 2017, Vol. 37
2017, Vol. 37文章信息
- 邵倩文, 刘镇盛, 章菁, 孙栋, 林施泉
- SHAO Qianwen, LIU Zhensheng, ZHANG Jing, SUN Dong, LIN Shiquan.
- 长江口及邻近海域浮游动物群落结构及季节变化
- Seasonal variation in zooplankton community structure in the Changjiang Estuary and its adjacent waters
- 生态学报[J]. 2017, 37(2): 683-691
- Acta Ecologica Sinica[J]. 2017, 37(2): 683-691
- http://dx.doi.org/10.5846/stxb201507271574
-
文章历史
- 收稿日期: 2015-07-27
- 网络出版日期: 2016-06-13





2. 国家海洋局海洋生态系统与生物地球化学重点实验室, 杭州 310012
2. Key Laboratory of Marine Ecosystem and Biogeochemistry, State Oceanic Administration, Hangzhou 310012, China
在典型的温带和亚热带地区,浮游动物是河口和近海生态系统的重要组成部分。浮游动物通过摄食浮游植物而影响初级生产,同时,又被更高营养阶层的动物(鱼、虾、鲸、海鸟等)捕食,其群落结构、种群动态和多样性影响许多鱼类和其他海洋动物资源的生物量,因而浮游动物在海洋食物网中起关键作用[1-2]。同时,河口和冲淡水扩散区常受到陆架水系物理和化学特征的调控,这些复杂因素又影响着浮游动物群落结构及其分布[3]。长江是我国最大的河流,水量丰沛,其平均年入海径流量约9.28×1011 m3,占输入东海总径流量的84.4%。长江冲淡水有明显的季节变化,5—10月份为丰水期,占全年的71.7%,11—4月份为枯水期,占全年的28.3%。在长江河口河水与海水经常交汇、径流与潮流相互抗衡的地带发育成最大浑浊带,最大浑浊带对河口泥沙的聚集和沉降起着十分重要的作用[4]。长江口及邻近海域主要受长江冲淡水、台湾暖流、浙江近岸流和黄海冷水团等水系共同影响,水文环境复杂多变,浮游动物群落在不同水域存在明显差异。
从1980年以来,对长江口及其邻近海域浮游动物的研究已有不少报道[5-13],但是中尺度调查研究相对匮乏。首先,研究海域的范围主要集中在124°E以西近岸河口区或东海区海域,迄今缺少从长江内河段(120°E)开始延伸至长江冲淡水扩散海域(127°E)中尺度海域的研究报道。其次,针对长江口及其邻近海域浮游动物群落的类群组成及四个季节变化的系统报道甚少。本文采用经典的形态学分类方法,结合同步观测的其他环境因子调查结果,对长江口及其邻近海域(包括长江内河段及长江冲淡水扩散海域)浮游动物群落结构及季节变化等方面进行研究,旨在揭示长江口及其邻近海域浮游动物群落结构时空变化特征,并且探讨浮游动物群落的季节变动与环境因子的关系,为长江口海域资源的可持续利用和海洋生态系统研究提供基础资料和科学依据。
1 材料和方法 1.1 调查区域、采样及分析方法2006—2007年,在长江口及其邻近海域29°30'N—32°30'N,120°00'E—127°30'E(图 1)开展春季(2007年4月—5月)、夏季(2006年7月—8月)、秋季(2007年10月—12月)和冬季(2006年12月—2007年2月)4个航次的海洋生物生态调查。海上调查由“中国海监49船”执行,调查海域共布设150个观测站(图 1)。

|
| 图 1 长江口及其邻近海域采样站位图 Fig. 1 Sampling stations in Changjiang Estuary and adjacent areas |
浮游动物现场采样中,使用装有进口流量计(德国HYDRO-BIOS公司-生物网口流量计E-Flow)的浅水I型浮游生物网(网长145 cm,网口内径50 cm,筛绢孔径0.505 mm,采集水深<30 m的浮游动物样品)或大型浮游生物网(网长280 cm,网口内径80 cm,筛绢孔径0.505 mm,采集水深>30 m的浮游动物样品)进行垂直拖网。海域水深<200 m时,从底层至表层进行垂直拖网,水深>200 m时,从200 m至表层垂直拖网采集样品。将每个测站采集的浮游动物样品装入600 cm3的塑料瓶中,体积分数为加入5%甲醛溶液固定保存。采集样品带回实验室后,剔除样品中的杂物,利用电子天平(Sartorius CP124S)称量浮游动物生物量,将称量完成后的样品置于体视显微镜下进行浮游动物的种属鉴定和个体计数。海上采样及实验室分析方法均按《海洋生物生态调查技术规程》[14]的规定执行。
