 2017, Vol. 37
2017, Vol. 37文章信息
- 耿静, 程淑兰, 方华军, 于贵瑞, 徐敏杰, 王磊, 李晓玉, 司高月, 何舜
- GENG Jing, CHENG Shulan, FANG Huajun, YU Guirui, XU Minjie, WANG Lei, LI Xiaoyu, SI Gaoyue, HE Shun.
- 氮素类型和剂量对寒温带针叶林土壤N2O排放的影响
- The effects of types and doses of nitrogen addition on soil N2O flux in a cold-temperate coniferous forest, northern China
- 生态学报[J]. 2017, 37(2): 395-404
- Acta Ecologica Sinica[J]. 2017, 37(2): 395-404
- http://dx.doi.org/10.5846/stxb201508041639
-
文章历史
- 收稿日期: 2015-08-04
- 网络出版日期: 2016-06-13





2. 中国科学院大学资源与环境学院, 北京 100049
2. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
氧化亚氮(N2O)是地球大气中三大温室气体之一,百年尺度上单分子N2O的增温潜势(GWP)是CH4和CO2的21倍和298倍,对全球变暖的贡献约占6%[1]。同时,对流层中的N2O通过扩散进入平流层,与O3反应生成NO破坏臭氧层,增加地面紫外辐射量[2]。IPCC第五次报告表明,2011年大气中N2O浓度为324 ppb,较工业化前的数值高出20%,近几十年平均增幅为0.25%/a。大气N2O的源估计为17.7 Tg N/a,自然源(农业、水体、生物燃烧等)和人为源(自然植被土壤、海洋)分别占37.8%和62.2%;大气N2O的汇估计为12.6 Tg N/a,主要在平流层被光解为NOx,最终转化成硝酸和硝酸盐等反应性氮[3]。其中,60%—70%的大气反应性氮以干湿沉降形式到达地表,导致当前全球大气氮沉降量高达105 Tg N/a[4-5],显著改变陆地生态系统碳、氮循环,降低生物多样性,进而影响陆地生态系统的结构和功能[6]。
自然植被下的土壤是N2O的主要排放源,占总自然源的60%[3, 7]。土壤N2O主要来源于硝化、反硝化和化学还原过程[8],介导上述过程的功能微生物群落包括硝化细菌、反硝化细菌以及部分菌根真菌[9]。土壤有效氮浓度、温度、水分、pH值、C/N均会显著改变土壤N2O产生菌群落的活性与组成,进而影响土壤N2O排放[10]。研究表明,外源性氮素输入对自然植被土壤N2O排放的影响有促进[11]、不变[12]和抑制[13]等多种结论,取决于生态系统类型、初始有效氮状态、土壤属性、施氮剂量、施氮类型与持续时间[14]。过去10多年来,有关森林、草地、湿地等自然植被土壤N2O排放对氮沉降增加的响应已开展了许多研究[15-16],取得了一些共识,但仍然存在以下两方面的不足:(1)过去氮沉降模拟控制实验施氮剂量普遍较高(一般>50 kg N hm-2 a-1),短期内只能观测到生态系统“氮饱和”中期或后期的响应特征,鲜有研究贫氮自然植被土壤N2O排放对多形态、低剂量氮输入的响应规律。(2)由于氧化态NO3-和还原态NH4+在土壤中的行为截然不同,过去野外氮沉降模拟控制实验多采用单一氮肥类型(如NH4NO3或尿素),鲜有研究区分NO3-和NH4+输入对土壤N2O排放的不同影响。
北方森林(Boreal forests)是仅次于热带森林的第二大森林群区,占全球陆地面积的14.5%,其土壤碳密度平均为296 t C/hm2[17];此外,由于该区温度较低,土壤氮素矿化缓慢,土壤有效氮极其匮乏,对外源性氮素响应十分敏感[18]。研究表明,外源性氮素输入会显著改变北方森林植物和微生物群落组成[19]、土壤碳氮转化与温室气体排放[20]、生态系统生产力和固碳潜力[21]。过去普遍认为,水热条件较好、土壤风化强烈的热带/亚热带森林土壤N2O排放较高,而温度较低、有效氮贫乏的高纬度地区森林土壤N2O排放量可以忽略不计[22-23]。然而,近年来一些研究发现,由于气温升高和氮沉降增加提高了高纬度地区森林、苔原等自然生态系统氮素的可利用性,导致该区土壤也大量排放N2O[24-25]。长期以来对高纬度地区自然植被土壤N2O排放的忽视,可能是导致全球N2O收支研究中诸多不确定性的原因之一[26]。大兴安岭寒温带针叶林是北方森林的南缘,面积占全国森林面积的29%,有关土壤N2O对外源性氮素输入尚未有实验报道。
基于上述分析,本文提出以下两个科学假设:(1)外源性氮素输入会显著增加北方森林土壤有效氮含量,提高硝化细菌和反硝化细菌的底物可利用性,进而促进土壤N2O的产生与排放。(2)鉴于北方森林土壤NH4+-N富集,土壤N2O产生可能以硝化过程为主,施加NH4+对土壤N2O的促进作用高于NO3-。为了验证上述两个假设,本文以大兴安岭寒温带针叶林为研究对象,构建多形态、低剂量的氮沉降模拟控制试验,通过野外监测土壤无机氮含量、土壤N2O通量及相关土壤属性,研究 NH4+、NO3-输入及两者交互作用对土壤氮素有效性和N2O排放通量的影响,明确寒温带针叶林土壤N2O通量的主控因子与环境驱动机制。研究结果有助于深入理解森林土壤N2O排放与土壤氮素转化过程、关键功能微生物群落之间的耦合机理,在提高寒温带针叶林氮素利用率、控制土壤N2O排放、降低区域碳源汇评估的不确定性等方面均具有重要的理论与实践意义。
