 2017, Vol. 37
2017, Vol. 37文章信息
- 管杰然, 商天其, 伊力塔, 叶诺楠, 余树全
- GUAN Jieran, SHANG Tianqi, YI Lita, YE Nuonan, YU Shuquan.
- 天目山常绿落叶阔叶混交林优势种生物量变化及群落演替特征
- Biomass change and community succession characteristics of dominant species in evergreen and deciduous broad-leaved mixed forests in Tianmu Mountain
- 生态学报. 2017, 37(20): 6761-6772
- Acta Ecologica Sinica. 2017, 37(20): 6761-6772
- http://dx.doi.org/10.5846/stxb201605070878
-
文章历史
- 收稿日期: 2016-05-07
- 修订日期: 2017-03-09





2. 亚热带森林培育国家重点实验室培育基地, 临安 311300
2. Nurturing Station for State Key Laboratory of Subtropical Silviculture, Lin'an 311300, China
森林生态系统样地也是近年来全球科研领域对森林生态系统进行结构与功能、内在规律和变化机制研究而采用的新的最科学、最有效的手段[1], 也是对森林生态系统的物质循环及能量平衡进行全面深入研究的不可替代的研究方法[2-4]。同时, 在森林生态系系统中选取具有代表性的典型区域建立固定样地, 对其进行长期的持续性监测和研究, 可以为全球气候和环境变化问题提供价值极高的研究基础数据, 并对解决这些全球首要热门问题提供理论依据、开拓新思路[5]。
设立森林生态系统样地是在研究森林生态系系统的结构与功能、内在规律与变化机制时, 被当今学术界广泛采用和推广的科学、有效的方法[5]。天目山国家级自然保护区是我国东南沿海一带典型的亚热带常绿落叶阔叶混交林, 而亚热带常绿落叶阔叶混交林也是森林生态系统的主要类型之一[5]。因此, 在天目山国家级自然保护区内选取典型的亚热带常绿落叶阔叶混交林设立样地进行定位长期观测和研究, 具有重要的价值和意义。
本文利用天目山自然保护区内设立的亚热带常绿落叶阔叶混交林样地分别于1996年和2012年时间跨度为16年的实地调查监测的数据进行综合处理和分析, 通过天目山自然保护区亚热带常绿落叶阔叶混交林各自优势种的生态位宽度、生态位重叠和种间联结结合生物量进行综合分析, 模拟样地内从1996年到2012年的群落组成和结构的变化, 并对天目山自然保护区亚热带常绿落叶阔叶混交林林生态系统的森林演替情况提供借鉴和参考[5]。
1 研究区概况天目山国家级自然保护区地处浙江省西北部临安市境内(119°24′11″—119°28′21″E, 30°18′30″—30°24′55″N), 浙皖两省交界处, 距杭州84 km, 面积4300 hm2, 海拔300—1556 m, 主峰仙人顶海拔1506 m[6]。天目气候属中亚热带向北亚热带过渡型, 受海洋暖湿气流影响, 季风强盛, 四季分明[7], 气候温和, 年平均气温14.8—8.8℃, 无霜期235—209 d, 年雨日159.2—183.1 d, 年降水量达1390—1870 mm, 春秋季较短, 冬夏季偏长[8], 为典型的中亚热带森林生态系统。该区主要植被类型有:常绿落叶阔叶混交林为典型的地带性植被(海拔850 m以下)、常绿落叶阔叶混交林(海拔850—1100 m)、落叶阔叶混交林(海拔1100—1380 m)及部分竹林和针叶林等。
2 研究方法 2.1 样地设置早在1996年, 选择天目山国家级自然保护区狮子尾典型常绿落叶阔叶混交林内, 建立了一块常绿落叶阔叶混交林固定样地(30.320°N, 119.230°E, 海拔1065 m), 面积为1 hm2(100 m×100 m), 并对样地内胸径(DBH)≥10 cm的木本植物进行每木检尺及每株定位, 记录其种类、胸径、树高、冠幅、枝下高及生长状况, 并用以西南原点和东南点为基点的三角坐标定点法, 记录了检尺植株的位置坐标信息[5]。在样地没有受到人为干扰的情况下, 于2012年再次对样地进行了复查, 同样以西南角为原点, 以东西向为横轴(x)、南北向为纵轴(y), 使用南方测绘NTS-300R全站仪复原1 hm2样地[5]。以标准每木检尺法对样地内胸径(DBH)≥1 cm的木本植物进行每木检尺及每株定位, 记录种类、胸径、树高、冠长等环境因子[9-10]及生长状况, 并用以西南角为原点的直角坐标系定点法, 记录了检尺植株的位置坐标信息[5]。
2.2 数据分析 2.2.1 优势种的确定优势种是指对于群落中其他种有很大影响, 而本身受其他种的影响最小的种[11], 也指群落中具有最大密度、盖度和生物量的种[5]。重要值是森林群落研究中被常用来衡量某个树种优势程度大小的重要指标[12]。
① 乔木层的重要值
层的重要值

|
(1) |
② 灌木、草本层重要值

|
(2) |
式中:相对多度指的是某物种株数占所有种总株数的相对值[5];相对显著度指的是某物种的胸高断面积占所有种的胸高断面积的相对值[5];相对高度指的是某物种的高度占所有种总高度的相对值;相对盖度指的是某物种的盖度占所有种总盖度的相对值[13]。
2.2.2 生态位宽度生态位为正好被一个种(或亚种)所占据的环境限制性因子单元[14], 这是生态位被第一次正式定义[5]。2002年Shea和Chesson重新将生态位定义为物种对每个生态位空间点的反应和效应[15]。本文采用了近几年被广泛引用的Levins[16]生态位宽度公式中的Shannon-Wiener信息指数计算方法。
① Levins指数[5]:
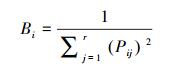
|
(3) |
式中, Bi为种i的生态位宽度;Pij=nij/Ni+, 它代表种i在第j个资源状态下的个体数占该种所有个体数的比例[17]。因此, 该式实际上是Simpson多样性指数[5]。
