 2017, Vol. 37
2017, Vol. 37文章信息
- 阿的鲁骥, 字洪标, 刘敏, 陈焱, 杨有芳, 王长庭
- ADE Luji, ZI Hongbiao, LIU Min, CHEN Yan, YANG Youfang, WANG Changting.
- 高寒草甸地下根系生长动态对积雪变化的响应
- Response of belowground root growth dynamics to snow cover change in alpine meadow
- 生态学报. 2017, 37(20): 6773-6784
- Acta Ecologica Sinica. 2017, 37(20): 6773-6784
- http://dx.doi.org/10.5846/stxb201605240995
-
文章历史
- 收稿日期: 2016-05-24
- 网络出版日期: 2017-06-01

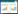




2. 西南民族大学生命科学技术学院, 成都 610041
2. School of Life and Technology, Southwest University for Nationalities, Chengdu 610041, China
根系是植物吸收营养元素和水分的重要器官, 也是陆地生态系统重要的组成部分, 在维持和提高土壤质量方面起着重要作用[1]。植被根系具有丰富的维持生态系统功能的生态多样性, 其地下分布格局和现存量反映了土壤中水分和养分的分配格局, 决定着植被对地下水分和养分的利用效率和潜力[2], 是生态系统碳分配和碳循环的核心环节之一[3]。另一方面, 根系对土壤环境变化十分敏感, 能及时地感知和响应土壤微环境的变化[1]。例如, 土壤温度过低往往会引起根系生长停滞[4];适当增加土壤水分有助于促进植物根系生长[5], 而土壤水分不足则会导致根系伸长受抑、分支减少[6];因此, 研究根系对环境变化的响应, 有助于更深入、全面地了解陆地生态系统对全球变化的响应机制。根系生态学受到了广泛关注, 但由于挖掘法、土块法、土钻法、剖面法等传统方法[7]工作量巨大且对根系具有较大的破坏性, 在很大程度上制约了对根系生长动态的研究[8]。微根窗技术(Minirhizotron mothed)[9]这一非破坏性的研究方法克服了传统研究方法的缺陷, 可在多个时段原位重复观测根系生长动态, 为根系生产力及其周转方面的研究提供了便利。近年来, 随着这一技术的兴起, 根系生态学得到了迅速发展[10]。
青藏高原是影响欧亚大陆乃至全球气候格局的重要因素[11], 同时, 青藏高原生态系统对全球变化十分敏感, 其改变往往早于周围地区, 能更加明显的对全球气候变化做出预警[12]。近年来, 雪生态学(Snow Ecology)[13]迅速发展并逐渐成为与全球变化密切相关的热点领域[12]。作为气候变化影响下的一项主要生态过程[14], 冰雪演变迅速改变了青藏高原地区雪被状况[15]。季节性积雪堆积与消融深刻地影响着青藏高原高寒草甸的群落特征[16]、土壤理化性质[17]及土壤微生物活动[18]。在青藏高原高寒草甸的根系生态学研究方面, 目前主要集中在增温[19]和演替[20]对根系的影响以及冻土区根系分布[21]等, 有关积雪量变化对高寒草甸根系影响的研究则报道较少。本课题组在青藏高原东缘红原县的高寒草甸开展积雪量野外控制试验, 参考前人研究方法[22-23], 在降雪后采用人工堆积方式设置不同积雪量对样方进行处理, 并运用微根窗技术对不同积雪量处理的群落根系生长动态进行连续观测, 旨在解决以下科学问题:(1)积雪变化如何影响高寒草甸植物根系生长动态?(2)造成这一影响的机制是什么?为青藏高原高寒草甸植物群落根系对气候变化的响应与适应机制研究提供基础数据与理论依据。
1 研究方法 1.1 实验区自然环境概况研究区位于四川西北部红原县境内的西南民族大学青藏高原基地, 地理坐标为32°49′ N, 102°34′ E, 海拔3485 m, 属于青藏高原向四川盆地过渡的高原区[24], 其气候类型为大陆性高原气候, 日温差大、霜冻期长, 四季变化不明显, 年均气温1.1℃, 年均相对湿度60%—70%;干湿季节分明, 年均雨量791.95 mm, 主要集中在5—9月, 年均蒸发量达1262.5 mm, 除6—8月外, 其余时间段降水量均小于蒸发量, 空气干燥;日照时间长, 太阳辐射强, 年均日照时间2158.7 h, 太阳辐射年总量为6194 MJ/m2[25];试验区土壤类型为高寒草甸土[26]。
试验区植被每年的生长季由4月开始到9月结束, 返青期主要集中在4月至5月, 在8月中上旬达到生长峰值;植被平均盖度在80%以上, 高度最高达到45—60 cm;植物种类主要包括莎草科的四川嵩草(Kobresia setchwanensis)和高山嵩草(K. pygmaea), 禾本科的四川剪股颖(Agrostis clavata)和垂穗披碱草(Elymus nutans), 杂类草中条叶银莲花(Anemone trullifolia)、钝苞雪莲(Saussurea nigrescens)和鹅绒委陵菜(Potentilla anserine)等[27]。
1.2 试验设计 1.2.1 样地设置和积雪量野外控制试验在试验区内选择地势相对一致、植物分布相对均匀的高寒草甸作为样地。采用随机区组试验, 在30 m×30 m的区域内均匀布设20个2 m×2 m的样方, 样方间至少间隔1.5 m作为缓冲区。
2013年11月至2014年3月期间, 在降雪后开展积雪量野外控制试验, 期间自然积雪量见表 1。共设置4个积雪量处理, 即CK、S1、S2和S3。其中, CK为自然降雪的积雪量, S1、S2和S3的积雪量分别为自然积雪量的2倍、3倍和4倍, 每个处理设置5个重复。具体操作方法如下:(1)在样地周围建立积雪场。在积雪场上均匀铺设2 m×2 m防水布若干, 并用地钉固定;(2)降雪结束后, 拔出地钉, 收集防水布上的积雪分别均匀堆积在S1、S2和S3的样方中。S1、S2和S3每个样方中的堆积量分别为1块、2块和3块防水布上的全部积雪。处理期内, CK、S1、S2、S3样方中积雪覆盖持续时间分别为14、17、19、20 d, 累计积雪量分别为71.4、143.4、214.2、285.6 mm。
