 2017, Vol. 37
2017, Vol. 37文章信息
- 王玉竹, 周萍, 王娟, 马蓓, 刘翊涵, 吴金水
- WANG Yuzhu, ZHOU Ping, WANG Juan, MA Bei, LIU Yihan, WU Jinshui.
- 亚热带几种典型稻田与旱作土壤中外源输入秸秆的分解与转化差异
- Decomposition and transformation of input straw in several typical paddy and upland soils in subtropical China
- 生态学报. 2017, 37(19): 6457-6465
- Acta Ecologica Sinica. 2017, 37(19): 6457-6465
- http://dx.doi.org/10.5846/stxb201607211481
-
文章历史
- 收稿日期: 2016-07-21
- 网络出版日期: 2017-05-27





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
土壤有机碳(SOC)积累取决于碳输入和输出的平衡, 不同的农业管理措施(如施肥、秸秆还田)主要通过增加有机物质输入促进SOC的积累。研究表明, 我国稻田土壤固碳效应明显, 固碳潜力较大, 其中以亚热带区稻田土壤的固碳能力最强, 其固碳速率明显高于旱作土壤甚至是林地土壤[1-4]。稻田土壤较高的固碳速率主要归因于长期植稻体系下水稻产量的增加导致的大量有机物质输入[2, 5]。此外, 我们前期对亚热带一种第四纪红色粘土母质稻田土壤的模拟培养试验表明, 与旱作土壤相比, 虽然稻田土壤微生物生物量更大, 微生物周转速率更快, 但是稻田土壤中添加的作物秸秆和土壤原有有机碳的矿化速率均显著低于旱作土壤[6-7]。这表明, 近30年来我国亚热带地区普遍观察到的稻田土壤较高的固碳速率也与合理的农业管理措施下稻田土壤有机碳相对较低的矿化损失有关。但是, 这一现象在不同母质发育的稻田土壤中是否具有普遍性, 还有待进一步验证。
作为土壤有机碳的重要来源之一, 外源输入秸秆进入土壤后发生一系列的物理-化学-微生物转化过程, 其活性组分是微生物分解产物的主要来源, 这些微生物分解产物进一步通过促进土壤团聚体形成以及与土壤矿物的吸附与化学键合而促进土壤有机碳的积累[8-10]。Spaccini等[11]的模拟培养试验认为, 旱作土壤中新鲜玉米秸秆趋于转化为腐殖酸和富啡酸, 土壤中新鲜植物残体通过矿化分解主要储存于腐殖质的亲水性组分中, 并提出原有腐殖质促进新鲜作物秸秆在土壤中积累的观点。Song等[12]也提出, 稻田土壤中新添加的玉米秸秆碳与土壤铁铝结合态有机碳、游离氧化铁以及胡敏素显著正相关, “新碳”可以通过与土壤矿质成分的化学键合而固定并进一步转化为有机碳的稳定性组分。由此我们推测, 既然外源输入秸秆在亚热带稻田土壤中的矿化率低于旱作土壤, 那么其分解转化产物在土壤碳库不同组分中的分配与稳定也可能与旱作土壤存在差异。
因此, 本研究通过室内模拟培养试验, 选取亚热带四种典型母质发育的稻田和毗邻的旱作土壤, 研究45%田间持水量(WHC)条件(排除水分的影响)下稻田和旱作土壤中外源输入秸秆的矿化与转化差异, 以明确不同母质发育稻田土壤中秸秆的矿化是否均一致性的低于邻近的旱作土壤, 并揭示外源输入秸秆的分解转化产物在土壤中的分配特点, 为进一步认识稻田土壤有机碳矿化和转化的过程与机理提供理论基础。
1 材料与方法 1.1 供试土壤选取湖南省水稻种植历史悠久的四种主要母质类型的稻田土壤为研究对象, 并以毗邻的相同母质的旱作土壤作为对照。四种母质类型的土壤分别为位于长沙县金井镇的花岗岩风化物(113°20′E, 28°33′N)、桃源县盘塘镇的第四纪红色粘土(111°32′E, 29°14′N)、桂阳县余田乡的板页岩风化物(112°69′E, 25°94′N)和桃源县双溪口乡的河流沉积物(111°62′E, 29°10′N)。所选稻田土壤长期种植制度为稻稻闲或稻油/烟轮作, 旱作土壤主要种植油菜、棉花、红薯等, 所有作物均为C3作物。每个样区随机5点采集耕层(0—20 cm)土壤样品, 剔除可见动植物残体, 一部分土样过2 mm筛, 调节土壤含水量至45% WHC, 置于25℃、100%空气湿度和黑暗条件下预培养7—10 d, 另取少量土样室内风干, 过0.149 mm筛, 用于测定土壤基本理化性状。供试土壤的基本理化性状见表 1。