② Shannon-Wiener信息指数[5]:
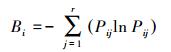
|
(4) |
该指数是以Shannon-Wiener信息公式为基础的。以上两个指数中的Bi值与生态位宽度的大小关系表现为正相关, 即Bi值越大, 说明生态位越宽, 当一个种的个体以相等的数目利用每一资源状态时, Bi值最大化, 即该种具有最宽的生态位;当种i的所有个体都集中在某一个资源状态下时, Bi值最小, 该种具有最窄的生态位[17]。
2.2.3 生态位重叠生态位重叠是指2个物种在利用食物、空间等资源时出现的重叠现象[17]。本文采用的生态位重叠计算公式为Levins(1968) 重叠指数[18]:

|
(5) |
式中, Oik代表种i的资源利用曲线与种k的资源利用曲线的重叠指数[5]。该指数实际上与种i的生态位宽度有关。所以Oik和Oki的值是不同的, 含义也不同[5]。当种i和种k在所有资源状态中的分布完全相同时, Oik最大, 其值为1, 表明种i与种k生态位完全重叠。相反, 当两个种不具有共同资源状态时, 它们的生态位完全不重叠, Oik=0[19]。
2.2.4 种间联结分析种间联结是指不同物种在空间分布上的相互关联性, 通常是由于群落生境的差异影响了物种分布而引起的, 是对各个物种在不同生境中相互影响、相互作用所形成的有机联系的反映[20-21], 它表示种间相互吸引或排斥的性质[22]。它是植物群落重要的数量和结构特征之一, 对正确认识群落结构、功能和分类有重要的指导意义, 并能为植被的经营管理、自然植被恢复和生物多样性保护提供理论依据[23]。
本文根据系数联结AC值和共同出现百分率PC值对研究区内1996年和2012年各自的优势种进行种间联结分析[5]。
(1) 联结系数AC说明种对之间的联结程度[5]。计算公式如下[24]:
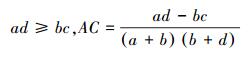
|
(6) |
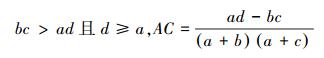
|
(7) |
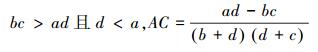
|
(8) |
AC的值域为[-1, 1][5]。当AC值越趋近于1时, 种对间的正联结性越强;当AC值越趋近于-1, 物种间的负联结性越强;当AC值越趋近于0时, 种对间的联结性越弱;当AC值为0, 种对间完全独立[25]。
(2) 共同出现百分率PC测度物种间正联结程度, 计算公式[26]:
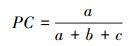
|
(9) |
PC值的大小在0和1之间变动, 其值越接近1, 表明该种对的正联结越紧密;反之, 其值越接近0, 则表明该种对的负联结越强[27]。
2.2.5 生物量变化利用1 hm2固定样地1996年和2012年监测数据, 依照生物量模型(表 1), 推算样地内植被生物量[5]。
| 类别Category | 模型公式Model formula | 模型变量Model variable | 适用树种Applicable tree species |
| 1 | W1 = W2+W3+W4 W2 = 0.0600 H 0.7934 D 1.8005 W3 = 0.137708 D1.487266 L 0.405207 W4 = 0.0417 H -0.0780 D 2.2618 |
W1为总生物量(kg), W2为树干生物量(kg), W3为树冠生物量(kg), W4为树根生物量(kg); |
马尾松(Pius. massoniana)、黄山松(P. taiwanensis)等 |
| 2 | W1 = W2+W3+W4 W2 = 0.064 7 H 0.895 9 D 1.488 0 W3 = 0.097 1 D 1.781 4 L 0.034 6 W4 = 0.061 7 H -0.103 74 D 2.115 252 |
H为树高(m), D为胸径(cm), L为冠长(m) | 杉木(Cunninghamia lanceolat)、柳杉(Cryptomeria fortunei)等 |
| 3 | W1 = W2+W3+W4 W2 = 0.056 0 H 0.809 9 D 1.814 0 W3 = 0.098 0 D 1.648 1 L 0.461 0 W4 = 0.054 9 H 0.106 8 D 2.095 3 |
青钱柳(Cyclocarya paliurus)、交让木(Daphniphyllum macropodum)、微毛柃(Eurya hebeclados)、缺萼枫香(Liquidambar acalycina)、天目木姜子(Litsea auriculata)、红果钓樟(Lindera erythrocarpa)等 | |
| 4 | W1 = W2+W3+W4 W2 = 0.080 3 H 0.781 5 D 1.805 6 W3 = 0.286 0 D 1.096 8 L 0.945 0 W4 = 0.247 0 H 0.174 5 D 1.795 4 |
小叶青冈(Cyclobalanopsis myrsinifolia)、东南石栎(Lithocarpus harlandi)、褐叶青冈(Cyclobalanopsis stewardiana)、梓树(Catalpa ovata)、蓝果树(Nyssa sinensis)等 | |
| 5 | W1 = W2+W3+W4 W2 = 0.044 4 H 0.719 7 D 1.709 5 W3 = 0.085 6 D 1.226 57 L 0.397 0 W4 = 0.045 9 H 0.106 7 D 2.024 7 |
枫香(Liquidamba formosana)、檫木(Sassafras tzumu)等 | |
| 6 | W1 = W2+W3+W4 W2 = 0.039 8 H 0.577 8 D 1.854 0 W3 = 2.80×10-1 D 0.835 7 L 0.274 0 W4 = 3.71×10-1 H 0.135 7 D 0.981 7 |