| 试验期Experiment period | 2013年 | 2014年 | |||
| 11月Nov. | 12月Dec. | 1月Jan. | 2月Feb. | 3月Mar. | |
| 自然降雪量Monthly snowfall/mm | 12.5 | 1.1 | 4.5 | 17.4 | 36.2 |
在积雪量野外控制试验期间, 用4台智能多点土壤温度记录仪(YM-01A, Handan, China)分别记录每个处理0—10 cm和10—20 cm的土层温度, 测量精度为±0.2 ℃, 温度分辨率为0.01℃。
1.2.3 微根管安装和数据读取2013年9月, 在每处理梯度中随机选取3个样方, 参照Johnson等[28]的方法在每个样方中安装1根微根管(长100 cm, 外径7 cm, 内径6.4 cm), 共计12根。微根管与地面成30°夹角, 露出地面部分约为20 cm, 垂直深度约为60 cm, 采集图像尺寸约为0.04 m2。微根管露出地面部分用配套橡胶盖封口, 并用胶带做防水处理, 再用黑布包裹地面部分以避免光射入影响根系生长。微根管周围用钻出的土填平, 使其与土壤紧密接触的同时尽量做到减小对管子周围的土壤扰动, 在不取数据时用黑色塑料袋包裹以减少管子的热量传导[29]。
于2014年4月27日至9月20日期间用CI-600 Root Scanning System (CID Bio-Science Inc., Camas, WA, USA)采集根系图像数据, 共采集10次, 平均间隔时间为16 d。每根微根管按垂直土层深度分为表层土壤(0—10 cm)和深层土壤(10—20 cm)分别采集图像数据。
1.2.4 根系数据处理方法采用根系分析软件WinRHZIO Tron MF (CID Bio-Science Inc., Camas, WA, USA)对所获图像数据进行处理, 以获取根长、直径、根尖数和表面积等参数, 然后对所获根系信息进行统计分析。
参照吴伊波等[30]的方法计算单位面积根系现存量(RLDa, g/m2), 由RLDa估算出根系现存量(g/m2)[31]。平均根系现存量(g/m2)为10次测定单位面积根系现存量的均值。
根系生产量(g/m2)为首次测定与末次测定根系现存量的差值。
单位体积的根长密度(RLDv, m/m3)按公式(1)计算:
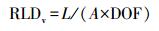
|
(1) |
式中, L为微根窗中观察到的根长(m), A(m2)为观测窗面积(图像大小), DOF(m)为微根管至周围土壤的距离, DOF值一般在0.002—0.003 m[32]。由于高寒草甸根系直径较小, 本研究在计算中DOF取0.003 m[30];通过比根长(SRL, m/g)将RLDv转化为单位体积的现存量(RBD, g/m3):
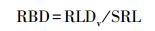
|
(2) |
式中, SRL为土钻法所得每克根(<1 mm)现存量的根长(m/g)。
以上单位体积根长密度通过乘以取样土壤剖面深度(D)转换成以单位面积为基础的现存量:
|
(3) |
根系净生产速率(RLDNGR)的计算方法参照王孟本等[33]的方法稍加改动:
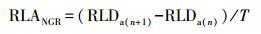
|
(4) |
式中, RLDNGR为根系净生产速率(g m-2 d-1), RLDa(n+ 1)和RLDa (n)分别为第n + 1与第n次观测中的单位面积根系现存量; T为相邻2次观测的间隔天数。RLDNGR为正值说明根系的生长速率大于死亡速率(正生长), RLDNGR为0表示根系的生长速率与死亡速率相等(零生长), RLDNGR为负值表示根系的死亡速率大于生长速率(负生长)[33]。
1.3 统计与分析采用SPSS 20.0对植物根系表面积、根尖数量、根系现存量、平均根系现存量以及根系生产量进行单因素方差分析(One-way analysis of variance, One-way ANOVA), 最小显著差数法(LSD)进行多重比较, 并对土壤环境因素(温度)与根系生长状况(根系表面积、根尖数量以及根系现存量)相关性分析(Pearson);采用Microsoft Office制作图表;应用Fuzzy数学隶属函数对不同积雪量处理的根系生长状况(根系表面积、根尖数量以及根系现存量)进行综合分析[34]。
2 结果与分析 2.1 积雪量对高寒草甸根系生长的影响 2.1.1 积雪量对根系表面积的影响在表层土壤中, 根系表面积在4月27日至8月5日均匀地随时间而逐步增加, 而在8月5日至观测结束(9月20日)呈“W”型变化(图 1)。CK与S3的根系表面积在8月5日达到峰值, 分别为(68.58±5.22) cm2和(48.20±9.99) cm2, 而S1与S2的根系表面积的峰值出现在9月5日, 分别为(72.93±9.37) cm2与(70.69±9.67) cm2。显著性检验表明, CK、S1和S2处理的根系表面积峰值间无显著差异, 而S3处理的根系表面积峰值则与其余处理间均表现出显著差异性(P<0.05)。
由图 1可见, 在深层土壤中, CK的根系表面积在4月27日至8月5日期间始终呈现增加的趋势, 在8月5日达到峰值[(31.98±10.24)cm2], 随后便逐步下降;S1在7月20日前呈现逐步增加趋势, 而7月20日至9月20日则表现为“M”型, 其峰值出现于9月5日[(69.32±8.29)cm2];S2的根系表面积峰值出现在9月5日[(38.84±4.79)cm2], 这一时间点之前呈现逐步增加趋势, 而之后则呈下降趋势;S3的根系表面积的变化规律整体上与S2处理类似, 峰值出现于8月19日[(25.52±8.10)cm2]。值得注意的是, S1与S2的根系表面积的增长速率显著高于对照组。