同步观测的参数有水温、盐度、悬浮物浓度和叶绿素a浓度。海水温度、盐度使用SYA2-2进行现场测定。以上参数采样和分析均按《海洋生物生态调查技术规程》[14]、《海洋化学调查技术规程》[15]和《海洋水文气象调查技术规程》[16]的相关规定实施。
1.2 数据处理本研究采用Surfer 11.0绘图软件绘制调查站位图和浮游动物种类数平面分布图,使用SPSS 20.0软件对浮游动物丰度和环境因子进行Pearson相关分析。本研究的环境因子主要包括表层温度、表层盐度、表层悬浮物浓度、底层温度、底层盐度、底层悬浮物浓度、叶绿素a浓度。
浮游动物的优势种根据每个物种的优势度(Y)来确定[17]:
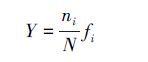
|
式中,ni为第i种的个体数,N为所有种类总个体数,fi为出现频率。Y值大于0.02的种类为优势种。
2 结果 2.1 长江口及其邻近海域浮游动物群落结构及季节变化长江口及其邻近海域四季共出现浮游动物460种,隶属7个门,15纲,36目,133科,246属,此外,还有54类浮游幼体(表 1)。其中,桡足类是浮游动物最优势的类群,有193种,占浮游动物总种类数的41.96%,端足类为第二优势类群,有51种,占11.09%,水螅水母为第三优势类群,有34种,占7.39%。其它类群包括:十足类26种,管水母、多毛类、枝角类各25种,毛颚动物15种,腹足类14种,糠虾类12种,涟虫类9种,介形类8种,栉板动物5种,海樽类6种,磷虾类和有尾类各4种,钵水母3种,等足类1种。
| 门类 Community |
类群 Groups |
春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | 全年Year | |||||
| 种数 Number |
/% | 种数 Number |
/% | 种数 Number |
/% | 种数 Number |
/% | 种数 Number |
/% | ||
| 刺胞动物Cnidaria | 水螅水母Hydromedusae | 19 | 8.26 | 29 | 9.15 | 18 | 5.83 | 8 | 5.80 | 34 | 7.39 |
| 管水母Siphonophora | 20 | 8.70 | 18 | 5.68 | 19 | 6.15 | 4 | 2.90 | 25 | 5.43 | |
| 钵水母Scyphomedusae | 1 | 0.43 | 2 | 0.63 | 1 | 0.32 | - | - | 3 | 0.65 | |
| 栉板动物Ctenophora | 栉水母Ctenophora | 3 | 1.30 | 5 | 1.58 | 2 | 0.65 | 1 | 0.72 | 5 | 1.09 |
| 环节动物Annelida | 多毛类Polychaeta | 10 | 4.35 | 16 | 5.05 | 15 | 4.85 | 1 | 0.72 | 25 | 5.43 |
| 软体动物Mollusca | 腹足纲Gastropoda | 4 | 1.74 | 13 | 4.10 | 10 | 3.24 | 4 | 2.90 | 14 | 3.04 |
| 节肢动物Arthropoda | 枝角类Cladocera | 13 | 5.65 | 16 | 5.05 | 10 | 3.24 | 4 | 2.90 | 25 | 5.43 |
| 桡足类Copepoda | 103 | 44.78 | 142 | 44.79 | 135 | 43.69 | 69 | 50.00 | 193 | 41.96 | |
| 介形类Ostracoda | 2 | 0.87 | 2 | 0.63 | 8 | 2.59 | 2 | 1.45 | 8 | 1.74 | |
| 糠虾类Mysida | 3 | 1.30 | 4 | 1.26 | 10 | 3.24 | 4 | 2.90 | 12 | 2.61 | |
| 端足类Amphipoda | 18 | 7.83 | 23 | 7.26 | 38 | 12.30 | 13 | 9.42 | 51 | 11.09 | |
| 涟虫类Cumacea | 4 | 1.74 | 4 | 1.26 | 7 | 2.27 | 4 | 2.90 | 9 | 1.96 | |
| 等足类Isopoda | - | - | - | - | 1 | 0.32 | - | - | 1 | 0.22 | |