1 材料与方法 1.1 研究区概况研究区位于内蒙古大兴安岭森林生态系统国家野外科学观测研究站以东的开拉气林场(50°20′—50°30′N,121°45′—122°00′E),属大兴安岭西北坡,海拔826 m。该地区是寒温带半湿润气候,冬季寒冷漫长,夏季凉爽多雨。年均气温-5.4℃,最高温出现在7月,最低温在1月。年降水量450—550 mm,其中60%集中于5—9月。年均日照2594 h,全年地表蒸发量800—1200 mm,无霜期80 d。该区主要物种为兴安落叶松(Larix gmelini)、白桦(Betula platyphylla)、杜鹃(Rhododendron simsii)、杜香(Ledum palustre)、红豆越橘(Vaccinium Vitisidaea)等。研究区的植被类型为杜香-落叶松林,林龄约150a。土壤类型为发育于花岗岩残积物上的棕色针叶林土,土壤腐殖质含量10%—30%,pH值为4.5—6.5。
1.2 试验设计为了模拟NH4+和NO3-两个主要离子的输入,参照大兴安岭地区实际大气氮沉降通量(2.5 kg N hm-2 a-1)。2010年5月设置了NH4Cl、KNO3和NH4NO33种氮肥和对照(Control,0 kg N hm-2 a-1)、低氮(Low N,10 kg N hm-2 a-1)、中氮(Medium N,20 kg N hm-2 a-1)、高氮(High N,40 kg N hm-2 a-1)4种水平的氮肥处理,分别模拟未来大气氮沉降增加4倍、8倍和16倍情景下,北方森林生态系统碳、氮循环关键过程的变化。试验采用裂区设计,施氮水平为主处理,施氮类型为副处理,每个施氮水平处理3次重复。为了降低微地形等环境异质性对实验结果造成影响,每个施氮水平下设置一个对照以增强施氮类型间对比。样方规格为10 m×20 m,两个相邻样方的间隔至少为10 m,共36个样方。将全年的氮沉降量平均分配到生长季(5—10月)各月,于每月月初将各形态氮肥溶于20 mL水中,用喷雾器均匀喷洒于各样方内。
1.3 土壤N2O排放通量监测土壤N2O排放量采用静态箱-气相色谱法测定,测定时段为2013年生长季(5—10月)。在每个样地中分别设置带槽的底座(50 cm×50 cm×10 cm)和盖箱(50 cm×50 cm×20 cm),在测定时,将槽内灌满水,打开风扇的电源,然后小心地把带有温度计和小风扇的盖箱沿槽放入。在40 min 时间段内,每隔10 min用100mL注射器抽取1次气样,同时记录大气温度、箱内温度和地下5 cm的温度值。测定N2O时气相色谱的柱箱温度为55 ℃,检测器ECD的温度为250℃;载气(干空气及高纯H2)流量分别为300 mL/min和50 mL/min,尾吹气(N2)流量为10 mL/min。利用土壤水分仪(TDR200,Spectrum Technologies,USA)测定10 cm土壤体积含水量。用气相色谱仪(7890A,Agilent,USA)分析N2O气体浓度,利用下述公式计算土壤N2O气体通量:
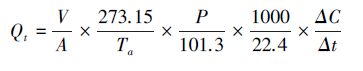
|
(1) |
式中,Qt为t 时刻N2O的排放通量(μg N m-2 h-1);V 为箱体的体积(m3);A 为取样时箱体所覆盖的面积(m2);Ta 为取样时的大气温度(K);P 为取样时的大气压值(kPa);ΔC 为Δt 时间内箱体内N2O浓度增量(ppb);Δt 为时间变化量(s)。
1.4 土壤采集与分析抽气结束后采集有机层和矿质层样品,采样频率为4次/月。就某个处理样方而言,移除未腐解的凋落物层(Oi),沿对角线用方形铁框(10 cm×10 cm×15 cm)和土钻(直径为2.5cm)分别采集有机层(Oae)和矿质层(0—10 cm)土壤样品,同层土壤5个土样均匀混合成一个样品。矿质层土样在田间条件下过 5 mm 筛,用冰盒带回实验室4℃下冷藏。土壤重量含水量采用烘干法测定,在105℃烘干24 h至恒重。土壤无机氮采用比色法测定,准确称取15 g左右的鲜土放入150 mL的塑料瓶中,加入100 mL 2 mol/L KCl溶液浸提,在回旋式振荡器上振荡1h后用定量滤纸过滤,滤液用流动化学分析仪(AA3,SEAL,Germany)测定NH4+-N、NO3--N浓度。
1.5 统计分析利用重复测定方差分析(RANOVA)比较不同施氮水平和施氮类型对土壤温度、含水量、无机氮含量和土壤N2O通量的影响,利用Tukey′s HSD进行均值间的多重比较。采用一元和多元逐步回归分析方法探讨土壤N2O通量与土壤环境因子之间的关系。所有数据利用SPSS 16.0软件进行分析,利用SigmaPlot 12.5软件进行绘图。
2 结果与分析 2.1 土壤温度和水分整个生长季,土壤5cm温度季节变化显著,整体上呈现单峰季节变化(表 1,P < 0.001)。对照处理土壤温度最高值和最低值分别出现在7月初和5月初,平均变化范围为0.70—15.03℃(图 1)。不同施氮处理下,土壤温度变化格局相似,增氮对土壤温度无明显影响(表 1)。
| 变异来源 Source of variance | 土壤温度 Soil temperature | 土壤水分 Soil moisture | 土壤N2O 通量 Soil N2O flux | 土壤NO3--N含量 Soil NO3--N content | 土壤NH4+-N含量 Soil NH4+-N content | |||
| O 层 O layer | M层 M layer | O层 O layer | M层 M layer | |||||
