研究区概况和研究方法 1.1 研究区概况研究地区位于广西壮族自治区桂林市西村, 为漓江流域核心地段, 地处广西东北部, 地理坐标为109°36′—111°29′E, 24°15′—26°23′N。属中亚热带湿润季风气候, 气候温和, 雨量充沛, 海拔100—500m。年平均气温18.9℃, 最冷的1月份平均气温7.8℃, 最热的7月份平均气温28℃;全年无霜期300d;年平均降雨量1949.5mm, 降雨量年分配不均, 秋、冬季干燥少雨;年平均蒸发量1490—1905mm[19]。本研究以不同恢复阶段檵木群落作为研究对象[19], 檵木群落样方概况见表 1。
| 恢复阶段 Restoration stage |
坡向 Slope aspect |
坡度/(°) Slope gradient/° |
海拔/m Altitude |
岩石裸露率/% Rock bare rate /% |
平均地径(胸径)/cm Mean ground dinameter(DBH) |
群落高度/m Mean height of community |
主要优势种 Main dominant Species |
| 灌木阶段Shrub stage | E | 15—20 | 150—200 | 30—40 | 0.75a | 1.16 | 檵木、龙须藤、红背山麻杆、蔓生莠竹 |
| 乔灌阶段Shrub-tree stage | E | 20—25 | 200—250 | 30—35 | 3.91b | 3.88 | 檵木、小巴豆、紫凌木、狭穗薹草、庐山香科科 |
| 乔林阶段Tree stage | SE, E | 15—20 | 220—280 | 25—30 | 4.48b | 4.35 | 檵木、小巴豆、龙须藤、狭穗薹草 |
| a:地径Mean ground diameter of shrub at shrub stage, b:胸径Mean diameter at breast height of tree at shrub-tree stage or tree stage | |||||||
2014年4月初, 在檵木群落3个恢复阶段固定样地中分别设置3个20m×20m的样方[18], 每个样方分别随机设置6个50cm×50cm的尼龙网收集框(孔径1mm), 离地面50cm, 每月的月初收集框内凋落物, 带回实验室于80℃条件下烘干。在2014年8月, 分别收集3个恢复阶段新鲜凋落未分解的凋落物, 带回实验室烘干, 分别与前几个月收集(收集框)的凋落物混合, 每个恢复阶段准确称取20g于15cm×25cm(孔径1mm)的凋落物尼龙网袋内, 每个恢复阶段分别为36袋, 共计108袋。于2014年9月初放置在原样地内分解, 每个样方内分别随机放置12袋并编号, 自放置之日起每3个月每个恢复阶段收回3袋, 共9袋, 带回实验室清除附着在凋落物上的沙土和侵入的根系, 每个恢复阶段其中3袋于80℃条件下烘干称重, 计算损失质量, 粉碎, 用于测定基质质量。
用重铬酸钾外加热法测定凋落物有机碳含量[19];用全自动凯氏定氮仪(Kjeltec TM 8400)测定凋落物全N含量;用钼锑抗比色法测定凋落物全P含量[20]。用浓硫酸水解和重铬酸钾氧化测定凋落物的纤维素含量;用醋酸分离和硫酸水解法测定凋落物的木质素含量[21]。
1.2.2 数据处理凋落物分解剩余率计算公式:
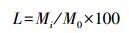
|
式中, L为凋落叶分解干质量剩余率(%), Mi为凋落物分解t时间后样品干重, M0为凋落物初始重量。
凋落物分解率公式:
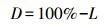
|
式中, D为凋落物分解率(%), L为凋落物分解剩余率(%)。
凋落物分解剩余率采用修正的Olson指数衰减模型进行拟合[22]:
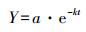
|
Y为凋落物残留率(%), t为凋落物分解时间, a为拟合参数, k为凋落物年分解系数。