|
| 图 1 积雪量对高寒草甸植被根系表面积动态的影响 Fig. 1 Effects of snowpack volume on the dynamic of root surface area in Alpine meadow 不同字母表示在0.05水平上差异显著, CK为自然降雪的积雪量, S1、S2和S3的积雪量分别为自然积雪量的2倍、3倍和4倍 |
综上可见, 根系表面积整体上随着积雪量增加呈现先增加后降低的趋势。S1处理下根系表面积总体上大于其余处理, CK与S2处理相当, 而S3处理下根系表面积最小。这一趋势在深层土壤中表现的更为明显, 其中, S1处理下根系表面积显著(P<0.05)高于其他处理。
2.1.2 积雪量对根尖数量的影响图 2反映了不同积雪量处理下高寒草甸植被根系根尖数量的动态变化。表层土壤中, CK、S1与S3的根尖数量峰值均出现在8月5日, 分别为(536.50±76.01)根、(547.33±68.82)根和(317.33±58.09)根。而S2根尖数量峰值出现于9月5日(535.00±72.50)根。S3的根尖数量峰值均显著低于其他处理(P<0.05);深层土壤中, 除CK根尖数量峰值出现在8月5日[(243.00±56.31)根]外, 其余各处理根尖数量峰值均出现在9月5日, S1、S2和S3分别为(381.00±48.54)根、(306.00±50.15)根和(173.00±29.50)根。在不同积雪量作用下, 根尖数量的变化规律整体上与根系表面积的变化规律相类似, 即随着积雪量的增加表现为先增加后减少。其中, S1处理根尖数量最高, 而S3处理根尖数量最低。

|
| 图 2 积雪量对高寒草甸植被根尖数量动态的影响 Fig. 2 Effects of snowpack volume on the dynamic of root tips in Alpine meadow |
图 3反映了不同处理下高寒草甸根系现存量的动态变化过程。表层土壤中, CK、S1、S3处理的根系现存量峰值均出现于8月5日, 分别为(558.33±50.80)g/m2、(617.47±42.75)g/m2和(413.33±39.41)g/m2, 而S2处理根系现存量峰值则出现于9月5日((612.62±47.80)g/m2);深层土壤中, CK的根系现存量峰值出现在8月5日((272.71±69.75)g/m2), S1处理的根系现存量峰值出现在8月19日((373.77±41.96)g/m2), S2与S3处理下根系现存量的峰值出现在9月5日, 分别为(334.53±21.11)g/m2和(210.74±20.82)g/m2。

|
| 图 3 积雪量对高寒草甸植被根系现存量动态的影响 Fig. 3 Effects of snowpack volume on the dynamic of root standing crop in Alpine meadow |
对平均根系现存量(图 4)的研究表明, 在表层土壤中, 4个处理平均根系现存量由高到低依次为S1、S2、CK和S3, 各处理间差异显著(P<0.05);而深层土壤中, 则表现为S1>CK>S2>S3, 各处理间差异显著(P<0.05)。

|
| 图 4 积雪量对高寒草甸植被根系平均现存量的影响 Fig. 4 Effects of snowpack volume on the mean root standing crop in Alpine meadow |
不同积雪量处理不同程度地影响了高寒草甸根系净生长速率动态(表 2)。
| 土层/cm Depth |
处理 Treatment |
净生产速率RLDNGR/ (g m-2 d-1) | ||||||||
| 5月14日 May 14 |
5月31日 May 31 |
6月19日 Jun 19 |
7月4日 Jul 4 |
7月20日 Jul 20 |
8月5日 Aug 5 |
8月19日 Aug 19 |
9月5日 Sep 5 |
9月20日 Sep 20 |
||
| 0—10 | CK | 0.69±0.01b | 4.01±1.25b | 3.00±1.39a | 3.23±1.27ab | 4.00±1.18a | 3.62±0.4b | -1.64±0.68b | 1.01±0.46a | -3.93±0.22c |
| S1 | 1.87±0.18a | 9.73±0.38a | 3.01±0.23a | 0.66±0.31b | 0.19±0.21b | 4.58±0.25a | -1.63±0.17b | 1.15±0.22a | -2.31±0.1b | |
| S2 | -0.42±0.69b | 2.25±0.25c | 1.25±0.06a | 5.52±0.12a | 3.55±0.33a | 3.69±0.14b | -0.12±0.13a | 1.68±0.26a | -4.9±0.23d | |
| S3 | 0.93±1.18ab | 3.85±0.95bc | 2.06±1.38a | 1.96±3.27b | 3.32±0.88a | 4.3±0.2b | -4.08±0.82c | 0.81±1.12a | -1.27±0.61a | |
| 10—20 | CK | 0.64±0.14b | 1.52±0.76b | 1.31±1a | 1.5±0.7c | 0.89±2.47ab | 1.17±1.79a | -1.05±0.08c | 0.55±0.7b | -1.83±1.35b |
| S1 | 1.99±0.71a | 0.95±0.22a | 1.00±0.16a | 3.93±0.46a | 2.11±0.51b | 0.68±0.56a | 0.39±0.25d | -1.35±0.54a | -1.66±0.37c | |