| 组内差异Within subjects (Multivariate) | ||||||||
| 月 份 Month | <0.001 | <0.001 | <0.001 | 0.12 | 0.002 | <0.001 | <0.001 | |
| 月份×施氮水平Month × N level | 0.98 | 0.02 | 0.18 | 0.43 | 0.16 | 0.71 | 0.06 | |
| 月份×施氮类型Month × N form | 0.84 | 0.11 | 0.33 | 0.32 | 0.09 | 0.48 | 0.33 | |
| 组间差异Between subjects | ||||||||
| 施氮水平N level | 0.79 | 0.03 | 0.003 | 0.05 | 0.14 | <0.001 | <0.001 | |
| 施氮类型N form | <0.001 | 0.04 | 0.01 | 0.16 | 0.12 | <0.001 | 0.003 | |

|
| 图 1 土壤温度和水分的季节变化及其对增氮的响应 Fig. 1 The seasonal variations and responses of soil temperature and soil moisture to N addition CK:对照 control;LAC:低氮氯化铵 low-NH4Cl;LPN:低氮硝酸钾 low-KNO3;LAN:低氮硝酸铵 low-NH4NO3;MAC:中氮氯化铵 medium-NH4Cl;MPN:中氮硝酸钾 medium-KNO3;MAN:中氮硝酸铵 medium-NH4NO3;HAC:高氮氯化铵 high-NH4Cl;HPN:高氮硝酸钾 high-KNO3;HAN:高氮硝酸铵 high-NH4NO3 |
整个生长季0—10 cm层土壤含水量季节波动明显,呈逐渐递减的趋势(图 1)。由于5、6月份土壤处于冻融期,土壤含水量较高,5月初对照处理土壤含水量值最高(27.48%)。秋季降水明显减少,8月中旬和9月末土壤含水量较低,最低值为5.86%。就某个月份而言,施氮水平对土壤含水量有显著影响(表 1,P =0.02)。施氮水平和施氮类型均显著改变了土壤体积含水量(表 1,P =0.03,P =0.04)。
2.2 土壤N2O通量整个生长季土壤N2O排放通量季节变化显著(表 1,P < 0.001)。除7月份外,其他月份土壤N2O排放均很低,对照处理土壤N2O通量变化范围为-1.19—5.13 μg N m-2 h-1。施氮后土壤N2O排放急剧增加,7月份出现明显的排放峰(47.77μg N m-2 h-1)(图 2)。施氮水平和施氮类型均对土壤N2O有极显著的影响(表 1,P =0.003,P =0.01)。随着增氮水平增加,土壤N2O排放量逐渐增加。就施氮类型而言,KNO3和NH4NO3的促进效应显著高于NH4Cl,说明硝态氮比铵态氮肥对土壤N2O排放量的影响更为显著。与对照相比,施加NH4NO3对土壤N2O排放的促进效应最强,不同施氮剂量处理土壤N2O通量的增幅度为442%—677%。

|
| 图 2 土壤N2O通量的季节变化及其对增氮的响应 Fig. 2 The seasonal variations and responses of soil N2O fluxes to N addition |
整个生长季有机层土壤NO3--N含量季节波动不大(表 1,P =0.12)。5月份有机层土壤NO3--N无明显积累,只在6月末和8月末出现两个峰值,且含量较低,整个生长季对照处理土壤NO3--N含量变化范围为0—1.56 mg/kg(图 3)。矿质层土壤NO3--N含量高于有机层,变化范围为0.21—9.09 mg/kg。施氮水平显著影响土壤有机层NO3--N含量(表 1,P =0.05),高剂量的氮输入累积效应更加明显(图 3)。施氮水平和施氮类型在P =0.1水平上边缘显著影响矿质层土壤NO3--N含量,低剂量施氮累积效应高于中、高剂量氮输入(图 3)。就某个月份而言,氮素类型也边缘显著影响土壤NO3--N含量(表 1,P =0.09)。

|
| 图 3 有机层、矿质层土壤无机氮含量的月变化及其对增氮的响应 Fig. 3 The monthly variations and responses of soil inorganic N contents in the organic and mineral layers to N addition |
整个生长季有机层和矿质层土壤NH4+-N含量季节变化显著(表 1,P < 0.001)。对照处理矿质层土壤NH4+-N含量的变化范围为5.4—21.84 mg/kg,有机层土壤NH4+-N含量更高,约为矿质层NH4+-N含量的15倍之多,最高值(443.02 mg/kg)和最低值(76.91 mg/kg)分别出现在5月末和8月中旬(图 3)。施氮水平和施氮类型极显著影响有机层土壤NH4+-N含量(表 1,P < 0.001)。与对照相比,有机层土壤NH4+-N含量整体上呈现积累趋势,而矿质层土壤只有高剂量NH4Cl和NH4NO3处理样方积累显著(图 3)。不同剂量的NH4NO3处理对土壤NH4+-N含量的增幅介于22%—49%之间。
2.4 土壤N2O通量与土壤变量之间的关系土壤N2O排放通量总体上与土壤温度的变化一致,呈现明显的单峰季节变化。土壤N2O通量与土壤温度、凋落物层NH4+-N含量正相关,而与其他环境因子相关性不显著(图 4);多元回归分析结果显示,土壤N2O通量主要受温度驱动,其次是凋落物层NH4+-N含量,两者能够解释其季节变异的27%(表 2),说明其他环境因子如土壤质地、有机质含量、pH、Eh含量对土壤N2O通量也会有显著的影响。将施氮水平作为分类变量,发现随着施氮水平的增加土壤N2O通量呈现增加的趋势。