| 土壤母质 Soil parent material |
利用方式 Land use |
pH (H2O) |
粘粒 Clay/% |
土壤有机碳 Soil organic carbon SOC/(g/kg) |
全氮 Total N/(g/kg) |
微生物生物量碳 Microbial biomass carbon MBC/(mg/kg) |
溶解性有机碳 Dissolved organic carbon DOC/(mg/kg) |
| 花岗岩风化物 | 稻田 | 5.02 | 10.82 | 20.29 | 1.79 | 439.31 | 53.34 |
| Granite weathering | 旱地 | 4.94 | 11.33 | 18.38 | 2.88 | 65.96 | 77.26 |
| 第四纪红色粘土 | 稻田 | 5.52 | 15.71 | 21.56 | 2.14 | 833.28 | 62.53 |
| Quaternary red clay | 旱地 | 5.03 | 28.66 | 10.70 | 1.29 | 26.79 | 134.77 |
| 板页岩风化物 | 稻田 | 6.92 | 22.18 | 33.08 | 1.39 | 311.91 | 116.74 |
| Shale weathering | 旱地 | 5.40 | 26.95 | 11.04 | 1.25 | 43.85 | 282.15 |
| 河流沉积物 | 稻田 | 5.34 | 10.88 | 18.57 | 0.92 | 228.36 | 365.08 |
| River sediment | 旱地 | 4.14 | 11.47 | 13.69 | 0.97 | 91.56 | 133.77 |
每种土壤分别设置2个处理:不添加秸秆(CK)和添加秸秆(S)。本试验添加的秸秆为粉碎过2 mm筛的玉米(C4作物)秸秆, 添加量为10 mg/g土(干重)。玉米秸秆的有机碳和全氮含量分别为497.34 g/kg和4.57 g/kg, 13C丰度为-12.74 ‰。
取经预培养过的土样每份20 g(干重)于120 mL培养瓶中, 按上述处理后, 随即用硅胶塞塞住瓶口, 并用704胶密封和塞子之间的空隙。硅胶塞的中间有一小孔, 内插玻璃管, 管外再套一段硅胶软管, 以适合的硅胶塞塞紧软管通口, 作为气体采样口[13]。随后将培养瓶置于25 ℃和黑暗条件下恒温培养180 d。每个处理重复3次。分别于培养的第0、0.5、1、2、5、10、20、40、60、80、100、140、180天采集气体, 测定气体的CO2浓度和δ13C值。
同时取经预培养过的土样每份400 g(干重)于4 L塑料杯中, 按上述处理好后, 置于50 L的塑料桶中, 桶底加少量去离子水以维持桶内100%的空气湿度, 密封塑料桶, 并不定期向桶底添加去离子水, 于25℃和黑暗条件下恒温培养180 d, 每个处理重复3次。培养结束后(第180天)取样测定土壤微生物生物量碳(MBC)、溶解性有机碳(DOC)、颗粒有机碳(POC)、铁铝结合态有机碳(Fe/Al-OC)、腐殖质碳(HMC)、水稳性团聚体各粒径(2000—250 μm、250—53 μm、< 53 μm)SOC及其各自的δ13C值。
1.3 分析测定与计算方法土壤MBC和DOC的测定采用熏蒸提取-碳自动分析仪法[14];土壤POC的测定参考周萍等[15]的方法;土壤Fe/Al-OC的测定采用徐建民等[16]的方法;土壤腐殖质组成采用窦森等[17]的方法;土壤水稳性团聚体采用Cambardella和Elliott[18]的方法。土壤pH采用水土比1:2.5, 用pH计(LRH-250-S)测定;SOC采用重铬酸钾氧化-外加热法测定[19];粘粒含量采用激光粒度分析仪(Mastersizer 2000) 测定;全氮测定采用半微量凯氏定氮法[20]。