毛竹(Phyllostachys heterocycla cv. pubescens) |
表 1中的适用树种为适用于该表生物量模型的样地中记录在内的树种。
模型公式中:胸径(D)使用钢围尺在被测树的树干1.3 m高处标准测量获得;树高(H)使用布鲁莱斯测高器以正确使用方法测量获得;冠长(L)使用皮尺对被测树树冠的平面投影长度进行测量获得。
3 结果与分析 3.1 重要值1996年样地中胸径(DBH)≥10 cm的树42种, 546株;2012年样地中胸径(DBH)≥10 cm的树51种, 606株。依据重要值计算公式, 在得到的结果中筛选出1996年中12种重要值≥10%的物种和2012年中12种重要值≥10%的物种分别作为两年内各自的优势种(表 2)。
| 序号 Serial number |
1996年 | 2012年 | ||
| 优势树种 Dominant species |
重要值/% Importance value |
优势树种 Dominant species |
重要值/% Importance value |
|
| 1 | 青钱柳Cyclocarya paliurus | 53.66 | 小叶青冈Cyclobalanopsis myrsinifolia | 40.86 |
| 2 | 杉木Cunninghamia lanceolata | 41.61 | 杉木Cunninghamia lanceolata | 39.36 |
| 3 | 小叶青冈Cyclobalanopsis myrsinifolia | 21.46 | 青钱柳Cyclocarya paliurus | 35.39 |
| 4 | 缺萼枫香Liquidambar acalycina | 21.02 | 交让木Daphniphyllum macropodum | 32.34 |
| 5 | 交让木Daphniphyllum macropodum | 14.50 | 东南石栎Lithocarpus harlandii | 28.33 |
| 6 | 微毛柃Eurya hebeclados | 14.19 | 缺萼枫香Liquidambar acalycina | 21.25 |
| 7 | 天目木姜子Litsea auriculata | 12.31 | 四照花Cornus kousa subsp. chinensis | 18.66 |
| 8 | 四照花Cornus kousa subsp. chinensis | 12.02 | 柳杉Cryptomeria fortunei | 18.17 |
| 9 | 柳杉Cryptomeria fortunei | 11.77 | 天目木姜子Litsea auriculata | 17.28 |
| 10 | 东南石栎Lithocarpus harlandii | 11.16 | 微毛柃Eurya hebeclados | 14.10 |
| 11 | 梓树Catalpa ovata | 10.86 | 蓝果树Nyssa sinensis | 11.49 |
| 12 | 红果钓樟Lindera erythrocarpa | 10.05 | 褐叶青冈Cyclobalanopsis stewardiana | 10.61 |
由表 2可知, 1996年样地中重要值≥10%的12个优势种按重要值大小排列分别为:青钱柳、杉木、小叶青冈、缺萼枫香、交让木、微毛柃、天目木姜子、四照花、柳杉、东南石栎、梓树、红果钓樟;而在2012年重要值≥10%的12个优势种按重要值大小排列分别为:小叶青冈、杉木、青钱柳、交让木、东南石栎、缺萼枫香、四照花、柳杉、天目木姜子、微毛柃、蓝果树、褐叶青冈[5]。
3.2 生态位宽度由表 3可知, 在1996年调查中发现:青钱柳的生态位宽度指数最大, Bi值为1.3165, 其他优势种的生态位宽度指数按由大到小的顺序为杉木>交让木>小叶青冈>缺萼枫香>微毛柃>四照花>天目木姜子>红果钓樟>柳杉>梓树>东南石栎[5];而在2012年变化为:小叶青冈的生态位宽度指数最大, Bi值为1.3177, 其他优势种的生态位宽度指数按由大到小的顺序为杉木>青钱柳>交让木>东南石栎>四照花>天目木姜子>微毛柃>柳杉>缺萼枫香>蓝果树>褐叶青冈[5]。
| 1996年 | 2012年 | 差值 Difference |
||
| 优势种 Dominant species |
Bi | 优势种 Dominant species |
Bi | |
| 东南石栎Lithocarpus harlandii | 0.6762 | 东南石栎Lithocarpus harlandii | 1.1655 | 0.4893 |
| 交让木Daphniphyllum macropodum | 0.9788 | 交让木Daphniphyllum macropodum | 1.1788 | 0.2000 |
| 柳杉Cryptomeria fortunei | 0.8205 | 柳杉Cryptomeria fortunei | 0.8796 | 0.0591 |
| 青钱柳Cyclocarya paliurus | 1.3165 | 青钱柳Cyclocarya paliurus | 1.2274 | -0.0890 |
| 缺萼枫香Liquidambar acalycina | 0.9146 | 缺萼枫香Liquidambar acalycina | 0.8794 | -0.0353 |
| 杉木Cunninghamia lanceolata | 1.2646 | 杉木Cunninghamia lanceolata | 1.3071 | 0.0425 |
| 四照花Cornus kousa subsp. chinensis | 0.8513 | 四照花Cornus kousa subsp. chinensis | 1.0646 | 0.2133 |
| 天目木姜子Litsea auriculata | 0.8289 | 天目木姜子Litsea auriculata | 1.0004 | 0.1715 |