|
| 图 4 土壤N2O通量与土壤变量之间的关系 Fig. 4 Relationships between soil N2O fluxes and soil variables |
| 土壤变量 Soil variables | 回归方程 Equation | 决定系数 R2 | P |
| 土壤温度 Soil temperature(Ts) | FN2O =0.68+0.37Ts | 0.04 | 0.0012 |
| 有机层NH4+-N含量 NH4+-N content in O layer (NH4+-NO) | FN2O =1.24+0.01NH4+-NO | 0.04 | 0.0053 |
| 多元回归 Multiple regression | FN2O=-7.51+0.47Ts+0.011NH4+-NO+0.85LN+2.37MN+2.86HN | 0.27 | <0.001 |
| Ts:土壤温度 soil temperature;NH4+-NO:有机层NH4+-N含量 NH4+-N content in O layer;LN:低氮处理 low N;MN:中氮处理 medium N;HN:高氮处理 high N | |||
土壤NH4+-N取决于氮的矿化、硝化、植物吸收、微生物固持、土壤有机质及矿物质吸附等多个过程的平衡4[27]。本研究发现施氮类型和剂量显著改变土壤NH+-N含量,且土壤NH4+-N的积累效应较NO3--N更加显著(表 2和图 3),该结果验证了本文第一个科学假设(即施氮增加土壤氮素有效性)。总体上,施氮一致导致有机层(O层)NH4+-N累积,但是只有施加铵态氮肥(NH4Cl和NH4NO3)时矿质层土壤NH4+-N累积才显著,该结果说明施氮引起有机氮的矿化大于NH4+的消耗,这与许多贫氮的北方森林、温带森林的结果相似4[28-29]。本研究模拟氮沉降输入采用地表喷洒方式,一部分外源性NH+首先被凋落物层有机质吸附;其次,大兴安岭地区植物和微生物均是喜铵的,植物优先利用NH4+-N 4[30]。Xu 等[31]利用15N示踪技术探讨不同纬度森林主要乔木对有效氮源(NH+、NO3-和氨基酸)的吸收利用格局,也发现植物优先吸收NH4+-N。Sheng等4[32]研究外源性氮素在南北森林中的运移规律时发现,北方森林中(15NH4)2SO4吸收速率明显高于K15NO3。在长时间尺度上,植物是氮素竞争的优势者[33],植物对NH+的选择性吸收也导致低氮处理矿质层土壤NH4+-N无明显累积;当施加的NH4+以及矿化的NH4+超过土壤物理化学和生物的固持容量,多余的NH4+才能在矿质层发生积累。该研究结果表明,如果将大兴安岭北方森林大气氮沉降背景值(2.5 kg N hm4-2 a-1)考虑在内,导致土壤NH+-N发生明显积累的大气氮沉降临界通量约为22.5 kg N hm-2a-1。
土壤NO3--N含量是土壤硝化、植物吸收、反硝化及液态淋溶等多个过程的净效应。施氮只边缘显著地改变O层和M层土壤NO3--N含量,低氮处理样方矿质层土壤NO3--N累积明显,研究结果表明施氮3a大兴安岭寒温带针叶林土壤NO3--N含量处于一种动态平衡状态(产生=损耗)。施加KNO3和NH4NO3直接增加土壤NO3--N含量,施加铵态氮刺激硝化细菌活性,增加土壤NO3--N含量。但是,本研究发现3a施氮并没有导致寒温带针叶林土壤NO3--N明显的累积,这与热带、亚热带森林增氮控制实验的结果不一致3[34-35],也远低于全球Meta分析的结果(NO--N淋溶增加461%)3[36]。造成上述格局的可能原因是:(1)施氮剂量较低,外源性氮输入主要被植物和微生物同化[32]。(2) 大兴安岭寒温带针叶林土壤为典型棕色针叶林土,质地较粗,砾石、砂粒、粉粒和粘粒含量分别为11.16%、51.76%、27.55%、9.53%[37],NO--N垂直渗漏明显,难以在表层积累。(3)寒温带针叶林土壤水分含量较高,冻融过程强烈,反硝化氮素损失明显[38]。许多研究表明,反硝化过程支配着森林土壤N2O产生过程[39-40],外源性氮输入直接或间接地增加反硝化细菌群落的底物,进而增加土壤氮素的气态排放(NO、N2O和N2)。本研究发现施氮显著促进土壤N2O排放也支持上述推论。
3.2 施氮类型和剂量对土壤N2O通量的影响自然状态下,寒温带针叶林土壤N2O年累计排放量为(1.23±0.42)kg N hm4-2 a-1,与傅民杰等[38]的观测结果相近。施氮一致增加寒温带针叶林土壤N2O排放,NH4NO3处理增幅为442%—677%,显著高于全球施氮试验的平均水平(134%)[36],说明贫氮的寒温带针叶林土壤N2O排放对外源性氮素输入十分敏感,应该倍加重视。此外,高剂量的硝态氮肥(KNO3和NH4NO3)输入的促进效应高于铵态氮肥,且NH4NO3增幅最大。研究结果表明,寒温带针叶林土壤N2O主要来源于硝化和反硝化过程,外源性NH+和NO3-对N2O排放的影响具有加和效应,并且土壤N2O产生来源于反硝化过程的比例更大,这与假设2相反。本研究发现,寒温带针叶林土壤N2O通量主要受温度驱动,其次是有机层NH4+-N,并且随着施氮水平的增加而增加(图 4和表 3)。温度主要控制 N2O 产生过程中的酶活性,在土壤水分、反应基质充足的条件下,土壤N2O排放与温度呈现显著的正相关4[41]。虽然土壤产生N2O的生物代谢过程主要由温度控制,但N2O排放并非完全取决于其产生速率,其排放过程还会受到底物有效性的影响。由于寒温带针叶林土壤无机氮主要以NH+-N为主,且积累在凋落物层,导致凋落物层NH4+-N转化过程支配着土壤N2O的排放。目前关于森林土壤N2O产生存在硝化和反硝化为主之争论4[14],本研究只是间接得出NH+-N转化效率高于NO3--N;下一步研究需要结合15N示踪技术和分子生物学方法,精细测定多个氮素转化过程速率以及介导氨化、硝化和反硝化过程的功能微生物群落丰度和组成,量化硝化和反硝化过程对N2O产生的相对贡献。