气体样品的CO2浓度用气相色谱仪(Agilent Technologies 7890A, USA)测定。CO2及土壤有机碳各组分13C丰度值(δ13C)用稳定性同位素质谱仪(Thermo Scientific MAT 253, 德国)测定, 其中POC和团聚体各粒径测定δ13C前须将土样烘干磨细过0.149 mm筛, MBC、DOC、Fe/Al-OC、HMC测定δ13C前须将浸提液冷冻干燥。δ13C (‰)值的计算方法如下:
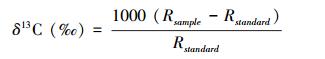
|
式中, R=13C/12C, Rsample与 Rstandard分别表示土壤样品和标准样品的相对丰度。
不同来源有机碳的计算:本研究供试土壤均种植C3作物, 添加玉米(C4)秸秆后进入土壤的新鲜有机物质相对富集13C, 可根据测定的δ13C丰度(‰), 通过下列公式计算出源于外源秸秆和源于本体土壤的有机质组分各占的比例[21]:
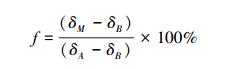
|
式中, f表示土壤样品中源于玉米碳的比例(%), δM为添加玉米秸秆后土壤样品的δ13C值, δA为玉米秸秆的δ13C值, δB为未添加玉米秸秆土壤样品的δ13C值。
1.4 统计分析试验结果均采用3次重复的平均值表示, 用Microsoft Excel 2007软件进行数据处理, 用SPSS 13.0统计软件进行ANOVA分析, LSD法进行差异显著性检验, 显著水平设为0.05, 极显著水平设为0.01。
2 结果与分析 2.1 外源输入秸秆在稻田和旱作土壤中的矿化动态在180 d的培养期内, 外源输入秸秆在土壤中的矿化可分为3个阶段(图 1):0—5 d为快速矿化阶段, 秸秆矿化率较高, 稻田和旱作土壤中分别为11%—14%和14%—19%;6—20 d为慢速矿化阶段, 秸秆的矿化率迅速降低, 分别为3%—7%和2%—7%;21—180 d为稳定矿化阶段, 秸秆矿化率变化较为平缓, 分别仅为1%—2%和2%—3%。培养结束后, 外源输入秸秆在花岗岩风化物、第四纪红色粘土、板页岩风化物和河流沉积物母质稻田土壤中的累积矿化率分别为18%、21%、19%和18%, 而在对应旱作土壤中的矿化率分别为28%、23%、21%和21%, 均以稻田土壤中秸秆的矿化率显著高于对应的旱作土壤(P < 0.05), 尤以花岗岩风化物母质发育的稻田与旱作土壤的差异最大(P < 0.05)。

|
| 图 1 外源输入秸秆在稻田和旱作土壤中的矿化率 Fig. 1 Mineralization ratio of input straw in paddy soils and upland soils 不同小写字母表示同一母质稻田与旱地的差异, P < 0.05 |
土壤中添加外源输入秸秆后, 土壤原有有机碳矿化量在培养的0—20 d内迅速增加, 至第20天时, 4种稻田土壤原有有机碳累积矿化量为120.49—260.89 μg/g, 高于CK 8%—39%(图 2), 差异达显著水平(P < 0.05)。同期内, 旱作土壤原有有机碳累积矿化量为69.22—226.30 μg/g, 高于CK 20%—78%(图 3), 差异达极显著水平(P < 0.01)。而在培养后期(20—180 d), 外源秸秆添加下稻田和旱作土壤原有有机碳的累积矿化量与CK之间无显著差异(P>0.05)。

|
| 图 2 添加外源秸秆对稻田土壤原有有机碳矿化的影响 Fig. 2 Influences of straw amendment on the mineralization of native organic C in paddy soils |

|
| 图 3 添加外源秸秆对旱作土壤原有有机碳矿化的影响 Fig. 3 Influences of straw amendment on the mineralization of native organic C in upland soils |