数据分析采用Excel 2010和SPSS 19.0软件, 绘图采用Sigmaplot 10.0软件。
2 结果与分析 2.1 凋落物分解动态由图 1可知, 檵木群落不同恢复阶段凋落物经过1a分解后, 灌木阶段、乔灌阶段和乔林阶段凋落物剩余率分别为59.58%, 61.79%和62.02%。檵木群落不同恢复阶段凋落物分解速率差异显著(P<0.05), 且灌木阶段>乔灌阶段>乔林阶段。檵木群落灌木阶段和乔灌阶段凋落物分解速率均在3—6月份达最快, 乔林阶段凋落物分解速率在6—9月份达最快。3个不同恢复阶段凋落物均在12月份到次年3月份其分解速率相对较慢。由表 2可知, 檵木群落灌木阶段、乔灌阶段和乔林阶段凋落物分解50%所需的时间分别为1.28a、1.38a、1.41a, 分解95%所需的时间分别为5.54a、5.97a和6.09a。凋落物年分解系数k大小顺序为:灌木阶段(0.541)>乔灌阶段(0.502)>乔林阶段(0.492)。

|
| 图 1 檵木群落不同恢复阶段凋落物分解动态 Fig. 1 Litter decomposition dynamic in different restoration stages of Loropetalum chinense communities 凋落物分解时间90d, 180d, 270d, 360d分别对应2014年12月、2015年3月、2015年6月、2015年9月 |
| 恢复阶段 Restoration stage |
回归方程 Regression equation |
R2 | k | t0.5/a | t0.95/a |
| 灌木阶段Shrub stage | y=102.00e-0.541t | 0.9957 | 0.541 | 1.28 | 5.54 |
| 乔灌阶段Shrub-tree stage | y=103.63e-0.502t | 0.9816 | 0.502 | 1.38 | 5.97 |
| 乔林阶段Tree stage | y=105.25e-0.492t | 0.9416 | 0.492 | 1.41 | 6.09 |
檵木群落不同恢复阶段凋落物除纤维素外其他初始养分含量差异显著(P < 0.05)。灌木、乔灌阶段C、P含量差异均不显著(P < 0.05), 但均显著大于乔灌阶段(P < 0.05)。灌木阶段N含量显著大于乔灌阶段和乔林阶段(P < 0.05)。乔林阶段木质素含量显著大于乔灌、灌木阶段(P < 0.05)。
| 恢复阶段 Restoration stage |
C/% | N/% | P/(g/kg) | 木质素/% Lignin/% |
纤维素/% Cellulose/% |
| 灌木阶段Shrub stage | 55.45±0.56a | 1.19±0.05a | 0.51±0.05a | 39.80±6.07b | 12.35±0.55a |
| 乔灌阶段Shrub-tree stage | 46.05±2.26b | 1.43±0.01b | 0.33±0.04b | 43.90±1.93b | 12.14±1.13a |
| 乔林阶段Tree stage | 58.06±0.65a | 1.58±0.11c | 0.49±0.03a | 62.50±2.12a | 12.3±1.14a |
| 不同小写字母表示凋落物初始养分含量在不同恢复阶段差异显著(P<0.05) | |||||
由图 2可知, 经过1a分解后, 檵木群落不同恢复阶段凋落物N、纤维素总体上呈增加趋势。初始N含量为乔林阶段显著大于乔灌阶段和灌木阶段, 经过1a分解后, 灌木阶段、乔灌阶段和乔林阶段N含量分别为初始含量的1.24、1.21和1.22倍。初始纤维素含量差别不大, 经过1a分解后, 灌木阶段、乔灌阶段和乔林阶段纤维素含量分别为初始含量的1.37、1.57和1.40倍。凋落物初始C含量在乔林阶段最大, 乔灌阶段最小, 且差异显著(P<0.05), 经过1a分解后, 灌木阶段、乔灌阶段和乔林阶段C含量分别为初始含量的1.55、1.31和0.84倍, 灌木、乔灌阶段总体上呈增加趋势, 乔林阶段呈减少趋势。凋落物P初始值为灌木阶段最大, 乔灌阶段最小, 且差异显著(P<0.05), 经过1a分解后, 灌木、乔林阶段P含量较初始含量有所减少, 乔灌阶段有所增加, 灌木阶段、乔灌阶段和乔林阶段P含量分别为初始含量的0.72、1.01和0.80倍。木质素初始含量在乔林阶段最大, 灌木阶段最小, 且差异显著(P<0.05), 经过1a分解后, 乔灌、乔林阶段木质素含量较初始含量有所减少, 灌木阶段有所增加, 灌木阶段、乔灌阶段和乔林阶段木质素含量分别为初始含量的1.18、0.86和0.34倍。