4 结论本研究以大兴安岭寒温带针叶林为研究对象,构建3种氮肥、4个施氮剂量的增氮控制试验,测定生长季土壤无机氮含量、土壤N2O通量及水热环境因子,分析施氮类型与水平对土壤氮素可利用性及土壤N2O排放的影响,探讨氮素富集条件下寒温带针叶林土壤N2O排放的环境驱动机制。得出以下3个研究结论:(1)施氮类型和剂量显著改变土壤无机氮含量,土壤NH4+-N的积累效应较NO3--N更加显著,大气氮沉降输入短期内(<3a)不会导致土壤氮素的大量流失。(2)施氮一致增加寒温带针叶林土壤N2O排放,增幅高于全球平均水平,外源性NH4+和NO3-输入对N2O排放的影响具有加和效应。由于N2O产生与消耗过程极其复杂,本文只探讨了水热因子和底物浓度对土壤N2O通量的影响,未能量化硝化和反硝化过程对土壤N2O产生的相对贡献。未来研究应从生物化学和微生物分子生态学角度出发,结合15N示踪技术和分子生物学方法,深入探讨氮素富集条件下土壤关键微生物功能群基因丰度、组成及其与氮素转化过程的耦合关系。
| [1] | Stocker T F, Qin D H, Plattner G K, Tignor M B, Allen S K, Boschung J, Nauels A, Yu X, Bex V, Midgley P M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2013 : 1 –1535. |
| [2] | Ravishankara A R, Daniel J S, Portmann R W. Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. Science , 2009, 326 (5949) : 123–125. DOI:10.1126/science.1176985 |
| [3] | 蔡祖聪, 徐华, 马静. 稻田生态系统CH4和N2O排放. 合肥: 中国科学技术大学出版社, 2009 : 1 –375. |
| [4] | Lamarque J F, Kiehl J T, Brasseur G P, Butler T, Cameron-Smith P, Collins W D, Collins W J, Granier C, Hauglustaine D, Hess P G, Holland E A, Horowitz L, Lawrence M G, McKenna D, Merilees P, Prather M J, Rasch P J, Rotman D, Shindell D, Thornton P. Assessing future nitrogen deposition and carbon cycle feedback using a multimodel approach: Analysis of nitrogen deposition. Journal of Geophysical Research , 2005, 110 (D19) . |
| [5] | Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science , 2008, 320 (5878) : 889–892. DOI:10.1126/science.1136674 |
| [6] | 王汝南. 模拟大气氮沉降对温带森林土壤温室气体交换通量的影响[D]. 北京: 北京林业大学, 2012. |
| [7] | Butterbach-Bahl K, Gasche R, Willibald G, Papen H. Exchange of N-gases at the Höglwald Forest-A summary. Plant and Soil , 2002, 240 (1) : 117–123. DOI:10.1023/A:1015825615309 |
| [8] | Baggs E M. A review of stable isotope techniques for N2O source partitioning in soils: recent progress, remaining challenges and future considerations. Rapid Communications in Mass Spectrometry , 2008, 22 (11) : 1664–1672. DOI:10.1002/(ISSN)1097-0231 |
| [9] | Nemergut D R, Townsend A R, Sattin S R, Freeman K R, Fierer N, Neff J C, Bowman W D, Schadt C W, Weintraub M N, Schmidt S K. The effects of chronic nitrogen fertilization on alpine tundra soil microbial communities: implications for carbon and nitrogen cycling. Environmental Microbiology , 2008, 10 (11) : 3093–3105. DOI:10.1111/emi.2008.10.issue-11 |