在180 d培养期内, 花岗岩风化物、第四纪红色粘土、板页岩风化物、河流沉积物母质稻田土壤原有有机碳的累积矿化量分别为376.17、303.41、167.91、225.38 μg/g, 相比CK分别增加了5%、9%、30%、28%(图 2);同期内旱作土壤原有有机碳累积矿化量分别为361.68、286.05、140.02、102.34 μg/g, 高于CK 37%、17%、53%、65%(图 3)。添加外源秸秆后, 所选几种稻田土壤原有有机碳累积矿化量的增长幅度均明显低于相同母质的旱作土壤(P < 0.01), 表现出较弱的激发效应, 特别是花岗岩风化物和第四纪红色粘土母质发育的稻田土壤的激发效应极小。
2.3 外源输入秸秆分解产物在有机碳不同组分中的分配这里仅就花岗岩风化物和第四纪红色粘土母质发育的稻田和旱作土壤为例, 分析培养结束后外源秸秆分解产物在土壤有机碳各组分中的转化与分配。从表 2可以看出, 外源秸秆分解产物在土壤有机碳组分中的转化以向Fe/Al-OC和POC分配为主, 分配比例分别为13%—24%和9%—21%, 其次为HMC(12%—15%), 而向活性组分MBC和DOC中的分配极少, 分配比例分别仅为2%—7%和0.1%-0.7%。
| 土壤母质 Soil parent material |
利用方式 Land use |
微生物 生物量碳 Microbial biomass carbon MBC |
溶解性 有机碳 Dissolved organic carbon DOC |
颗粒有机碳 Particulate organic carbon POC |
铁铝结合态 有机碳 Fe/Al-bound organic carbon Fe/Al-OC |
腐殖质碳 Humus carbon HMC |
| 花岗岩风化物 | 稻田 | 5.5±0.3Bd | 0.1±0.0De | 20.6±0.8Ab | 23.6±1.9Aa | 15.3±1.7Ac |
| Granite weathering | 旱地 | 2.1±0.1Dc | 0.2±0.0Cd | 17.4±0.7Ba | 17.9±1.4Ba | 12.6±1.7Bb |
| 第四纪红色粘土 | 稻田 | 6.5±0.1Ac | 0.3±0.0Bd | 14.6±0.5Cb | 17.3±1.1Ba | 14.3±1.6Ab |
| Quaternary red clay | 旱地 | 3.5±0.1Cc | 0.7±0.1Ad | 8.7±0.9Db | 13.37±1.2Ca | 11.5±1.1Ba |
| 大写字母表示不同土壤同一有机碳组分中的差异, 小写字母表示同一土壤不同有机碳组分中的差异(P < 0.05) | ||||||
在花岗岩风化物母质发育的稻田和旱作土壤中, 外源秸秆分解产物在Fe/Al-OC中的分配比例分别为24%和18%, 在POC中分别为21%和17%, 在HMC中分别为15%和13%, 在MBC中分别为6%和2%, 在DOC中分别为0.1%和0.2%。除了DOC以外, 外源秸秆分解转化产物在有机碳组分中的分配比例均以稻田土壤高于旱作土壤。同样, 在第四纪红色粘土母质发育的稻田和旱作土壤中, 外源秸秆分解产物在Fe/Al-OC中的分配比例分别为17%和13%, 在POC中分别为15%和9%, 在HMC中分别为14%和11%, 在MBC中分别为7%和4%, 在DOC中分别为0.3%和0.7%, 也是以稻田土壤有机碳各组分中的分配比例高于旱作土壤(DOC除外)。
2.4 外源输入秸秆分解产物在水稳性团聚体中的分配外源输入秸秆分解产物在水稳性团聚体中的分配如图 4所示。在花岗岩风化物母质发育的稻田和旱作土壤中, 外源输入秸秆分解产物在2000—250 μm、250—53 μm、< 53 μm粒径中的分配比例分别为10%和7%、7%和5%、6%和3%;在第四纪红色粘土母质发育的稻田和旱作土壤相应粒径中的分配比例分别为13%和6%、8%和7%、5%和7%。两种母质中稻田和旱作土壤之间外源秸秆分解产物的分配差异均表现在2000—250 μm粒径, 以稻田土壤相对较高, 而其余两个粒径的相同粒径中稻田和旱作土壤之间秸秆分解产物的分配比例并无明显差异(P>0.05)。

|