| 磷虾类Euphausiacea | 4 | 1.74 | 3 | 0.95 | 3 | 0.97 | 3 | 2.17 | 4 | 0.87 | |
| 十足类Decapoda | 11 | 4.78 | 19 | 5.99 | 15 | 4.85 | 8 | 5.80 | 26 | 5.65 | |
| 毛颚动物Chaetognatha | 箭虫纲Sagittoidea | 7 | 3.04 | 12 | 3.79 | 11 | 3.56 | 8 | 5.80 | 15 | 3.26 |
| 尾索动物Appendicularia | 有尾纲Sagittoidea | 3 | 1.30 | 3 | 0.95 | 3 | 0.97 | 4 | 2.90 | 4 | 0.87 |
| 海樽纲Thaliacea | 5 | 2.17 | 6 | 1.89 | 3 | 0.97 | 1 | 0.72 | 6 | 1.30 | |
| 合计Total | 230 | 317 | 309 | 138 | 460 | ||||||
| 浮游幼体Pelagic larva | 27 | 43 | 28 | 21 | 54 | ||||||
浮游动物群落种类组成有明显季节变化,夏季(317种)是浮游动物物种最丰富的季节,秋季(309种)高于春季(230种),冬季(138种)种类数最低。春季,长江口海域共出现浮游动物17类,230种,占全年出现浮游动物种类数的50.00%,另有浮游幼体27类。其中,长江口门区、舟山群岛东南部和东海东北部海域浮游动物种类丰富,形成3个高值区(>40种)(图 2)。夏季,随着水温的上升,浮游动物种类明显增多,共出现浮游动物17类,317种,为长江口海域浮游动物四季种类数的峰值,占全年出现浮游动物种类数的68.91%,另有浮游幼体43类。种类数高值区位于长江口门区、舟山群岛东部和东海东北部陆架海域,大部分水域种类数大于65种。种类数低值区位于杭州湾和长江口内河段,浮游动物种类数少于20种。夏季从近岸往外海、调查海域北部往南部,浮游动物种类数分布呈递增趋势(图 2)。秋季,长江口海域共出现浮游动物18类,309种,占全年出现浮游动物种类数的67.17%,另有浮游幼体28类。浮游动物种类数维持在高水平,其中长江口门区和东海东北部陆架海域形成2个高值区(>70种)(图 2)。冬季,长江口海域共出现浮游动物16类,138种,占全年出现浮游动物种类数的30.00%,另有浮游幼体21类。长江口门区海域浮游动物种类相对丰富,形成高值区(>20种)(图 2)。总体上,长江口及其邻近海域浮游动物种类数外海高于近海,南部高于北部。

|
| 图 2 长江口及其邻近海域浮游动物种类数分布和季节变化 Fig. 2 Seasonal variation and distribution of zooplankton species number in Changjiang Estuary and its adjacent waters |
在河口和近岸水域,环境变量和人类活动因素影响着浮游动物群落的分布和时空变化,陆源径流携带来丰富生源物质,引起近岸水体营养盐浓度的变化,从而改变浮游动物物种多样性和优势种演替[18]。根据浮游动物对温度、盐度和环境的适应性,及其生态习性和分布规律,参考《海洋浮游生物学》等专著和文献[19-23],本文将长江口及其邻近海域浮游动物大体分为5种生态类群。
(1) 近岸低盐类群
该浮游动物类群种类繁多,主要分布于近岸低盐水域和长江口内河段,是长江口海域主要的生态类群,代表种群有真刺唇角水蚤Labidocera euchaeta、针刺拟哲水蚤Paracalanus aculeatus和太平洋纺锤水蚤Acartia pacifica等。在该类群中,有少数种类属河口低盐性种和淡水种,如:火腿伪镖水蚤Pseudodiaptomus poplesia、虫肢歪水蚤Tortanus vermiculus等。
(2) 广温广盐类群
该类群在长江口海域浮游动物数量上占优势,在长江口水系交汇混合区广泛分布,4个不同季节均有出现。常见种有中华哲水蚤Calanus sinicus、亚强次真哲水蚤Subeucalanus subcrassus和百陶带箭虫Zonosagitta bedoti等。
(3) 低温高盐类群
这一类群的浮游动物种类和数量少,出现频率较低,主要出现在中层及深层水域,随着深层水的涌升和黄海冷水团的作用出现在东海区。代表种有芦氏拟真刺水蚤Paraeuchaeta russelli和太平洋磷虾Euphausia pacifica等。
(4) 高温广盐类群
这一类群浮游动物数量少,但分布广,常出现在温度较高的夏季和秋季。代表种有孔雀丽哲水蚤Calocalanus pavo和左突唇角水蚤Labidocera sinilobata等。