| [10] | Ernfors M, Rütting T, Klemedtsson L. Increased nitrous oxide emissions from a drained organic forest soil after exclusion of ectomycorrhizal mycelia. Plant and Soil , 2011, 343 (1/2) : 161–170. |
| [11] | Zhu T B, Meng T Z, Zhang J B, Yin Y F, Cai Z C, Yang W Y, Zhong W H. Nitrogen mineralization, immobilization turnover, heterotrophic nitrification, and microbial groups in acid forest soils of subtropical China. Biology and Fertility of Soils , 2013, 49 (3) : 323–331. DOI:10.1007/s00374-012-0725-y |
| [12] | Corre M D, Veldkamp E, Arnold J, Wright S J. Impact of elevated N input on soil N cycling and losses in old-growth lowland and montane forests in Panama. Ecology , 2010, 91 (6) : 1715–1729. DOI:10.1890/09-0274.1 |
| [13] | Fenn M E, Poth M A, Terry J D, Blubaugh T J. Nitrogen mineralization and nitrification in a mixed-conifer forest in southern California: controlling factors, fluxes, and nitrogen fertilization response at a high and low nitrogen deposition site. Canadian Journal of Forest Research , 2005, 35 (6) : 1464–1486. DOI:10.1139/x05-068 |
| [14] | 方华军, 程淑兰, 于贵瑞, 王永生, 徐敏杰, 党旭升, 李林森, 王磊, 李晓玉, 司高月. 森林土壤氧化亚氮排放对大气氮沉降增加的响应研究进展. 土壤学报 , 2015, 52 (2) : 262–271. |
| [15] | Zhang W, Mo J M, Yu G R, Fang Y T, Li D J, Lu X K, Wang H. Emissions of nitrous oxide from three tropical forests in Southern China in response to simulated nitrogen deposition. Plant and Soil , 2008, 306 (1/2) : 221–236. |
| [16] | Wei D, Xu-Ri, Liu Y W, Wang Y H, Wang Y S. Three-year study of CO2 efflux and CH4/N2O fluxes at an alpine steppe site on the central Tibetan Plateau and their responses to simulated N deposition. Geoderma , 2014, 232-234 : 88–86. DOI:10.1016/j.geoderma.2014.05.002 |
| [17] | Lal R. Forest soils and carbon sequestration. Forest Ecology and Management , 2005, 220 (1/3) : 242–258. |
| [18] | Luyssaert S, Schulze E D, Börner A, Knohl A, Hessenmöller D, Law B E, Ciais P, Grace J. Old-growth forests as global carbon sinks. Nature , 2008, 455 (7210) : 213–215. DOI:10.1038/nature07276 |
| [19] | Allison S D, Hanson C A, Treseder K K. Nitrogen fertilization reduces diversity and alters community structure of active fungi in boreal ecosystems. Soil Biology and Biochemistry , 2007, 39 (8) : 1878–1887. DOI:10.1016/j.soilbio.2007.02.001 |