|
| 图 2 檵木群落不同恢复阶段凋落物分解养分残留量动态 Fig. 2 Dynamic of nutrient remaining ratios in different restoration stages of Loropetalum chinense communities |
檵木群落不同恢复阶段凋落物基质质量动态变化与分解速率相关分析表明(表 4), 灌木阶段凋落物分解速率与C、N、P、C/P、N/P呈显著或极显著相关性, 乔灌阶段凋落物分解速率与N、木质素/N呈显著或极显著相关性, 乔林阶段凋落物分解速率与N、纤维素、C/N、木质素/N呈显著或极显著相关性。
| 项目 | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 |
| Y1 | 0.641** | 0.814** | -0.659** | -0.099 | 0.498 | 0.379 | 0.580* | 0.591* | -0.328 | 0.408 | 0.428 | 0.474 |
| Y2 | 0.401 | 0.852** | -0.175 | -0.411 | 0.491 | 0.168 | 0.343 | 0.341 | -0.560* | -0.292 | 0.434 | 0.363 |
| Y3 | 0.019 | 0.743** | -0.339 | -0.757** | 0.538* | -0.348 | 0.230 | 0.403 | -0.814** | -0.486 | 0.364 | 0.371 |
| **P < 0.01, *P < 0.05; X1:有机碳organic C; X2:全N total N; X3:全P total P; X4:木质素lignin; X5:纤维素cellulose; X6: C/N; X7: C/P; X8: N/P; X9:木质素/N lignin/N; X10:木质素/P lignin/P; X11:纤维素/N cellulose/N; X12:纤维素/P cellulose/P; Y1:灌木阶段分解速率litter decomposition rate in shrubs stage; Y2:乔灌阶段分解速率litter decomposition rate in Shrub-tree stage; Y3:乔林阶段分解速率litter decomposition rate in tree stage | ||||||||||||
在众多生态系统中, 凋落物分解每年释放的养分可以满足林木生长需要量的69%—87%, 凋落物分解速率的高低在很大程度上决定了土壤养分的供应能力和生态系统生产力的大小[9]。檵木群落3个不同恢复阶段凋落物年分解系数k值大小顺序为:灌木阶段>乔灌阶段>乔林阶段, 分解系数k值越大, 凋落物分解速度越快[22], 表明本研究中檵木群落灌木阶段凋落物分解相对较快, 乔林阶段较慢。檵木群落不同恢复阶段凋落物分解的基质质量及化学计量对各阶段分解速率影响不同。有研究表明, 凋落物分解速率与N, P含量紧密相关, 一般认为, N, P含量越高, 分解越快[23]。同时, 凋落物基质质量对凋落物分解速率的影响与物种种类和环境条件有很大关系[24]。本研究中N初始含量随正向演替进行逐渐增大, 檵木群落3个不同恢复阶段凋落物N含量总体上升, 本研究中3个不同恢复阶段分解速率均与N呈极显著正相关, 这与宋新章等[9]研究结果相似, 这可能是因为外源氮在凋落物上被微生物固持[25], N含量升高会加速凋落物分解[26]。