| 图 4 培养180 d后外源秸秆分解产物在土壤水稳性团聚体中的分配 Fig. 4 Distribution of decomposition products of input straw in water-stable aggregates of paddy and upland soils after the incubation for 180 d 不同小写字母表示不同土壤同一粒径之间差异显著, P < 0.05 |
土地利用方式对土壤有机碳库的动态变化起关键作用, 主要通过不同的种植制度、田间管理方式改变土壤结构与养分状况, 改变土壤生物、物理及化学特性, 导致土壤有机碳矿化产生差异[22-23]。通常稻田土壤的微生物数量群落以及功能多样性均高于旱作土壤, 促进了微生物对作物秸秆的利用, 从而导致稻田土壤较旱作土壤更利于外源输入秸秆的矿化[24-25]。已有许多研究表明, 稻田土壤外源输入秸秆的矿化高于旱作土壤, 比如黄东迈等[26]运用14C示踪技术研究发现, 水稻和玉米秸秆在淹水稻田土壤中的矿化速率快于旱地土壤。Inubushi等[27]发现稻田土壤CO2释放量高于旱作土壤约55%。赵次娴等[28]对添加生物质炭后稻田和旱作土壤有机碳的矿化特征研究中表明, 生物质炭施用于旱作土壤比稻田具有更好的固碳减排效果。简燕等[29]也在研究中指出稻田土壤自养微生物同化碳的矿化速率明显大于旱作土壤。但我们前期[6, 30]对亚热带一种稻田土壤外源输入秸秆矿化的研究报道与上述文献报道的结果并不一致, 尽管稻田土壤微生物生物量远大于旱作土壤, 但稻田土壤中外源输入秸秆的矿化率及其对土壤原有有机碳矿化的激发效应均显著低于旱作土壤, 从而表现出较高的有机碳积累效应。在此基础上, 本研究进一步发现, 外源输入秸秆在不同母质发育稻田土壤中的累积矿化率均低于对应的旱作土壤, 具有一定的普遍性, 表明稻田土壤有机质周转可能存在特殊的生物地球化学过程, 从而更有利于外源输入秸秆在稻田土壤中的积累。
好氧条件会促进土壤有机碳的矿化[31], 且有机碳矿化随着土壤粘粒含量的增加而减少, 粘粒含量较高的土壤对作物秸秆分解起保护作用[32-33]。本研究中稻田和旱作土壤均在45%WHC条件下培养, 且稻田土壤粘粒含量均不同程度地小于旱作土壤, 稻田土壤的环境条件似乎更有利于有机碳的矿化。但是, 本研究稻田土壤中外源输入秸秆及其对原有有机碳矿化的激发效应并非如上述所说会高于旱作土壤, 而是受到了明显的抑制, 这表明水分条件和粘粒含量可能并非影响稻田土壤有机碳矿化的主要因素。秸秆输入通过影响土壤微生物群落组成及活性从而对土壤原有有机碳矿化产生正的或负的激发效应[34-35]。关于稻田土壤有机碳矿化的一些研究也认为, 外源秸秆添加使土壤原有有机碳矿化表现出显著的负激发效应[7, 36]或无显著激发效应[6, 30]。本研究结果也显示, 尽管稻田土壤碳库与微生物生物量均大于旱作土壤, 但外源秸秆添加对稻田土壤原有有机碳矿化的激发效应显著低于对应的旱作土壤, 且花岗岩风化物和第四纪红色粘土母质发育的稻田土壤几乎无激发效应。而旱作土壤添加秸秆后可能由于团聚体的物理保护作用减弱, 导致土壤有机碳矿化的激发效应随秸秆添加量的增加而增强, 从而使土壤稳定性有机碳的比例减少[37]。
土壤中外源秸秆分解快慢是土壤微生物活动强弱的外在表现, 以往研究认为微生物数量和活性决定了土壤有机碳的分解[38]。但是, Kemmitt等[39]提出的“控制阀”假设认为土壤有机质的分解包括由非生物活性有机质通过非生物过程转化为生物活性有机质, 再由生物过程进行矿化两个阶段。前者为土壤有机质分解的控制性阶段, 主要机制包括:化学氧化和水解、从团聚体和生物不可及空隙的释放、从固相中的解析以及胞外酶的作用。本研究中稻田土壤微生物生物量碳含量及外源秸秆分解产物在微生物生物量碳中的分配比例均高于对应的旱作土壤, 但稻田土壤中外源输入的秸秆并未表现出较高的矿化率, 这也从侧面验证了稻田土壤中外源输入秸秆的矿化不完全受土壤微生物的影响。近年来也有研究指出, 土壤微生物生物量及其活性、区系组成并不能有效指示有机碳矿化, 与土地利用变化直接关联的土壤结构、土壤团聚体以及与之相关的物理化学性状可能在土壤有机碳的矿化过程中发挥重要的作用, 并进一步影响土壤有机碳的积累[40-41]。Wu[6]等也在研究中发现新鲜作物秸秆矿化并不随土壤粘粒含量的增加而减少, 土壤中存在大量的铁铝氧化物能减少土壤有机碳的矿化, 更好的保护有机质及其分解产物。