(5) 高温高盐类群
这是一类适温、适盐性较高的浮游动物,广泛分布于受台湾暖流和黑潮影响的水域,该类群种类丰富。代表种有精致真刺水蚤Euchaeta concinna、平滑真刺水蚤Euchaeta plana和肥胖软箭虫Flaccisagitta enflata等。
2.3 浮游动物优势种的季节变化长江口及其邻近海域的浮游动物优势种呈现出明显的季节更替现象。中华哲水蚤和百陶带箭虫为长江口及其邻近海域四季优势种(表 2)。春季,优势种有中华哲水蚤、针刺拟哲水蚤、百陶带箭虫。其中,中华哲水蚤平均丰度达(158.6±425.9)个/m3,优势度为0.399。夏季,随着水温升高,长江径流量增大,外海暖水种进入研究海域,浮游动物物种多样性上升,优势种集中度明显下降,背针胸刺水蚤成为该海域第一优势种,优势度为0.105。秋季,浮游动物优势种的集中度下降为四季的最低值,亚强次真哲水蚤成为第一优势种,但其平均丰度仅为(17.7±45.1)个/m3,其它优势种的平均丰度也不高。冬季,浮游动物优势种的集中度逐渐回升,但丰度明显下降,第一优势种中华哲水蚤的优势度为0.253,平均丰度仅为(4.8±17.2)个/m3。
| 季节Season | 优势种(优势度Y)Dominant species (Dominance Y) |
| 春季Spring | 中华哲水蚤(0.399)、针刺拟哲水蚤(0.028)、百陶带箭虫(0.026) |
| 夏季Summer | 背针胸刺水蚤(0.105)、太平洋纺锤水蚤(0.077)、中华哲水蚤(0.058)、百陶带箭虫(0.040)、针刺拟哲水蚤(0.037)、精致真刺水蚤(0.033)、平滑真刺水蚤(0.024)、肥胖软箭虫(0.024)、真刺唇角水蚤(0.022) |
| 秋季Autumn | 亚强次真哲水蚤(0.079)、百陶带箭虫(0.068)、平滑真刺水蚤(0.055)、芦氏拟真刺水蚤(0.037)、中华哲水蚤(0.032)、肥胖软箭虫(0.024) |
| 冬季Winter | 中华哲水蚤(0.253)、真刺唇角水蚤(0.047)、百陶带箭虫(0.041)、捷氏歪水蚤(0.038)、精致真刺水蚤(0.028) |
长江口及其邻近海域环境因子有明显季节变化(表 3)。水温的季节变化特征为:夏季>秋季>春季>冬季,夏季底层水温明显低于表层水温,其它季节表层水温和底层水温相近;表层盐度季节变化特征为:冬季>春季>秋季>夏季,底层盐度季节变化特征为:春季>冬季>夏季>秋季,其中,春季和夏季表层平均盐度明显高于底层平均盐度;表层悬浮物浓度的季节变化为:冬季>春季>秋季>夏季,底层悬浮物浓度的季节变化为:秋季>春季>冬季>夏季,夏季悬浮物平均浓度四季最低,春季和秋季底层悬浮物平均浓度明显高于表层,冬季表层和底层悬浮物平均浓度差别不大;叶绿素a浓度的季节变化为:夏季>春季>秋季>冬季。
| 环境因子 | 春季 | 夏季 | 秋季 | 冬季 |
| Environmental factors | Spring | Sumer | Autumn | Winter |
| 表层水温surface temperature/℃ | 15.42±1.66 | 27.75±1.63 | 21.01±1.69 | 11.79±3.47 |
| 底层水温bottom temperature/℃ | 15.28±1.77 | 23.52±3.99 | 21.07±1.58 | 11.93±3.47 |
| 表层盐度surface salinity | 24.70±10.88 | 21.07±11.57 | 23.26±11.61 | 25.36±10.88 |
| 底层盐度bottom salinity | 26.17±10.66 | 24.94±12.32 | 24.12±11.89 | 25.71±10.68 |
| 表层悬浮物浓度 Concentration of surface suspended matter/(mg/L) |
370.63±1084.71 | 118.83±237.01 | 273.93±654.32 | 449.52±1035.61 |
| 底层悬浮物浓度 Concentration of bottom suspended matter/(mg/L) |
545.83±1348.64 | 177.54±383.54 | 666.90±1368.52 | 495.75±1037.08 |
| 叶绿素a浓度Concentration of chlorophyll a/(mg/m3) | 2.04±2.23 | 4.82±8.70 | 1.55±3.04 | 0.97±0.98 |