灌木阶段与P紧密相关, 灌木阶段分解前和分解1a后P含量均较高, 这可能是导致灌木阶段分解系数k值较大的原因之一, 因而灌木阶段凋落物分解速率较其他阶段快。研究表明, 凋落物中C含量越低, 分解速率越快[27]。在凋落物分解1a后, 乔林阶段C含量最低, 灌木阶段最高, 此时乔林阶段分解速率较其他阶段快, 这与一般规律相符合。木质素在凋落物分解过程中属于难分解成分, 纤维素较木质素易分解[28], 在木质素含量高(>25%)的凋落物中, 木质素是预测凋落物分解的较好指标[29]。本研究中3个不同恢复阶段的木质素初始含量均高于目前其他研究报道中的含量(5%—30%)[30], 灌木阶段初始木质素含量最低, 在分解初始时期其凋落物分解最快, 乔林阶段初始木质素含量最高, 在分解初始时期其凋落物分解最慢。随着凋落物分解的进行, 灌木阶段木质素含量逐渐累积并高于乔灌、乔林阶段的木质素含量;乔林阶段含量最低, 因而分解后期乔林阶段分解速率较灌木阶段快, 这与李雪峰等[31]研究一致, 这也证实了木质素含量高限制了凋落物的分解。张艳等[32]对樟(Cinnamomum camphora)和红椿(Toona ciliata)两种凋落叶的研究表明, 随着分解时间的延长, 纤维素含量降低, 而这与本研究结果相反。本研究中3个不同恢复阶段的纤维素总体上升, 这可能是与木质素酶和纤维素酶活性有关[32]。
除凋落物基质质量外, 化学计量值也是预测凋落物分解速率的重要指标[33-34]。大量研究表明, C/N, C/P值越小, 凋落物分解速度越快[24, 27, 35]。在本研究中, 灌木阶段C/N、C/P值总体上较其他恢复阶段高, 其分解速率较其他恢复阶段快, 这可能是因为随着植被的演替, 乔林阶段伴生有相对较多种革质叶片的乔木树种, 如:刺叶冬青(Ilex bioritsensis)、石山樟(Cinnamomum saxatile)等[17], 革质叶片树种角质层发达, 木质素、纤维素、单宁等难降解物质含量较多, 不利于分解[36], 这些因素可能是导致乔林阶段凋落物分解速率较灌木阶段和乔灌阶段慢, 因而乔林阶段分解系数k值较其他阶段小, 也可能与喀斯特森林对N、P元素利用效率有关[15], 这也说明随着檵木群落正向演替的进行, 凋落物分解速率降低。N/P可作为N或P含量是否限制分解的指标[24], N/P较高时分解较慢, N/P较低时凋落物分解较快[37]。Townsend等[38]认为叶片N/P比值<16主要受N的影响。本研究各恢复阶段N/P在分解过程中比值均小于16, 系统受N限制, 在分解过程中, 灌木阶段N/P较低, 这也可能是灌木阶段凋落物分解较快的原因。木质素/N也是最能反映凋落物分解速率的指标之一[39], 3个不同恢复阶段木质素/N值总体下降, 其值随着植被恢复正向演替而逐渐减小, 可见, 植被恢复对凋落物基质质量产生了明显的影响。
气候、土壤环境等也是影响凋落物分解的重要因素。本研究表明, 在1a的分解期内, 檵木群落3个不同恢复阶均在12—3月份(冬春季)凋落物分解最慢, 在6—9月份(夏秋季)分解相对较快, 这与许多研究结果基本一致[40-41]。研究区进行实验年份的12—3月份, 为冬春交替季节, 此时降雨少, 天气寒冷, 这种气象条件可能使微生物活性代谢较弱和淋溶作用减弱[22], 从而导致凋落物分解较慢。灌木阶段和乔灌阶段在6月份凋落物分解均最快, 乔林阶段在9月份凋落物分解最快。这可能是由于此时雨水较多, 气温逐渐升高, 微生物活性代谢及淋溶作用增强[22], 导致凋落物分解相对较快。本研究中, 灌木阶段凋落物剩余率最小, 乔林阶段最大, 这可能是随着檵木群落的恢复, 乔林阶段群落郁闭度逐渐增大[17], 一定程度是影响了乔林阶段的分解速率, 郁闭度是影响凋落物分解速率的主要因子[42]。徐涵湄等[43]研究表明, 受冰雪灾害的毛竹林分解速率与郁闭度呈显著负相关, 竹林郁闭度随雪灾程度增加而减小, 分解速率也随之加快。森林郁闭度不同, 林窗面积不同, 林内的水热条件微环境也随之不同[32], 郁闭度大, 林内有对凋落物分解不太有利的小气候条件[44-45]。Zhang[46]等研究表明, 凋落物的年分解速率与林窗内的小气候因子相关, 郁闭度大的群落相对能形成林内“小气候”, 对外界气候条件的变化往往具有一定的时滞效应, 这可能是导致乔林阶段凋落物分解速率达最大值的出现时间靠后的原因。