| 微毛柃Eurya hebeclados | 0.8691 | 微毛柃Eurya hebeclados | 0.9284 | 0.0593 |
| 小叶青冈Cyclobalanopsis myrsinifolia | 0.9170 | 小叶青冈Cyclobalanopsis myrsinifolia | 1.3177 | 0.4007 |
| 红果钓樟Lindera erythrocarpa | 0.8205 | 褐叶青冈Cyclobalanopsis stewardiana | 0.7696 | — |
| 梓树Catalpa ovata | 0.7846 | 蓝果树Nyssa sinensis | 0.7760 | — |
在1996年样地胸径(DBH)≥10 cm的优势种中, 青钱柳生态位宽度指数Bi值最大, 说明了青钱柳在1996年时具有最大生长优势和群落地位, 而到了2012年其生态位宽度指数Bi值有所下降(Bi=1.2274, 差值为-0.0890), 且青钱柳是在16年间生态位宽度指数Bi值是下降幅度最大的优势种。从1996年到2012年生态位宽度指数Bi值变小的优势种还有缺萼枫香(1996年Bi值为0.9146, 2012年Bi值为0.8794, 差值为-0.0353), 其优势地位从1996年的第6位变为2012年的第10位;其余10个优势种的生态位宽度指数Bi值从1996年到2012年皆增大:在1996年中, Bi值为0.9170(第5位)的小叶青冈到了2012年成为了生态位宽度指数Bi值最大的优势种, Bi值达到1.3177, 差值为0.4007, 成为了样地内最具优势地位的物种[5]。而从1996年到2012年间生态位宽度指数Bi值增大幅度最大的(差值最大)优势种是东南石栎, 其Bi值从1996年排在第12的0.6762到2012年上升到了排在第5的1.1655, 差值为0.4893, 其优势地位显著增加。同样可以发现:样地中, 生态位宽度指数Bi值从1996年到2012年变化很小, (差值很小)的优势种有3个, 分别为柳杉(差值为0.0591, )、杉木(差值为0.0425) 和微毛柃(差值为0.0593)[5]。
3.3 生态位重叠由表 4可知, 相比于1996年, 2012年样地中优势种的生态位重叠值<0.1的大大减少(30对变为4对, 22.73%变为3.03%), 而生态位重叠值>0.5的数量显著增加(25对变为53对, 18.94%变为31.82%), 且1996年中存在4对不重叠的生态位(东南石栎和四照花、四照花和东南石栎、东南石栎对红果钓樟、红果钓樟对东南石栎), 而2012年不存在不重叠的生态位[5]。这表明, 2012年的优势种相比于1996年, 生态位重叠程度更高, 对资源的共享和利用程度更高[5]。
| 取值范围 Range |
1996年 | 2012年 | ||
| 对数 Logarithm |
% | 对数 Logarithm |
% | |
| <0.1 | 30 | 22.73 | 4 | 3.03 |
| 0.1—0.3 | 48 | 36.36 | 47 | 35.61 |
| 0.3—0.5 | 25 | 18.94 | 39 | 29.55 |
| >0.5 | 25 | 18.94 | 42 | 31.82 |
| 无重叠No overlap | 4 | 3.03 | — | — |
| 总计Total | 132 | 100.00 | 132 | 100.00 |
(1) 联结系数AC值分析
对1996年优势种的种间联结分析进行联结系数AC值计算结果(图 1)所示, 对2012年优势种的种间联结分析进行联结系数AC值计算结果(图 2)所示。

|
| 图 1 1996年优势种AC值 Fig. 1 AC value of dominant species in 1996 1:东南石栎, Lithocarpus harlandii;2:红果钓樟, Lindera erythrocarpa;3:交让木, Daphniphyllum macropodum;4.柳杉, Cryptomeria fortunei;5:青钱柳, Cyclocarya paliurus;6:缺萼枫香, Liquidambar acalycina;7:杉木, Cunninghamia lanceolata;8:四照花, Cornus kousa subsp. chinensis;9:天目木姜子, Litsea auriculata;10:微毛柃, Eurya hebeclados;11:小叶青冈, Cyclobalanopsis myrsinifolia;12:梓树, Catalpa ovata |

|
| 图 2 2012年优势种AC值 Fig. 2 AC value of dominant species in 2012 1:东南石栎, Lithocarpus harlandii;2:褐叶青冈, Cyclobalanopsis stewardiana;3:交让木, Daphniphyllum macropodum;4:蓝果树, Nyssa sinensis;5:柳杉, Cryptomeria fortunei;6:青钱柳, Cyclocarya paliurus;7:缺萼枫香, Liquidambar acalycina;8:杉木, Cunninghamia lanceolata;9:四照花, Cornus kousa subsp. chinensis;10:天目木姜子, Litsea auriculata;11:微毛柃, Eurya hebeclados;12:小叶青冈, Cyclobalanopsis myrsinifolia |