浮游动物丰度与环境因子间的Pearson相关性系数见表 4。春季,浮游动物丰度与温度和盐度呈现极显著正相关(P<0.01),与悬浮物浓度呈现显著负相关(P<0.05);夏季,浮游动物丰度与悬浮物浓度呈现极显著正相关(P<0.01),与表层温度呈现极显著负相关(P<0.05),与底层盐度和叶绿素a浓度呈现显著正相关(P<0.05);秋季,浮游动物丰度与盐度和叶绿素a浓度呈现极显著正相关(P<0.01),与悬浮物浓度呈现极显著负相关(P<0.01);冬季,浮游动物丰度与盐度呈现极显著负相关(P<0.01)。
| 春Spring | 夏Summer | 秋Autumn | 冬Winter | ||||
| 环境 | 相关系数 | 环境 | 相关系数 | 环境 | 相关系数 | 环境 | 相关系数 |
| ST | 0.225** | ST | -0.283** | ST | -0.052 | ST | -0.113 |
| BT | 0.253** | BT | -0.084 | BT | -0.043 | BT | -0.127 |
| SS | 0.218** | SS | 0.134 | SS | 0.608** | SS | -0.234** |
| BS | 0.236** | BS | 0.182* | BS | 0.587** | BS | -0.234** |
| SSM | -0.176* | SSM | 0.323** | SSM | -0.257** | SSM | -0.074 |
| BSM | -0.185* | BSM | 0.316** | BSM | -0.217** | BSM | -0.093 |
| Chl a | 0.015 | Chl a | 0.165* | Chl a | 0.270** | Chl a | -0.035 |
| ST:表层温度 surface temperature;BT:底层温度 bottom temperature;SS:表层盐度 surface salinity;BS:底层盐度 bottom salinity;SSM:表层悬浮物浓度 concentration of surface suspended matter;BSM:底层悬浮浓度物 concentration of bottom suspended matter;Chl a:叶绿素a浓度 concentration of chlorophyll a;**P <0.01;*P <0.05 | |||||||
本研究共鉴定出浮游动物460种,浮游幼体54类。其中,桡足类有193种,占总种类数的41.96%,其次是端足类和水螅水母,分别占总种类数的11.09%和7.39%。桡足类是该研究区域的主要生态类群,这一结果和历史资料报道的结果[5-13]一致。本研究海域范围覆盖从长江内河段至长江冲淡水扩散区外海陆架127°E海域,而从1980s以来,长江口及其邻近海域浮游动物的研究海域的范围主要集中在124°E以西近岸水域,调查海域面积相对较小,且大部分的研究都仅局限于单个或两个季节的浮游动物群落结构研究,未涉及群落季节变化。因此,本文有关长江口及其邻近海域浮游动物群落结构组成种类数明显高于已报道的文献数据(表 5)[5-13]。王金辉[9]等人对2004年2月—11月长江口海域浮游动物群落季节变化进行研究,显示种类数呈现秋季(91种)>夏季(77种)>冬季(73种)>春季(24种)。本研究的结果与其报道的结果存在差异,呈现夏季(317种)>秋季(309种)>春季(230种>冬季(138种)。图 2表明近岸区域浮游动物群落的物种多样性要明显低于远海区域,同时长江冲淡水的影响范围往往延伸至123°E以东的区域,而上述研究的调查水域仅局限于123°E以西近岸水域,因此其浮游动物调查的结果尚不能全面反映长江冲淡水扩散海域浮游动物群落结构特征。
| 经纬度 | 调查时间 | 春季 | 夏季 | 秋季 | 冬季 | 参考文献 |
| Longitude and latitude | Time | Spring | Summer | Autumn | Winter | References |
| 31°00'N—31°32'N 121°21'25″E—122°30'E |
1999—2000年 | - | 87 | - | - | [5] |
| 30°40'00″N—31°36'36″N 121°23'E—123°40'E |
2002年 | - | 74 | - | - | [6] |
| - | 2000—2003年 | 82 | 125 | - | - | [7] |
| - | 2003 | - | 258 | - | - | [8] |