综上所述, 檵木群落不同恢复阶段凋落物分解速率受多种因素的的影响。凋落物分解过程和影响分解速率的因素之间的相互作用复杂, 可能还有许多因素未考虑在内, 如土壤动物、土壤微生物等, 且本研究的周期也不够长。
| [1] | Hobbie S E. Effects of plant species on nutrient cycling. Trends in ecology & evolution, 1992, 7(10): 336–339. |
| [2] | Solly E F, Schöning I, Boch S, Kandeler E, Marhan S, Michalzik B, Müller J, Zscheischler J, Trumbore S E, Schrumpf M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. plant and soil, 2014, 382(1): 203–218. |
| [3] | 窦荣鹏, 江洪, 余树全, 马元丹, 宋新章, 郭培培, 张小川. 4种亚热带树木凋落叶的分解研究. 浙江林学院学报, 2010, 27(2): 163–169. |
| [4] | 孙晓辉. 胡桃楸天然次生林凋落物分解的生态学过程. 防护林科技, 2015(5): 9–15. |
| [5] | 陶楚, 杨小波, 万春红, 李东海, 符有利, 高文嵩, 冯丹丹, 杨琦. 海南铜鼓岭保护区2个不同演替阶段森林凋落叶的分解特性. 热带生物学报, 2015, 6(1): 69–77. |
| [6] | 曾锋, 邱治军, 许秀玉. 森林凋落物分解研究进展. 生态环境学报, 2010, 19(1): 239–243. |
| [7] | 王敏英, 刘强. 海南岛中部丘陵地区植被恢复过程中凋落物动态及土壤碳氮含量变化. 广西植物, 2012, 32(5): 630–636. |
| [8] | 陈金玲, 金光泽, 赵凤霞. 小兴安岭典型阔叶红松林不同演替阶段凋落物分解及养分变化. 应用生态学报, 2010, 21(9): 2209–2216. |
| [9] | 宋新章, 江洪, 余树全, 马元丹, 周国模, 窦荣鹏, 郭培培. 中亚热带森林群落不同演替阶段优势种凋落物分解试验. 应用生态学报, 2009, 20(3): 537–542. |
| [10] | 张庆费, 宋永昌, 吴化前, 由文辉. 浙江天童常绿阔叶林演替过程凋落物数量及分解动态. 植物生态学报, 1999, 23(3): 250–255. |
| [11] | 曹向文, 赵洋毅, 熊好琴, 段旭, 舒树淼. 滇东喀斯特石漠化地区不同植被模式土壤酶活性与有机碳. 东北林业大学学报, 2015, 43(11): 79-83, 97. DOI:10.3969/j.issn.1000-5382.2015.11.016 |
| [12] | 李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义. 山地学报, 2003, 21(2): 129–139. |
| [13] | 朱金兆, 刘建军, 朱清科, 吴钦孝. 森林凋落物层水文生态功能研究. 北京林业大学学报, 2002, 24(5/6): 30–34. |
| [14] | 朱双燕, 王克林, 曾馥平, 曾昭霞, 宋同清. 桂西北喀斯特次生林凋落物养分归还特征. 生态环境学报, 2009, 18(1): 274–279. |
| [15] | 曾昭霞, 王克林, 刘孝利, 曾馥平, 宋同清, 彭晚霞, 张浩, 杜虎. 桂西北喀斯特区原生林与次生林鲜叶和凋落叶化学计量特征. 生态学报, 2016, 36(7): 19071914. |