由图 1、图 2和表 5可知:1996年12种重要值≥10%的优势种共66对种间联结联结系数AC值结果中, 表现为正联结的有19对, 占总对数的28.79%, 其中极显著正关联的有1对, 占1.52%、显著正关联的有10对, 占15.15%、不显著正相关的有8对, 占12.12%[5]。表现为无关联的有18对, 占27.27%。表现为负联结的有29对, 占总对数的43.94%, 其中不显著负关联的有6对, 占9.09%、显著负关联的有17对, 占25.76%、极显著负关联的有6对, 占9.09%;2012年12种重要值≥10%的优势种共66对种间联结联结系数AC值结果中, 表现为正联结的有38对, 占总对数的57.58%, 其中极显著正关联的有13对, 占19.70%、显著正关联的有12对, 占18.18%、不显著正相关的有13对, 占19.70%。表现为无关联的有9对, 占13.64%[5]。表现为负联结的有19对, 占总对数的28.79%, 其中不显著负关联的有7对, 占10.61%、显著负关联的有8对, 占12.12%、极显著负关联的有4对, 占6.06%[5]。1996年和2012年的正负关联比分别为0.66和2.0[5]。
| 联结性 Connection |
AC | 1996年 | 2012年 | ||
| 种对数 Species logarithm |
% | 种对数 Species logarithm |
% | ||
| 正联结 | AC≥0.60 | 1 | 1.52 | 13 | 19.7 |
| Positive connection | 0.20≤AC<0.60 | 10 | 15.15 | 12 | 18.18 |
| 0.05≤AC<0.20 | 8 | 12.12 | 13 | 19.7 | |
| 负联结 | AC<-0.60 | 6 | 9.09 | 4 | 6.06 |
| Negative connection | -0.60≤AC<-0.20 | 17 | 25.76 | 8 | 12.12 |
| -0.20≤AC<-0.05 | 6 | 9.09 | 7 | 10.61 | |
| 无关联No association | -0.05≤AC<0.05 | 18 | 27.27 | 9 | 13.64 |
| AC值:联结系数, association coefficient | |||||
(2) 共同出现百分率PC值分析
对1996年优势种的种间联结分析进行共同出现百分率PC值计算结果(图 3)所示, 对2012年优势种的种间联结分析进行共同出现百分率PC值计算结果(图 4)所示。

|
| 图 3 1996年优势种PC值 Fig. 3 PC value of dominant species in 1996 1:东南石栎, Lithocarpus harlandii;2:红果钓樟, Lindera erythrocarpa;3:交让木, Daphniphyllum macropodum;4:柳杉, Cryptomeria fortunei;5.青钱柳, Cyclocarya paliurus;6:缺萼枫香, Liquidambar acalycina;7:杉木, Cunninghamia lanceolata;8:四照花, Cornus kousa subsp. chinensis;9:天目木姜子, Litsea auriculata;10:微毛柃, Eurya hebeclados;11:小叶青冈, Cyclobalanopsis myrsinifolia;12:梓树, Catalpa ovata |

|
| 图 4 2012年优势种PC值 Fig. 4 PC value of dominant species in 2012 1:东南石栎, Lithocarpus harlandii;2:褐叶青冈, Cyclobalanopsis stewardiana;3:交让木, Daphniphyllum macropodum;4:蓝果树, Nyssa sinensis;5:柳杉, Cryptomeria fortunei;6:青钱柳, Cyclocarya paliurus;7:缺萼枫香, Liquidambar acalycina;8:杉木, Cunninghamia lanceolata;9:四照花, Cornus kousa subsp. chinensis;10:天目木姜子, Litsea auriculata;11:微毛柃, Eurya hebeclados;12:小叶青冈, Cyclobalanopsis myrsinifolia |
由图 3、图 4和表 6可知, 1996年12种重要值≥10%的优势种共66对种间联结共同出现百分率PC值结果中, 表现为正联结的有32对, 占总对数的48.48%, 其中极显著正关联的有1对, 占1.52%、显著正关联的有10对, 占15.15%、不显著正相关的有21对, 占31.82%[5]。表现为无关联的有3对, 占4.55%;2012年12种重要值≥10%的优势种共66对种间联结共同出现百分率PC值结果中, 表现为正联结的有47对, 占总对数的71.21%, 其中极显著正关联的有5对, 占7.58%、显著正关联的有21对, 占31.82%、不显著正相关的有21对, 占31.82%。表现为无关联的有0对[5]。
| 联结性 Connection |
PC | 1996年 | 2012年 | ||
| 种对数 Species logarithm |
% | 种对数 Species logarithm |
% | ||
| 正联结 | PC≥0.7 | 1 | 1.52 | 5 | 7.58 |
| Positive connection | 0.4≤PC<0.7 | 10 | 15.15 | 21 | 31.82 |
| 0.2≤PC<0.4 | 21 | 31.82 | 21 | 31.82 | |
| 负联结Negative connection | — | — | — | — | — |
| 无关联No association | 0≤PC<0.2 | 3 | 4.55 | 0 | 0 |
| PC值:共同出现百分率percentage co-occurrence | |||||
根据1996年和2012年天目山常绿落叶阔叶混交林样地中, 胸径(DBH)≥10 cm的乔木树种的生物量统计中发现其生物量分别为151.03 t和148.53 t, 单位生物量分别达到了151.03 t/hm2和148.53 t/hm2。即, 2012年乔木生物总量反而比1996年减少了2.50 t[5]。鉴于此, 随着16年的时间跨度, 样地中胸径(DBH)≥10 cm的乔木生物总量减少的问题, 进行了进一步的分析[5]。