| 30°30'N—31°45'N, 123°00'E以西 |
2004 | 24 | 77 | 91 | 73 | [9] |
| 29°30'N—32°00'N 123°E以西 |
2005 | - | 104 | 95 | - | [10] |
| - | 2006 | - | 295 | - | - | [11] |
| 29.90°N—31.61°N 121.04°E—122.77°E |
2009 | 72 | - | - | - | [12] |
| 30.5°N—32.0°N 122.0°E—123.5°E |
2011 | 97 | - | - | - | [13] |
| 29°30'N—32°30'N 120°00'E—127°30'E |
2006—2007年 | 230 | 317 | 309 | 138 | 本研究 |
本研究优势种的组成中桡足类为最优势类群,箭虫也是重要类群之一。浮游动物生态类群以广温广盐和近岸低盐种类为主,夏、秋季常出现平滑真刺水蚤和肥胖软箭虫等高温高盐暖水种和热带外海种,这与长江口及邻近海域东南部和南部常年受黑潮水和台湾暖流等高温高盐水不同程度的影响相符。桡足类的中华哲水蚤在春季和冬季为第一优势种,优势度非常高,分别为0.253和0.399,这和以往冬季和春季的调查结果[7, 9]一致。在夏、秋季,中华哲水蚤的优势度有所下降,第一优势种的位置分别被背针胸刺水蚤和亚强次真哲水蚤代替,这与刘光兴[8]等和朱延忠[12]等人报道的夏、秋季第一优势种是中华哲水蚤有所差异,但中华哲水蚤仍为本研究夏、秋季浮游动物优势种之一,这主要因本研究调查海域尺度远大于上述研究区域,而长江口及冲淡水海域区域性环境特征差异对统计分析结果产生影响。长江三峡工程已于1997年11月实现了大江截流,2009年工程竣工。随着三峡工程建设,对长江口生态与环境的研究已有报道[8, 11, 24],刘光兴[8]等和陈洪举[11]等人比较了长江三峡工程一期蓄水前后的浮游动物群落结构,发现浮游动物种类数量没有下降反而增多,浮游动物群落结构也没有出现显著变化的结论。本研究比较了三峡工程二期蓄水前历史资料中所记录的浮游动物种类数,表明研究海域的浮游动物物种多样性仍比较高,其种类数并未下降,而是出现了增多的趋势,并且,浮游动物的群落结构也没有出现显著的变化。浮游动物群落结构和多样性的变化反映海洋生态系统的波动,因此,长江口及其邻近海域生态系统变化依然需要长期跟踪调查。
3.2 环境因子对浮游动物群落的影响长江口及其邻近海域在长江冲淡水、台湾暖流和江浙近岸流等多个水系的共同作用影响下,形成独特海洋理化环境,浮游动物的群落分布与水温、盐度、营养盐等都密切相关[25]。环境因子与浮游动物丰度的相关分析(表 4)表明,盐度是影响浮游动物群落丰度分布的主要环境因子,本结果与徐兆礼[7]、刘光兴[8]等、陈洪举[11]等对长江口及其附近海区的研究结果一致。本研究的调查范围主要包括长江河口段至长江冲淡水扩散的海域,受长江径流和外海水等共同影响,长江口及邻近海域水环境的空间差异明显,从长江口到外海存在明显的盐度梯度,盐度的变化影响了浮游动物群落结构,其影响主要有两个方面:1) 在长江冲淡水主干影响的区域内,盐度相对较低,该海域的浮游动物群落组成以河口种和低盐种为主;2) 在陆架外海高盐区和123°E以东受黑潮及其分支台湾暖流影响的区域,外海水团携带了大量的高温高盐种(如:肥胖软箭虫等),对群落组成起主导作用。春季、夏季和秋季,研究海域的黑潮势力强,外海的高温高盐种逐渐地向近岸扩展,而在冬季,受到北方冷空气和寒流的频繁影响,大量高温高盐种消失,浮游动物仅在长江口附近形成小范围的低盐种密集区。在其它河口水域的研究报道中,Li[26]等人对珠江口海域及Mouny[27]等人对英吉利海峡塞纳河口的研究也表明,盐度是影响河口水域浮游动物群落的重要因子。
长江口属于特大型淤泥底质三角洲地带,长江径流每年携带大量泥沙进入长江口水域,在水动力作用下发生沉降、运输和再悬浮等过程。在长江咸淡水交汇区,由于径流和潮流相互作用,出现长江口浑浊带,悬沙浓度较高[28]。本研究表明,在悬浮物平均浓度最低且叶绿素a的平均浓度最高的夏季,浮游动物丰度与悬浮物浓度和叶绿素a浓度呈正相关。而在悬浮物平均浓度较高、叶绿素a的平均浓度较低的秋季,浮游动物丰度均与悬浮物浓度呈现负相关,而与叶绿素a浓度呈正相关。长江冲淡水携带了大量营养盐,有助于浮游植物的生长代谢,但是当海水中悬沙浓度过高时,水体透明度降低,浮游植物的有效辐射光强减弱,浮游植物现存量减少,这将抑制浮游动物生长繁殖的营养需求,本研究与何文珊[29]等和杜萍[30]等人的研究结果一致。Deniz zkundakci[31]等人的研究表明悬沙还会影响滤食性浮游动物的摄食。