| [16] | 俞国松, 王世杰, 容丽, 冉景丞. 茂兰喀斯特森林主要演替群落的凋落物动态. 植物生态学报, 2011, 35(10): 1019–1028. |
| [17] | 马姜明, 吴蒙, 占婷婷, 梁士楚. 漓江流域岩溶区檵木群落不同恢复阶段物种组成及多样性变化. 生态环境学报, 2013, 22(1): 66–71. |
| [18] | 马姜明, 占婷婷, 莫祖英, 梁士楚. 漓江流域岩溶区檵木群落不同恢复阶段主要共有种生态位变化. 西北植物学报, 2012, 32(12): 2530–2536. DOI:10.3969/j.issn.1000-4025.2012.12.024 |
| [19] | 彭懿, 陈秋波, 王春燕. 烘箱加热法测定橡胶树不同器官有机碳含量. 热带农业科学, 2009, 29(10): 13–16. DOI:10.3969/j.issn.1009-2196.2009.10.003 |
| [20] | 鲁如坤. 土壤农化分析方法. 北京:中国农业科技出版社, 2000. |
| [21] | 范鹏程, 田静, 黄静美, 累文泉, 邱会东. 花生壳中纤维素和木质素含量的测定方法. 重庆科技学院学报:自然科学版, 2008, 10(5): 64–65. |
| [22] | 张尚炬. 短枝木麻黄凋落小枝分解与单宁和养分含量的关系模型. 福建林学院学报, 2014, 34(2): 120–125. |
| [23] | Martínez A, Monroy S, Pérez J, Larrañaga A, Basaguren A, Molinero J, Pozo J. In-stream litter decomPosition along an altitudinal gradient: does substrate quality matter?. Hydrobiologia, 2016, 766(1): 17–28. DOI:10.1007/s10750-015-2432-9 |
| [24] | 葛晓改, 曾立雄, 肖文发, 黄志霖, 周本智. 三峡库区森林凋落叶化学计量学性状变化及与分解速率的关系. 生态学报, 2015, 35(3): 779–787. |
| [25] | 邓长春, 蒋先敏, 刘洋, 张健, 陈亚梅, 和润莲. 高山林线交错带高山杜鹃的凋落物分解. 生态学报, 2015, 35(6): 176–1778. |
| [26] | 杨新芳, 鲍雪莲, 胡国庆, 邵帅, 周锋, 叶佳舒, 解宏图, 梁超. 大兴安岭不同火烧年限森林凋落物和土壤C、N、P化学计量特征. 应用生态学报, 2016, 27(5): 1359–1367. |
| [27] | 林波, 刘庆, 吴彦, 何海. 森林凋落物研究进展. 生态学杂志, 2004, 23(1): 60–64. |
| [28] | 郭剑芬, 杨玉盛, 陈光水, 林鹏, 谢锦升. 森林凋落物分解研究进展. 林业科学, 2006, 42(4): 93–100. |
| [29] | 潘冬荣, 柳小妮, 申国珍, 谢宗强, 罗璐, 刘蕾. 神农架不同海拔典型森林凋落物的分解特征. 应用生态学报, 2013, 24(12): 3361–3366. |
| [30] | 阿姆森KA. 森林土壤: 性质和作用. 林伯群, 周重光, 译. 北京: 科学出版社, 1984: 33-46. |
| [31] | 李雪峰, 韩士杰, 郭忠玲, 郑兴波, 宋国正, 李考学. 红松阔叶林内凋落物表层与底层红松枝叶的分解动态. 北京林业大学学报, 2006, 28(3): 8–13. |
| [32] | 张艳, 张月枯, 张健, 杨万勤, 邓长春, 李建平, 李勋, 唐仕姗, 张明锦. 马尾松人工林林窗大小对两种凋落叶难降解物质含量的影响. 植物生态学报, 2015, 39(8): 785–796. DOI:10.17521/cjpe.2015.0075 |
| [33] | 许新建, 陈金耀, 俞新妥. 武夷山六种杉木伴生树种落叶养分归还的研究. 建林学院学报, 1995, 15(3): 213–217. |