由表 7可知, 1996年和2012年样地中胸径(DBH)≥10 cm的优势种的生物量分别为79.14 t和128.61 t, 增大了49.47 t[5]。其中, 作为1996年优势种而在2012年中丢失优势地位的红果钓樟和梓树的生物量分别为0.59 t和5.05 t, 总量为5.64 t;在1996年林分中未出现的褐叶青冈和蓝果树到了2012年成为了主要的优势树种, 重要值分别达到10.61%和11.49%, 其胸径(DBH)≥10 cm的植株生物量分别达到4.25 t和8.37 t, 总量为12.62 t[5]。另外, 作为两个年份中都是主要优势树种的东南石栎、交让木、小叶青冈和柳杉, 从1996年到2012年间, 重要值分别增大了17.81%、17.72%、22.25%、0.23%, 生物量分别增加了3.99、8.42、16.64、11.74 t, 较各自在1996年生物量的涨幅分别为129.90%、174.46%、146.09%和164.81%, 生物量的增长量和增长速率远远大于其他树种(包括1996年的主要优势树种)[5]。
| 优势种Dominant species | 生物量Biomass/t | ||||
| 1996年 | 2012年 | 1996年 | 2012年 | 差值 Difference |
涨幅/% amount of increase |
| 东南石栎 | 东南石栎 | 3.07 | 7.06 | 3.99 | 129.90 |
| 交让木 | 交让木 | 4.83 | 13.25 | 8.42 | 174.46 |
| 柳杉 | 柳杉 | 7.12 | 18.86 | 11.74 | 164.81 |
| 青钱柳 | 青钱柳 | 14.84 | 11.49 | -3.35 | -22.60 |
| 缺萼枫香 | 缺萼枫香 | 10.34 | 13.06 | 2.72 | 26.27 |
| 杉木 | 杉木 | 12.04 | 13.56 | 1.52 | 12.61 |
| 四照花 | 四照花 | 1.12 | 1.82 | 0.71 | 63.51 |
| 天目木姜子 | 天目木姜子 | 7.33 | 8.09 | 0.75 | 10.29 |
| 微毛柃 | 微毛柃 | 1.42 | 0.78 | -0.64 | -44.96 |
| 小叶青冈 | 小叶青冈 | 11.39 | 28.03 | 16.64 | 146.09 |
| 红果钓樟 | 褐叶青冈 | 0.59 | 4.25 | — | — |
| 梓树 | 蓝果树 | 5.05 | 8.37 | — | — |
| 总计Total | 79.14 | 128.61 | 49.47 | — | |
由图 5可知, 2012年样地中胸径在1—5 cm和5—10 cm的乔木数量非常大, 为3785株, 占乔木总量的86.20%。据测算, 2012年中胸径在1—10 cm的乔木总生物量为25.15 t, 其中2012年12个优势种1—10 cm的的植株总生物量为16.03 t, 占63.74%。由此可知, 虽然2012年样地乔木现阶段生物量的值不大, 但是生物量的增长潜力非常大[5]。

|
| 图 5 样地乔木径级分布 Fig. 5 Diameter distribution of the trees in 1996 and 2012 |
根据表 8, 研究中发现:1996年样地中的10棵胸径(DBH)>60 cm的大树, 在没有受到人为干扰的情况下, 到了2012年死亡(包括断顶死亡、倒地死亡、腐烂死亡)的8棵大树中, 绝大部分是作为1996年主要优势种的柳杉、缺萼枫香、青钱柳, 它们总生物量的和达到了10.33 t, 占到了1996年中胸径(DBH)≥10 cm乔木树总生物量的6.84%[5]。可见, 1996年中生物量较大的大树的死亡也是导致2012年样地内的生物总量减小的重要原因。
| 优势树种 Dominant species |
胸径/cm diameter at breast height |
树高/m Tree height |
枝下高/m Clear bole height/m |
生物量/t Biomass |
| 黄山松Pinus taiwanensis Hayata * | 69.40 | 30.00 | 17.50 | 2.49 |
| 柳杉Cryptomeria fortunei * | 68.70 | 30.00 | 17.00 | 1.25 |
| 柳杉Cryptomeria fortunei | 61.40 | 30.00 | 18.00 | 1.13 |
| 柳杉Cryptomeria fortunei * | 60.20 | 35.00 | 22.00 | 1.10 |
| 柳杉Cryptomeria fortunei * | 68.10 | 25.00 | 12.50 | 1.14 |
| 柳杉Cryptomeria fortunei * | 64.40 | 35.00 | 22.00 | 1.23 |
| 柳杉Cryptomeria fortunei | 62.80 | 35.00 | 21.00 | 1.18 |
| 柳杉Cryptomeria fortunei * | 63.70 | 32.00 | 19.50 | 1.15 |
| 缺萼枫香Liquidambar acalycina * | 68.30 | 30.00 | 17.50 | 1.08 |
| 青钱柳Cyclocarya paliurus * | 64.00 | 25.00 | 12.00 | 0.88 |
| 总计Total | — | — | — | 12.64 |
| *为死亡大树 | ||||