| S2 | 0.83±0.15b | 0.35±0.19c | 0.58±0.08a | 2.92±0.49b | 3.24±0.11a | 3.14±0.17a | 2.81±0.43a | 0.82±0.77b | -2.52±0.07b | |
| S3 | 0.94±0.42b | 1.26±0.81bc | 0.68±0.58a | -0.1±0.88d | 1.3±0.34ab | 1.5±0.49a | 1.19±0.14b | 0.86±0.22b | -1.14±0.14a | |
| 不同字母表示在0.05水平上差异显著, CK为自然降雪的积雪量, S1、S2和S3的积雪量分别为自然积雪量的2倍、3倍和4倍 | ||||||||||
在表层土壤中, CK在5月14日至8月5日期间根系保持相对平稳的正生长(3.00—4.01 g m-2 d-1)。S1处理下, 根系于5月14日至5月31日、7月20日至8月5日这两段时间内出现正生长高峰, 其净生长速率分别为9.73 g m-2 d-1与4.58 g m-2 d-1。S2处理的根系净生长速率相对平稳, 高峰期出现于6月19日至7月4日(5.52 g m-2 d-1)。S3的植被净生长速率变化规律与S1类似, 在同一时期出现了两个正生长的高峰, 但其净生长速率则低于S1处理, 分别为3.85 g m-2 d-1与4.30 g m-2 d-1。在表层土壤中, 8月5日之后, 所有处理均出现了正负生长交替出现, 呈负生长占主导地位的趋势。
在深土壤中, CK的根系生长状态相对稳定, 未出现明显波动;S1的根系在4月27日至5月14日、6月19日至7月20日这两段时间内维持了相对较高的正生长;S2处理的根系生长的高峰期主要出现在6月4日至8月19日;S3处理下根系净生长速率始终维持在一个相对较低的水平。
2.1.5 积雪量对根系生产量的影响根系生产量反应植物群落根系在整个生长季的生长情况。从图 5可见, 表层土壤的根系生产量随着积雪量的增加而呈现先增加后降低的趋势, 根系生产量从高到低依次为S1、CK、S3和S2。除S2、S3无显著差异外, 其余各处理间均存在显著差异(P<0.05);深层土壤中, 根系生产量随着积雪量增加呈现先增加后降低的趋势更加显著, 从高到低依次为S1、S2、S3、CK, 各处理间根系生产量均具有显著差异(P<0.05)。

|
| 图 5 积雪量对高寒草甸植被根系生产量的影响 Fig. 5 Effects of snowpack volume on the root production in Alpine meadow |
应用模糊数学中的隶属函数值法对不同积雪量对植物根系影响程度进行了综合评价(表 3), 结果表明, 表层土壤(0—10 cm)中, 积雪量对植物根系影响的程度由高到低依次为S2 (0.897)、S1 (0.881)、CK(0.562) 和S3(0.281);下层土壤(10—20 cm)中, 积雪量对植物根系影响的程度由高到低依次为S1(0.737)、S2(0.468)、CK(0.162) 与S3(0.058)。综合上下土层的评判结果发现, 不同积雪量对高寒草甸植物根系影响程度表现为S1>S2>CK>S3。
| 土层深度Soil depth/cm | 处理Treatment | 根系表面积Root surface area | 根尖数量 Root tips |
根系现存量 Root standing crop |
综合评判 Synthetic evaluation |
排位 Ranking |
| 0—10 | CK | 0.820 | 0.868 | 0 | 0.562 | 3 |
| S1 | 1 | 1 | 0.644 | 0.881 | 2 | |
| S2 | 0.840 | 0.852 | 1 | 0.897 | 1 | |
| S3 | 0 | 0 | 0.843 | 0.281 | 4 | |
| 10—20 | CK | 0.245 | 0.242 | 0 | 0.162 | 3 |
| S1 | 1 | 1 | 0.212 | 0.737 | 1 | |
| S2 | 0.179 | 0.225 | 1.000 | 0.468 | 2 | |
| S3 | 0 | 0 | 0.175 | 0.058 | 4 |
积雪量变化显著的影响了不同土层的土壤温度(图 6)。2013年11月至2014年4月期间, 表层土壤(0—10 cm)的土壤温度整体上表现为随着积雪量的增加而降低。其中, 2013年11月、12月、2014年1月、4月不同处理间土壤温度差异显著(P<0.05)。随着大气温度逐渐回暖, 各处理间表层土壤温度差异逐渐减小;在深层土壤(10—20 cm)中, 2013年11月各处理间土壤温度无显著差异。随后的12月、1月及2月土壤温度随着积雪量的增加而呈现先降后增的趋势, 部分处理间存在显著差异(P<0.05)。3月至6月, 土壤温度随着积雪量的增加而降低, 其中, 3月、4月及6月部分处理间土壤温度差异显著(P<0.05)。7月各处理间土壤温度无显著差异。

|
| 图 6 积雪量对高寒草甸土壤温度动态的影响 Fig. 6 Effects of snowpack volume on the dynamic of soil temperature in Alpine meadow |
相关性分析表明(表 4), 在表层土壤(0—10 cm)中, 土壤温度与根系现存量、表面积、根尖数量间均有显著相关(P<0.05);在下层土壤(10—20 cm)中, 土壤温度与根系生长状况间表现为正相关关系, 但未达到显著水平。
| 根系参数 Root parameters |
与土壤温度的相关系数Pearson correlation index | |