| [34] | 向元彬, 黄从德, 胡庭兴, 涂利华, 杨利林, 张志, 徐伟. 不同密度巨按人工林凋落物分解过程中基质质量的变化. 西北农林科技大学学报:自然科学版, 2015, 43(4): 65–72. |
| [35] | 林成芳, 杨玉盛, 郭剑芬, 陈光水, 袁一丁. 米槠天然林树木根系分解和化学组分变化. 应用生态学报, 2009, 20(6): 1278–1284. |
| [36] | 王意锟, 方升佐, 曲宏辉, 唐罗忠, 宋浩. 森林凋落物分解的影响因素. 林业科技开发, 2012, 26(1): 5–9. |
| [37] | 潘复静, 张伟, 王克林, 何寻阳, 梁士楚, 韦国富. 典型喀斯特峰丛洼地植被群落凋落物C:N:P生态化学计量特征. 生态学报, 2011, 31(2): 335–343. |
| [38] | Townsend A R, Cleveland C C, Asner G P, Bustamante M M. Controls over foliar N: P ratios in tropical rain forests. Ecology, 2007, 88(1): 107–118. DOI:10.1890/0012-9658(2007)88[107:COFNRI]2.0.CO;2 |
| [39] | 刘勇, 李国雷. 不同林龄油松人工林叶凋落物分解特性. 林业科学研究, 2008, 21(4): 500–505. |
| [40] | 金龙, 吴志祥, 杨川, 管利民, 赖华英. 不同林龄橡胶凋落物叶分解特性与有机碳动态研究. 热带作物学报, 2015, 36(4): 698–705. |
| [41] | 赵春梅, 曹建华, 李晓波, 兰国玉, 彭宗波, 蒋菊生. 橡胶林枯落物分解及其氮素释放规律研究. 热带作物学报, 2012, 33(9): 1535–1539. |
| [42] | 张保刚, 梁慧春, 吴祥云, 任杰. 辽东山地影响胡桃楸天然次生林凋落物分解因子研究. 辽宁农业科学, 2014(4): 19–23. |
| [43] | 徐涵湄, 丁九敏, 刘胜, 卜晓莉, 王邵军, 汪家社, 徐自坤, 阮宏华. 雪灾对武夷山毛竹林凋落物分解动态的影响. 南京林业大学学报:自然科学版, 2010, 34(3): 131–135. |
| [44] | Lin N, Bartsch N, Heinrichs S, Vor T. Long-term effects of canopy opening and liming on leaf litter production, and on leaf litter and fine-root decomposition in a European beech (Fagus sylvatica L.) forest. Forest Ecology and Management, 2015, 338: 183–190. DOI:10.1016/j.foreco.2014.11.029 |
| [45] | Sariyildiz T. Effects of gap-size classes on long-term litter decomposition rates of beech, oak and chestnut species at high elevations in Northeast Turkey. Ecosystems, 2008, 11(6): 841–853. DOI:10.1007/s10021-008-9164-x |
| [46] | Zhang Q S, Liang Y W. Effects of gap size on nutrient release from plant litter decomposition in a natural forest ecosystem. Canadian Journal of Forest Research, 1995, 25(10): 1627–1638. DOI:10.1139/x95-177 |