基于对1996年和2012年样地监测数据的整理和计算结果, 本文意在阐述和分析天目山常绿落叶阔叶混交林从1996年到2012年的优势种变化特征, 并分析其群落的组成和结构的变化及发展趋势[5, 29-30]。从本文的计算分析中可知:2012年的优势种相比于1996年, 生态位重叠程度更高, 对资源的共享和利用程度更高[5, 19-31];且根据种间联结的AC值和PC值分析结果得到, 2012年的优势种相比于1996年, 负联结和无关联的比例减小, 正关联比例增加, 特别是极显著正关联和显著正关联的比例显著增加。从森林群落演替的角度出发, 亚热带常绿落叶阔叶混交林的顶级群落为耐荫的常绿阔叶林[32]。1996年到2012年间的天目山常绿落叶阔叶混交林的优势种及其变化表明了其正处于森林群落演替的中期, 在这个演替阶段中, 其发生着以下显著的变化:
(1) 从本文的计算分析中可知:1996年样地中, 青钱柳生态位宽度指数Bi值最大, 同时青钱柳也是1996年样地中重要值最大的树种, 这说明青钱柳在1996年时具有最明显的生长优势和最大的群落地位[5, 33]。并且, 1996年样地中, 青钱柳的生物量也是最大的, 这说明青钱柳在1996年时是群落中生长情况最好的树种[5];而到了2012年青钱柳生态位宽度指数Bi值明显有所下降, 且在12个优势种中青钱柳的下降幅度最大, 青钱柳的生物量也是1996年12个优势种当中唯一一个降低的树种[5], 其株数也从1996年的79株下降到了2012年的62株。结合青钱柳和其他优势种的种间联结结果, 充分说明了青钱柳在1996年到2012年间的林分水平呈现出衰退的趋势, 这说明青钱柳在天目山常绿落叶阔叶混交林的演替过程中处于劣势甚至逐渐被淘汰[5]。
此外, 样地中从1996年到2012年间, 生态位宽度指数Bi值及生物量的涨幅非常小甚至减少, 优势度下降的优势种还有柳杉、杉木、微毛柃、缺萼枫香红果钓樟和梓树(其中只有柳杉的生物量涨幅很大, 原因是柳杉本身作为天目山的主要树种, 生物量基数非常大), 并且红果钓樟和梓树到了2012年失去了优势地位, 说明这6个树种在天目山常绿落叶阔叶混交林的演替过程中不具有优势[5]。
(2) 从本文的计算分析中还可以看到:在1996年中, 重要值和生态位宽度指数Bi值为均排在第5位的小叶青冈到了2012年成为了重要值和生态位宽度指数Bi值均显著的优势种, 成为了2012年样地内最具优势地位的树种[5]。并且, 小叶青冈在的生物量从1996年到2012年也有较大的增加, 但涨幅没有达到峰值的原因是小叶青冈的生物量有很大比重分布在了胸径5—10 cm的幼树中, 且在2012年样地内胸径在5—10 cm的幼树生物量统计中小叶青冈的值很大, 表明小叶青冈其生物量的增长潜力非常大[5]。结合小叶青冈在1996年和2012年的种间联结分析结果, 表明了小叶青冈在1996年到2012年间的生长呈现出良好增长的趋势, 这说明小叶青冈在天目山常绿落叶阔叶混交林的演替过程中处于明显优势的地位[5]。
同样值得一提的是1996年东南石栎的重要值和生态位宽度指数Bi值都排在第12位, 但在2012年其重要值和生态位宽度指数Bi值都排在了第5位, 东南石栎和小叶青冈的重要值和生态位宽度指数Bi值从1996年到2012年的增长幅度是样地优势种中最大的[5]。并且, 东南石栎的生物量在1996年时是12个优势种中最小的, 到了2012年时其生物量有了明显上涨, 涨幅超过100%[5]。这说明东南石栎在1996年到2012年间的生长呈现出良好增长的趋势, 这说明东南石栎在天目山常绿落叶阔叶混交林的演替过程中处于明显优势的地位[5]。
同时, 样地中从1996年到2012年间, 生态位宽度指数Bi值及生物量的涨幅有明显上涨, 优势度增大的优势种还有交让木、四照花和天目木姜子(其中交让木从1996年到2012年间生物量的涨幅是12个主要优势种中最大的, 接近200%)[5]。此外, 蓝果树和褐叶青冈到了2012年成为了主要优势树种, 说明这5个树种在天目山常绿落叶阔叶混交林的演替过程中具有优势。
4.2 结论综上所述, 通过本文研究可以得出, 从1996年到2012年间, 天目山常绿落叶阔叶混交林样地演替过程中[5]:小叶青冈和东南石栎具有明显的优势地位, 褐叶青冈、蓝果树、交让木、四照花和天目木姜子也具有一定的优势[5];而梓树、红果钓樟、柳杉、杉木、微毛柃和缺萼枫香不具有优势, 且青钱柳处于劣势甚至被逐渐淘汰的地位。这与天目山常绿落叶阔叶混交林演替趋势相吻合(从针阔混交林逐渐转型为阔叶林, 阔叶树种将取代柳杉等针叶树种成为主要优势种)[5, 34-38]。
| [1] | 罗佳, 张灿明, 牛艳东, 姚敏. 森林生态系统大样地定位研究综述. 湖南林业科技, 2013, 40(4): 79–81. |
| [2] | 周晓峰. 森林生态功能与经营途径. 北京:中国林业出版社, 1999: 1–4. |
| [3] | 李伟民, 甘先华. 国内外森林生态系统定位研究网络的现状与发展. 广东林业科技, 2006, 22(3): 104–108. |
| [4] | 刘峰, 刘红霞, 梁军, 张星耀. 中国森林生态系统定位研究现状与趋势. 安徽农学通报, 2007, 13(11): 89–91. DOI:10.3969/j.issn.1007-7731.2007.11.039 |
| [5] | 管杰然. 天目山常绿落叶阔叶林群落主要结构特征变化分析. 浙江农林大学, 2016. DOI:10.11833/j.issn.2095-0756.2016.06.001 |
| [6] | 浙江天目山国家级自然保护区. 浙江林业, 2007, (8): 34-34. |
| [7] | 方国景, 汤孟平. 天目山常绿阔叶林优势种群胸径的空间连续性分析. 浙江农林大学学报, 2014, 31(5): 663–667. DOI:10.11833/j.issn.2095-0756.2014.05.001 |
| [8] | 尤志勉, 张森尧, 姚振生. 浙江天目山自然保护区药用蕨类植物资源调查. 亚热带植物科学, 2007, 36(3): 59–63. |
| [9] | 张华柳, 伊力塔, 余树全, 余小龙, 陈亚峰, 张朋伟. 嵊州市公益林生物量及生态效益价值评价. 林业资源管理, 2011(1): 78–85. |
| [10] | 伊力塔, 严晓素, 余树全, 包春泉, 屠永海, 潘昌尧. 浙江省不同森林类型林分健康指标体系. 南京林业大学学报:自然科学版, 2012, 36(1): 145–148. |