| 0—10 cm土层Soil depth 0—10 cm | 10—20cm土层Soil depth 10—20 cm | |
| 根系现存量Root standing crop | 0.594* | 0.212 |
| 根系表面积Root surface area | 0.612* | 0.245 |
| 根尖数量Root tips | 0.687* | 0.237 |
| *表示在0.05水平显著相关(双尾) | ||
由于积雪是热的不良导体, 具有较低的导热性与较大的热容量[35], 可通过影响能量平衡、大气循环、土壤水分蒸发等过程, 对土壤温度产生一定的影响[36-38]。积雪覆盖深度、雪层密度以及积雪覆盖时间均会强烈地影响冻结期土壤的冻结速率和融化期土壤升温状况[39], 积雪覆盖在冬季可防止土壤热量的散失, 使土壤温度高于大气温度; 春季温度回升时则阻止土壤温度升高, 使回升时间滞后[40]。在本研究中, 通过实际观测发现, 尽管大气温度低于0℃, 但青藏高原较高的热辐射水平[41]依旧使得积雪迅速融化, 导致研究区内积雪覆盖时间缩短, 积雪对地面的保温作用大幅降低。积雪融化后渗入地下迅速冻结形成地下冰, 这些地下冰能够有效阻止地表水和土壤水分的下渗[42], 使得地下冰在土壤表层聚集。与此同时, 由于青藏高原昼间较高的热辐射水平以及巨大的昼夜温差[43], 导致地下冰的反复冻融, 在该过程中地下冰层逐步下移, 地下冰储量的不同也改变了反复冻融过程的持续时间及冰层下移的速率。例如, 研究发现2013年11月到2014年4月间, 0—10 cm土层温度随着积雪量的增加而降低, 但在10—20 cm土层中, 同样的变化规律则延迟到2014年3—6月发生。而当地下冰逐渐消失后, 植被状况[44]是决定不同处理间土壤温度变化的主要因子。
作为陆地生态系统重要组成部分的根系, 其生长与根际环境密切相关。土壤水分与温度直接控制着根系的生长与发育[45-46]。在一定范围内, 温度升高有利于根系的生长发育[47], 而低温胁迫则导致根系生长受阻[48]甚至受损[49]。本研究结果表明, 积雪量变化对生长季节的土壤温度产生了显著影响, 随着积雪量增加, 土壤温度逐渐降低。同时, 不同积雪量也对根系的生长状况产生了较大的影响, 积雪量达到143.4 mm时(S1), 根系生长状况最佳;积雪量达到214.2 mm(S2)时, 根系生长期延迟, 但最终的生长状况仍略优于对照(CK); 积雪量达到285.6 mm(S3)时, 地下根系的生长受到抑制。相关性分析表明, 在不同土层中, 根系生长均与土壤温度呈正相关关系, 这与Pregitzer等[50]的研究结果相似。一些研究认为, 地下冰融化和积雪融化产生的水分有利于返青[48]。但是, 过高的土壤含冰量和低地温条件对植物根系生长十分不利, 冻融对土壤结构的破坏会增加植物须根的死亡[51]。生长季初期(4—5月)草甸植物对水分需求量相对较少, 但对温度要求较高, 因此, 地下冰融化和积雪融化对土壤温度回升的负面影响必然间接地影响到植物及其根系的生长。同时, 青藏高原特殊的气候条件引起的冻土层反复冻融过程中, 融化时显著的淋溶作用和冻结时强烈的物理破坏作用以及对土壤微生物的间接作用, 都会影响到根系的生长和存活[52-53]。因此, 积雪量适当增加, 能满足植物生长初期对水分的需求, 有利于植物根系的生长。但积雪量增加过多, 会引起地下冰储量增加, 进而加重土壤冻融过程引起的负面效应, 导致土壤温度过低, 从而抑制了根系的生长。
综上所述, 地下冰储量、土壤温度和土壤水分等多个环境因子共同作用决定了高寒草甸植物根系生长状况, 而这些因子间往往又存在着或协同或拮抗的关系, 例如:地下冰储量的增加在降低土壤温度的同时增加土壤含水量。冬季不同积雪量对环境因子造成了不同的影响, 改变了不同时期其土壤环境, 影响了高寒草甸植物根系的生长状况。在某一时间段内, 多种环境因子共同作用为根系的生长营造了一个有利环境, 我们称其为“促进期”, 反之则为“抑制期”, “促进期”越长(“抑制期”越短), 可能越有利于根系生长。随着积雪量的逐渐增加, “促进期”逐渐延长, 当积雪量达到143.4 mm(S1)时, “促进期”可能占据主导, 此时根系生长状况也最佳;而随着积雪量的进一步增加, “促进期”逐渐缩短, 土壤环境逐步不利于根系生长, 最终当积雪量达到285.6 mm(S3)时, “抑制期”可能占主导作用, 根系生长受到抑制。
4 结论研究结果表明, 冬季积雪量的改变使土壤温度发生变化, 进而影响高寒草甸植物群落根系的生长动态。随着积雪量的增加, 0—10 cm土层土壤温度逐渐降低;同样的变化规律也出现在了10—20 cm土层, 但时间上有所延迟。同时, 积雪量的改变对高寒草甸植物群落根系的生长状况造成了影响, 当冬季积雪量达到143.4 mm, 最有利于根系生长, 根系表面积、根尖数量、根系现存量以及根系生产量达到最大值, 同时根系生长旺盛期也有所提前和延长;随着积雪量的进一步增加, 积雪对根系生长的正影响逐渐降低, 生长旺期逐渐推迟甚至消失。相关性分析表明, 在不同土层中, 根系生长均与土壤温度呈正相关关系。这些研究结果说明, 冬季降雪在一定程度上控制着高寒草甸生长季节的土壤温度, 从而影响了高寒草甸植被群落根系的生长发育, 最终影响到高寒草甸生态系统中碳分配与碳循环过程。但积雪量的增加不仅会影响土壤温度, 同时还可能改变土壤水分以及土壤结构, 而土壤水分与土壤结构变化是否是造成根系生长动态变化的原因则还需要进一步的研究证实。
致谢:感谢西南民族大学青藏高原生态保护与畜牧业高科技研究示范基地提供试验样地。
| [1] | 梁建生, 张建华, 曹显祖. 根系环境温度变化对根系吸水和叶片蒸腾的影响. 植物学报, 1998, 40(12): 1152–1158. DOI:10.3321/j.issn:1672-9072.1998.12.011 |