| [20] | Maljanen M, Jokinen H, Saari A, Strömmer R, Martikainen P J. Methane and nitrous oxide fluxes, and carbon dioxide production in boreal forest soil fertilized with wood ash and nitrogen. Soil Use and Management , 2006, 22 (2) : 151–157. DOI:10.1111/sum.2006.22.issue-2 |
| [21] | Magnani F, Mencuccini M, Borghetti M, Berbigier P, Berninger F, Delzon S, Grelle A, Hari P, Jarvis P G, Kolari P, Kowalski A S, Lankreijer H, Law B E, Lindroth A, Loustau D, Manca G, Moncrieff J B, Rayment M, Tedeschi V, Valentini R, Grace J. The human footprint in the carbon cycle of temperate and boreal forests. Nature , 2007, 447 (7146) : 849–851. DOI:10.1038/nature05847 |
| [22] | Werner C, Butterbach-Bahl K, Haas E, Hickler T, Kiese R. A global inventory of N2O emissions from tropical rainforest soils using a detailed biogeochemical model. Global Biogeochemical Cycles , 2007, 21 (3) : GB3010. |
| [23] | Koehler B, Corre M D, Veldkamp E, Wullaert H, Wright S J. Immediate and long-term nitrogen oxide emissions from tropical forest soils exposed to elevated nitrogen input. Global Change Biology , 2009, 15 (8) : 2049–2066. DOI:10.1111/gcb.2009.15.issue-8 |
| [24] | Repo M E, Susiluoto S, Lind S E, Jokinen S, Elsakov V, Biasi C, Virtanen T, Martikainen P J. Large N2O emissions from cryoturbated peat soil in tundra. Nature Geoscience , 2009, 2 (3) : 189–192. DOI:10.1038/ngeo434 |
| [25] | Elberling B, Christiansen H H, Hansen B U. High nitrous oxide production from thawing permafrost. Nature Geoscience , 2010, 3 (7) : 506–506. DOI:10.1038/ngeo893 |
| [26] | Zaehle S, Dalmonech D. Carbon-nitrogen interactions on land at global scales: current understanding in modelling climate biosphere feedbacks. Current Opinion in Environmental Sustainability , 2011, 3 (5) : 311–320. DOI:10.1016/j.cosust.2011.08.008 |
| [27] | 方华军, 程淑兰, 于贵瑞. 森林土壤碳、氮淋失过程及其形成机制研究进展. 地理科学进展 , 2007, 26 (3) : 29–37. |
| [28] | 胡艳玲, 韩士杰, 李雪峰, 赵玉涛, 李东. 长白山原始林和次生林土壤有效氮含量对模拟氮沉降的响应. 东北林业大学学报 , 2009, 37 (5) : 36–38. |
| [29] | Gao W L, Cheng S L, Fang H J, Chen Y, Yu G R, Zhou M, Zhang P L, Xu M J. Effects of simulated atmospheric nitrogen deposition on inorganic nitrogen content and acidification in a cold-temperate coniferous forest soil. Acta Ecologica Sinica , 2013, 33 (2) : 114–121. DOI:10.1016/j.chnaes.2013.01.008 |
| [30] | 崔晓阳, 宋金凤. 原始森林土壤NH4+/NO3-生境特征与某些针叶树种的适应性. 生态学报 , 2005, 25 (11) : 3082–3092. |