温度也是影响浮游动物丰度的重要因子,尤其在春季和夏季(表 4)。春季,研究海域水温升高有助于浮游动物的生长和繁殖。丁峰元[32]等人对我国东海的调查和朱延忠[12]等人对长江口的调查结果均表明温度是影响浮游动物春季分布的主要因子。夏季,海水表层温度普遍升高,高温对浮游动物的生长代谢不利,而底层水温相对较低,部分浮游动物存在度夏机制,比如中华哲水蚤夏季有两个月的时间生活在底层冷水团中,且生物学特征具有明显的休眠特征[33],这可能是中华哲水蚤未能成为本研究夏季第一优势种的主要原因之一。
致谢: 国家海洋局第二海洋研究所陈建芳研究员提供长江口海域水悬浮物数据,黄大吉研究员提供温、盐数据,郝强副研究员提供叶绿素a数据,“海监49号”科学考察船全体船员在海上采样过程中给予帮助,香港科技大学刘红斌教授对写作给予帮助,特此致谢。| [1] | Mousseau L, Lefevre D, Narcy F, Nival P, Andersen V. A one-month study of the zooplankton community at a fixed station in the Ligurian Sea: the potential impact of the species composition on the mineralization of organic matter. Biogeosciences Discussions , 2009, 6 (1) : 995–1019. DOI:10.5194/bgd-6-995-2009 |
| [2] | Madin L P, Horgan E F, Steinberg D K. Zooplankton at the Bermuda Atlantic Time-series Study (BATS) station: diel, seasonal and interannual variation in biomass, 1994-1998. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography , 2001, 48 (8/9) : 2063–2082. |
| [3] | Morgan C A, de Robertis A, Zabel R W. Columbia River plume fronts. I. Hydrography, zooplankton distribution, and community composition. Marine Ecology Progress Series, 2005, 299: 19-31. |
| [4] | 沈焕庭, 潘定安. 长江河口最大浑浊带. 北京: 海洋出版社, 2001 . |
| [5] | 郭沛涌, 沈焕庭, 刘阿成, 王金辉, 杨元利. 长江河口浮游动物的种类组成、群落结构及多样性. 生态学报 , 2003, 23 (5) : 892–900. |
| [6] | 纪焕红, 叶属峰. 长江口浮游动物生态分布特征及其与环境的关系. 海洋科学 , 2006, 30 (6) : 23–30. |
| [7] | 徐兆礼. 长江口邻近水域浮游动物群落特征及变动趋势. 生态学杂志 , 2005, 24 (7) : 780–784. |
| [8] | 刘光兴, 陈洪举, 朱延忠, 齐衍萍. 三峡工程一期蓄水后长江口及其邻近水域浮游动物的群落结构. 中国海洋大学学报 , 2007, 37 (5) : 789–794. |
| [9] | 王金辉, 徐韧, 秦玉涛, 孙亚伟, 刘材材. 长江口基础生物资源现状及年际变化趋势分析. 中国海洋大学学报 , 2006, 36 (5) : 821–828. |
| [10] | 章飞燕, 唐静亮, 李道季, 方涛, 王彪. 夏、秋季长江口及毗邻海域浮游动物的分布与变化. 水生生物学报 , 2009, 33 (6) : 1219–1225. |
| [11] | 陈洪举, 刘光兴. 2006年夏季长江口及其邻近水域浮游动物的群落结构. 北京师范大学学: 自然科学版 , 2009, 45 (4) : 393–398. |
| [12] | 朱延忠, 刘录三, 郑丙辉, 王瑜. 春季长江口及毗邻海域浮游动物空间分布及与环境因子的关系. 海洋科学 , 2011, 35 (1) : 59–65. |
| [13] | 汤新武, 蔡德所, 陈求稳, 王丽, 韩瑞. 三峡工程三期蓄水初期长江口水域春季浮游动物群落特征及其与环境的关系. 环境科学学报 , 2015, 35 (4) : 1082–1088. |