| [2] | Schenk H J. Vertical vegetation structure below ground: scaling from root to globe//Esser K, Lüttge U, Beyschlag W, Murata J, eds. Progress in Botany. Berlin Heidelberg: Springer, 2005, 66: 341-373. |
| [3] | 郝艳茹, 彭少麟. 根系及其主要影响因子在森林演替过程中的变化. 生态环境学报, 2005, 14(5): 762–767. |
| [4] | 程建峰, 陈素珍, 潘晓云, 陈凤梅, 方加海, 刘宜柏. 土壤温度对陆稻根系生长发育的影响. 江西农业大学学报, 2000, 22(1): 6–10. |
| [5] | 杨永辉, 武继承, 吴普特, 黄占斌, 赵西宁, 管秀娟, 何方. 保水剂用量对小麦不同生育期根系生理特性的影响. 应用生态学报, 2011, 22(1): 73–78. |
| [6] | 王秋菊, 李明贤, 赵宏亮, 迟力勇. 控水灌溉对水稻根系生长影响的试验研究. 中国农学通报, 2008, 24(8): 206–208. |
| [7] | 黄瑞冬. 植物根系研究方法的发展. 沈阳农业大学学报, 1991, 22(2): 164–168. |
| [8] | Matamala R, Gonzàlez-Meler M A, Jastrow J D, Norby R J, Schlesinger W H. Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science, 2003, 302(5649): 1385–1387. DOI:10.1126/science.1089543 |
| [9] | Cheng W X, Coleman D C, Box J E J. Root dynamics, production and distribution in agroecosystems on the Georgia piedmont using minirhizotrons. Journal of Applied Ecology, 1990, 27(2): 592–604. DOI:10.2307/2404304 |
| [10] | 白文明, 程维信, 李凌浩. 微根窗技术及其在植物根系研究中的应用. 生态学报, 2005, 25(11): 3076–3081. DOI:10.3321/j.issn:1000-0933.2005.11.039 |
| [11] | Kutzbach J E, Prell W L, Ruddiman W F. Sensitivity of Eurasian climate to surface uplift of the Tibetan Plateau. The Journal of Geology, 1993, 101(2): 177–190. DOI:10.1086/648215 |
| [12] | Zhang Y Q, Welker J M. Tibetan alpine tundra responses to simulated changes in climate: aboveground biomass and community responses. Arctic and Alpine Research, 1996, 28(2): 203–209. DOI:10.2307/1551761 |
| [13] | Madan N J. Snow ecology: an interdisciplinary examination of snow-covered ecosystems. Journal of Ecology, 2001, 89(6): 1097–1098. DOI:10.1111/jec.2001.89.issue-6 |
| [14] | 李培基. 青藏高原积雪对全球变暖的响应. 地理学报, 1996, 51(3): 260–265. |
| [15] | Oechel W C, Callaghan T V, Gilmanov T G, Holten J I, Maxwell B, Molau U, Sveinbjörnsson B. Global Change and Arctic Terrestrial Ecosystems. New York: Springer, 1997. |
| [16] | 胡霞, 吴宁, 尹鹏, 吴彦. 川西高原季节性雪被覆盖下凋落物输入对土壤微生物数量及生物量的影响. 生态科学, 2013, 32(3): 359–364. |
| [17] | Liu L, Wu Y, Wu N, Xu J J, Mao Y, Luo P, Zhang L. Effects of freezing and freeze-thaw cycles on soil microbial biomass and nutrient dynamics under different snow gradients in an alpine meadow (Tibetan Plateau). Polish Journal of Ecology, 2010, 58(4): 717–728. |
| [18] | Schimel J P, Mikan C. Changing microbial substrate use in Arctic tundra soils through a freeze-thaw cycle. Soil Biology and Biochemistry, 2005, 37(8): 1411–1418. DOI:10.1016/j.soilbio.2004.12.011 |
| [19] | 余欣超, 姚步青, 周华坤, 金艳霞, 杨月娟, 王文颖, 董世魁, 赵新全. 青藏高原两种高寒草甸地下生物量及其碳分配对长期增温的响应差异. 科学通报, 2015, 60(4): 379–388. |