| [31] | Xu X L, Li Q K, Wang J Y, Zhang L M, Tian S N, Zhi L, Li Q R, Sun Y. Inorganic and organic nitrogen acquisition by a fern Dicranopteris dichotoma in a subtropical forest in South China. PLoS One , 2014, 9 (5) : e90075. DOI:10.1371/journal.pone.0090075 |
| [32] | Sheng W P, Yu G R, Fang H J, Jiang C M, Yan J H, Zhou M. Sinks for inorganic nitrogen deposition in forest ecosystems with low and high nitrogen deposition in China. PLoS One , 2014, 9 (2) : e89322. DOI:10.1371/journal.pone.0089322 |
| [33] | Kuzyakov Y, Xu X L. Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytologist , 2013, 198 (3) : 656–669. DOI:10.1111/nph.12235 |
| [34] | Fang H J, Yu G R, Cheng S L, Zhu T H, Zheng J J, Mo J M, Yan J H, Luo Y Q. Nitrogen-15 signals of leaf-litter-soil continuum as a possible indicator of ecosystem nitrogen saturation by forest succession and N loads. Biogeochemistry , 2011, 102 (1/3) : 251–263. |
| [35] | Wang Y S, Cheng S L, Fang H J, Yu G R, Xu M J, Dang X S, Li L S, Wang L. Simulated nitrogen deposition reduces CH4 uptake and increases N2O emission from a subtropical plantation forest soil in southern China. PLoS One , 2014, 9 (4) . |
| [36] | Lu M, Yang Y H, Luo Y Q, Fang C M, Zhou X H, Chen J K, Yang X, Li B. Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis. New Phytologist , 2011, 189 (4) : 1040–1050. DOI:10.1111/nph.2011.189.issue-4 |
| [37] | Fang H J, Yu G R, Cheng S L, Zhu T L, Wang Y S, Yan J H, Wang M, Cao M, Zhou M. Effects of multiple environmental factors on CO2 emission and CH4 uptake from old-growth forest soils. Biogeosciences , 2010, 7 (1) : 395–407. DOI:10.5194/bg-7-395-2010 |
| [38] | 傅民杰, 王传宽, 王颖, 刘实, 丁爽. 气候暖化对解冻期不同纬度兴安落叶松林土壤氧化亚氮释放的影响. 应用生态学报 , 2009, 20 (7) : 1635–1642. |
| [39] | Zhang J B, Cai Z C, Zhu T B. N2O production pathways in the subtropical acid forest soils in China. Environmental Research , 2011, 111 (5) : 643–649. DOI:10.1016/j.envres.2011.04.005 |
| [40] | Zhang J B, Cai Z C, Zhu T B, Yang W Y, Müller C. Mechanisms for the retention of inorganic N in acidic forest soils of southern China. Scientific Reports , 2013, 3 : 2342. |
| [41] | Livesley S J, Grover S, Hutley L B, Jamali H, Butterbach-Bahl K, Fest B, Beringer J, Arndt S K. Seasonal variation and fire effects on CH4, N2O and CO2 exchange in savanna soils of northern Australia. Agricultural and Forest Meteorology , 2011, 151 (11) : 1440–1452. DOI:10.1016/j.agrformet.2011.02.001 |