| [11] | 张金屯. 数量生态学(第二版). 北京:科学出版社, 2011: 19–20. |
| [12] | 王育松, 上官铁梁. 关于重要值计算方法的若干问题. 山西大学学报:自然科学版, 2010, 33(2): 312–316. |
| [13] | 徐悦, 钱逸凡, 伊力塔, 余树全. 普陀山森林群落结构与生物多样性研究. 浙江林业科技, 2013, 33(5): 74–80. |
| [14] | Grinnell J. The niche-relationships of the California thrasher. The Auk, 1917, 34(4): 427–433. DOI:10.2307/4072271 |
| [15] | Shea K, Chesson P. Community ecology theory as a framework for biological invasions. Trends in Ecology & Evolution, 2002, 17(4): 170–176. |
| [16] | Levins R. Evolution in Changing Environments: Some Theoretical Explorations. Princeton: Princeton University Press, 1968: 95–99. |
| [17] | 张金屯. 数量生态学(第二版). 北京:科学出版社, 2011: 115–117. |
| [18] | 郝兴明, 李卫红, 陈亚宁. 新疆塔里木河下游荒漠河岸(林)植被合理生态水位. 植物生态学报, 2008, 32(4): 838–847. |
| [19] | 伊力塔, 韩海荣, 豪树奇, 康峰峰. 灵空山辽东栎群落特征及其物种多样性. 林业科学, 2010, 46(9): 164–171. DOI:10.11707/j.1001-7488.20100927 |
| [20] | Wang B S, Li M G, Peng S L. Acta Population. Guangzhou: Sunyatseni University Press, 1989. |
| [21] | Greig-Smith P. Quantitative Plant Ecology. 3rd ed. Oxford: Blackwell Scientific Publications, 1983: 105–128. |
| [22] | 史作民, 刘世荣, 程瑞梅, 蒋有绪. 宝天曼落叶阔叶林种间联结性研究. 林业科学, 2001, 37(2): 29–35. DOI:10.11707/j.1001-7488.20010204 |
| [23] | 张金屯. 数量生态学. 北京:科学出版社, 2004. |
| [24] | 王乃江, 张文辉, 陆元昌, 范少辉, 王勇. 陕西子午岭森林植物群落种间联结性. 生态学报, 2010, 30(1): 67–78. |
| [25] | 刘金福, 洪伟, 樊后保, 林荣福. 天然格氏栲林乔木层种群种间关联性研究. 林业科学, 2001, 37(4): 117–123. DOI:10.11707/j.1001-7488.20010419 |
| [26] | 郭志华, 卓正大, 陈洁, 吴梅凤. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 1997, 21(5): 424–432. |
| [27] | 张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学, 2015, 51(1): 12–21. |
| [28] | 袁位高, 江波, 葛永金, 朱锦茹, 沈爱华. 浙江省重点公益林生物量模型研究. 浙江林业科技, 2009, 29(2): 1–5. |
| [29] | 丁松, 欧阳杰, 吕丹, 欧阳勋志. 井冈山国家自然保护区栲树林优势种群生态位特征. 中南林业科技大学学报, 2014, 34(7): 99–103. |
| [30] | 陈志伟, 伊贤贵, 王贤荣, 谢春平, 南程慧, 汪小飞. 黄山微毛樱群落主要种群生态位特征. 南京林业大学学报:自然科学版, 2014, 38(S1): 39–46. |
| [31] | 赵永华, 雷瑞德, 何兴元, 贾夏. 秦岭锐齿栎林种群生态位特征研究. 应用生态学报, 2004, 15(6): 913–918. |
| [32] | 魏强, 凌雷, 张广忠, 柴春山, 王多锋, 陶继新, 薛睿. 兴隆山森林群落不同演替阶段优势乔木种群结构特征. 南京林业大学学报:自然科学版, 2015, 39(5): 59–66. |
| [33] | 吴友贵, 叶珍林, 周荣飞, 杨辉, 骆争荣. 百山祖常绿阔叶林优势种群的生态位. 广西植物, 2016, 36(2): 186–192. DOI:10.11931/guihaia.gxzw201408035 |
| [34] | 丁圣彦, 宋永昌. 常绿阔叶林植被动态研究进展. 生态学报, 2004, 24(8): 1765–1775. |
| [35] | 周小勇, 黄忠良, 欧阳学军, 李炯, 官丽莉, 徐国良, 张池. 鼎湖山季风常绿阔叶林原锥栗-厚壳桂-荷木群落演替. 生态学报, 2005, 25(1): 37–44. |
| [36] | 彭少麟, 方炜, 任海, 黄忠良, 孔国辉, 余清发, 张德强. 鼎湖山厚壳桂群落演替过程的组成和结构动态. 植物生态学报, 1998, 22(3): 245–249. |
| [37] | 张家城, 陈力. 亚热带多优势种森林群落演替现状评判研究. 林业科学, 2000, 36(2): 116–121. DOI:10.11707/j.1001-7488.20000219 |
| [38] | 袁金凤, 胡仁勇, 慎佳泓, 张磊, 张晓宇, 于明坚. 4种不同演替阶段森林群落物种组成和多样性的比较研究. 植物研究, 2011, 31(1): 61–66. |