| [20] | 张燕堃, 张灵菲, 张新中, 冶娜娜, 江小雷. 不同草地恢复措施对高寒草甸植物根系特征的影响. 兰州大学学报:自然科学版, 2014, 50(1): 107–111. |
| [21] | 岳广阳, 赵林, 王志伟, 邹德富, 张乐乐, 乔永平, 赵拥华, 牛丽. 多年冻土区高寒草甸根系分布与活动层温度变化特征的关系. 冰川冻土, 2015, 37(5): 1381–1387. |
| [22] | Starr G, Oberbauer S F, Ahlquist L E. The photosynthetic response of Alaskan tundra plants to increased season length and soil warming. Arctic, Antarctic, and Alpine Research, 2008, 40(1): 181–191. DOI:10.1657/1523-0430(06-015)[STARR]2.0.CO;2 |
| [23] | Wipf S, Stoeckli V, Bebi P. Winter climate change in alpine tundra: plant responses to changes in snow depth and snowmelt timing. Climatic Change, 2009, 94(1/2): 105–121. |
| [24] | 徐洪灵, 张宏, 张伟. 川西北高寒草甸土壤呼吸速率日变化及温度影响因子比较. 四川师范大学学报:自然科学版, 2012, 35(3): 405–411. |
| [25] | 高永恒, 陈槐, 罗鹏, 吴宁, 王根绪. 放牧强度对川西北高寒草甸植物生物量及其分配的影响. 生态与农村环境学报, 2008, 24(3): 26–32. |
| [26] | Gao Y H, Luo P, Wu N, Chen H, Wang G X. Grazing intensity impacts on carbon sequestration in an alpine meadow on the eastern tibetan plateau. Research Journal of Agriculture and Biological Sciences, 2007, 3(6): 642–647. |
| [27] | Li G Y, Liu Y Z, Frelich L E, Sun S C. Experimental warming induces degradation of a Tibetan alpine meadow through trophic interactions. Journal of Applied Ecology, 2011, 48(3): 659–667. DOI:10.1111/j.1365-2664.2011.01965.x |
| [28] | Johnson M G, Tingey D T, Phillips D L, Storm M J. Advancing fine root research with minirhizotrons. Environmental and Experimental Botany, 2001, 45(3): 263–289. DOI:10.1016/S0098-8472(01)00077-6 |
| [29] | 王长庭, 王启兰, 景增春, 冯秉福, 杜岩功, 龙瑞军, 曹广民. 不同放牧梯度下高寒小嵩草草甸植被根系和土壤理化特征的变. 草业学报, 2008, 17(5): 9–15. DOI:10.11686/cyxb20080502 |
| [30] | 吴伊波, 车荣晓, 马双, 邓永翠, 朱敏健, 崔骁勇. 高寒草甸植被细根生产和周转的比较研究. 生态学报, 2014, 34(13): 3529–3537. |
| [31] | Norby R J, Ledford J, Reilly C D, Miller N E, O'Neill E G, Schlesinger W H. Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(26): 9689–9693. DOI:10.1073/pnas.0403491101 |
| [32] | Sanders J L, Brown D A. A new fiber optic technique for measuring root growth of soybeans under field conditions. Agronomy Journal, 1978, 70(6): 1073–1076. DOI:10.2134/agronj1978.00021962007000060043x |
| [33] | 王孟本, 荀俊杰, 陈建文, 史建伟. 晋西北黄土区幼龄柠条细根的净生长速率. 生态学报, 2010, 30(5): 1117–1124. |
| [34] | 苏秀红, 宋小玲, 强胜, 段惠. 不同地理种群紫茎泽兰种子萌发对干旱胁迫的响应. 应用与环境生物学报, 2005, 11(3): 308–311. |
| [35] | 魏丹, 陈晓飞, 王铁良, 邹艳丽, 杨忠臣. 不同积雪覆盖条件下土壤冻结状况及水分的迁移规律. 安徽农业科学, 2007, 35(12): 3570–3572. DOI:10.3969/j.issn.0517-6611.2007.12.088 |
| [36] | Osterkamp T E. The recent warming of permafrost in Alaska. Global and Planetary Change, 2005, 49(3/4): 187–202. |