| [14] | 国家海洋局"908"专项办公室. 海洋生物生态调查技术规程. 北京: 海洋出版社, 2006 . |
| [15] | 国家海洋局"908"专项办公室. 海洋化学调查技术规程. 北京: 海洋出版社, 2006 . |
| [16] | 国家海洋局"908"专项办公室. 海洋水文气象调查技术规程. 北京: 海洋出版社, 2006 . |
| [17] | 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志 , 1989, 8 (4) : 13–15, 19-19. |
| [18] | Park G S, Marshall H G. Estuarine relationships between zooplankton community structure and trophic gradients. Journal of Plankton Research , 2000, 22 (1) : 121–136. DOI:10.1093/plankt/22.1.121 |
| [19] | 郑重, 李少菁, 许振祖. 海洋浮游生物学. 北京: 海洋出版社, 1984 . |
| [20] | 唐启升. 中国专属经济区海洋生物资源与栖息环境. 北京: 科学出版社, 2006 . |
| [21] | 郑执中. 黄海和东海西部浮游动物羣落的结构及其季节变化. 海洋与湖沼 , 1965, 7 (3) : 199–204. |
| [22] | 陈亚瞿, 朱启琴, 陈清潮. 东海浮游动物量的分布特征. 海洋学报 , 1980, 2 (4) : 115–121. |
| [23] | 何德华, 杨关铭, 沈伟林, 刘红斌. 浙江沿岸上升流区浮游动物生态研究——Ⅱ.浮游动物种类分布与多样度. 海洋学报 , 1987, 9 (5) : 617–626. |
| [24] | 线薇薇, 刘瑞玉, 罗秉征. 三峡水库蓄水前长江口生态与环境. 长江流域资源与环境 , 2004, 13 (2) : 119–123. |
| [25] | 曹勇, 陈吉余, 张二凤, 陈沈良, 曹卫承. 三峡水库初期蓄水对长江口淡水资源的影响. 水科学进展 , 2006, 17 (4) : 554–558. |
| [26] | Li K Z, Yin J Q, Huang L M, Tan Y H. Spatial and temporal variations of mesozooplankton in the Pearl River Estuary, China. Estuarine, Coastal and Shelf Science , 2006, 67 (4) : 543–552. DOI:10.1016/j.ecss.2005.12.008 |
| [27] | Mouny P, Dauvin J C. Environmental control of mesozooplankton community structure in the Seine Estuary (English Channel). Oceanologica Acta , 2002, 25 (1) : 13–22. DOI:10.1016/S0399-1784(01)01177-X |
| [28] | 周华君. 长江口最大浑浊带特性研究. 重庆交通学院学报 , 1994, 13 (2) : 8–15. |
| [29] | 何文珊, 陆健健. 高浓度悬沙对长江河口水域初级生产力的影响. 中国生态农业学报 , 2001, 9 (4) : 24–27. |
| [30] | 杜萍, 徐晓群, 刘晶晶, 曾江宁, 陈全震, 寿鹿, 廖一波, 周青松. 椒江口春、秋季浮游动物分布特征及与主要环境因子的关系. 应用与环境生物学报 , 2011, 17 (4) : 486–494. |
| [31] | ÖzkundakciD, DugganI C, HamiltonD P. Does sediment capping have post-application effects on zooplankton and phytoplankton?. Hydrobiologia , 2011, 661 (1) : 55–64. |
| [32] | 丁峰元, 李圣法, 董婧, 程家骅. 春季东海区近海浮游动物群落结构及其影响因子. 海洋渔业 , 2005, 27 (1) : 26–32. |
| [33] | 唐启升, 苏纪兰, 孙松, 张经, 黄大吉, 金显仕, 仝龄. 中国近海生态系统动力学研究进展. 地球科学进展 , 2005, 20 (12) : 1288–1299. |