| [37] | Gong G, Entekhabi D, Cohen J. Modeled northern hemisphere winter climate response to realistic Siberian snow anomalies. Journal of Climate, 2003, 16(23): 3917–3931. DOI:10.1175/1520-0442(2003)016<3917:MNHWCR>2.0.CO;2 |
| [38] | Brooks P D, Williams M W, Schmidt S K. Microbial activity under alpine snowpacks, Niwot Ridge, Colorado. Biogeochemistry, 1996, 32(2): 93–113. |
| [39] | Mellander P E, Laudon H, Bishop K. Modelling variability of snow depths and soil temperatures in Scots pine stands. Agricultural and Forest Meteorology, 2005, 133(1/4): 109–118. |
| [40] | 常娟, 王根绪, 高永恒, 王一博. 青藏高原多年冻土区积雪对沼泽、草甸浅层土壤水热过程的影响. 生态学报, 2012, 32(23): 7289–7301. |
| [41] | 陈继, 盛煜, 程国栋. 从地表能量平衡各分量特点论青藏高原多年冻土工程中的冻土保护措施. 冰川冻土, 2006, 28(2): 223–228. |
| [42] | Niu G Y, Yang Z L. Effects of frozen soil on snowmelt runoff and soil water storage at a continental scale. Journal of Hydrometeorology, 2006, 7(5): 937–952. DOI:10.1175/JHM538.1 |
| [43] | 刘志民, 杨甲定, 刘新民. 青藏高原几个主要环境因子对植物的生理效应. 中国沙漠, 2000, 20(3): 309–313. |
| [44] | 马雪莹, 李小梅, 祝修高, 傅佳丽, 彭璐. 校园绿化植被对夏季高温的调节功能研究. 福建师大学报:自然科学版, 2016, 32(2): 28–36. |
| [45] | Caldwell M M, Pearcy R W. Exploitation of Environmental Heterogeneity by Plants: Ecophysiological Processes above-and Belowground. New York: Academic Press, 1994. |
| [46] | Loik M E, Redar S P, Harte J. Photosynthetic responses to a climate-warming manipulation for contrasting meadow species in the Rocky Mountains, Colorado, USA. Functional Ecology, 2000, 14(2): 166–175. DOI:10.1046/j.1365-2435.2000.00411.x |
| [47] | 李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895–905. |
| [48] | Walker M D, Ingersoll R C, Webber P J. Effects of interannual climate variation on phenology and growth of two alpine forbs. Ecology, 1995, 76(4): 1067–1083. DOI:10.2307/1940916 |
| [49] | 刘鸿先, 曾韶西, 王以柔, 李平, 陈德峰, 郭俊彦. 低温对不同耐寒力的黄瓜(Cucumic sativus)幼苗子叶各细胞器中超氧物歧化酶(SOD)的影响. 植物生理学报, 1985, 11(1): 48–57. |
| [50] | Pregitzer K S, King J S, Burton A J, Brown S E. Responses of tree fine roots to temperature. New Phytologist, 2000, 147(1): 105–115. DOI:10.1046/j.1469-8137.2000.00689.x |
| [51] | 杜子银, 蔡延江, 王小丹, 鄢燕, 鲁旭阳, 刘淑珍. 土壤冻融作用对植物生理生态影响研究进展. 中国生态农业学报, 2014, 22(1): 1–9. |
| [52] | Herrmann A, Witter E. Sources of C and N contributing to the flush in mineralization upon freeze-thaw cycles in soils. Soil Biology and Biochemistry, 2002, 34(10): 1495–1505. DOI:10.1016/S0038-0717(02)00121-9 |
| [53] | Feng X J, Nielsen L L, Simpson M J. Responses of soil organic matter and microorganisms to freeze-thaw cycles. Soil Biology and Biochemistry, 2007, 39(8): 2027–2037. DOI:10.1016/j.soilbio.2007.03.003 |