本文研究结果显示, 稻田土壤中外源输入秸秆向POC、Fe/Al-OC、HMC和2000—250 μm水稳性团聚体分配的比例较高, 分别为15%—21%、17%—24%、14%—15%和10%—13%, 均明显高于对应的旱作土壤。相比于旱作土壤, 外源输入秸秆在稻田土壤中较低的矿化率可能与其分解转化产物受土壤水稳性团聚体的物理保护、与铁铝氧化物的化学键合以及向腐殖质组分的分配作用较强有关。但是, 稻田土壤中外源输入秸秆的分解转化产物在DOC中的分配比例却低于旱作土壤。而DOC作为微生物分解有机碳的直接利用底物, 导致外源秸秆在旱作土壤中通过DOC分解矿化的损失较高。这也进一步从侧面说明, 稻田土壤对有机碳的物理保护和化学结合稳定作用较强, 且团聚体的物理保护作用进一步促进SOC的化学稳定化过程, 从而促使稻田土壤相比旱作土壤拥有较低的有机碳矿化速率和较高的有机碳积累能力[42]。雷敏等[43]也提到亚热带稻田土壤有机碳物理和化学保护作用的增强, 有利于稻田土壤维持较高的固碳速率, 从而促进有机碳的积累。Song等[12]报道指出土壤外源碳最终主要是通过转化为稳定性腐殖质碳组分被固存。
4 结论在180 d培养期内, 45% WHC条件下所选4种典型母质发育的稻田土壤中外源输入秸秆累积矿化率及其对土壤原有有机碳矿化的激发效应均一致性地低于对应的旱作土壤, 可能具有一定普遍性。与旱作土壤相比, 稻田土壤中外源输入秸秆的分解产物受水稳性团聚体的物理保护、与铁铝氧化物的化学键合以及向有机碳稳定组分的分配均较强, 从而可能抑制了外源秸秆的分解矿化, 促进其在稻田土壤中的较高积累。
致谢: 感谢中国科学院亚热带农业生态研究所公共技术服务中心王久荣主任、袁红朝、耿梅梅对本研究工作的帮助。| [1] | Pan G X, Xu X W, Smith P, Pan W N, Lal R. An increase in topsoil SOC stock of China's croplands between 1985 and 2006 revealed by soil monitoring. Agriculture, Ecosystem & Environment, 2010, 136(1/2): 133–138. |
| [2] | Wu J S. Carbon accumulation in paddy ecosystems in subtropical China: Evidence from landscape studies. European Journal of Soil Science, 2011, 62(1): 29–34. DOI:10.1111/ejs.2011.62.issue-1 |
| [3] | Kalbitz K, Kaiser K, Fiedler S, Kölbl A, Amelung W, Bräuer T, Cao Z H, Don A, Grootes P, Jahn R, Schwark L, Vogelsang V, Wissing L, Kögel-Knabner I. The carbon count of 2000 years of rice cultivation. Global Change Biology, 2012, 19(4): 1107–1113. |
| [4] | Huang S, Sun Y N, Zhang W J. Changes in soil organic carbon stocks as affected by cropping systems and cropping duration in China's paddy fields: a meta-analysis. Climatic Change, 2012, 112(3/4): 847–858. |
| [5] | Yan X Y, Cai Z C, Wang S W, Smith P. Direct measurement of soil organic carbon content change in the croplands of China. Global Change Biology, 2011, 17(3): 1487–1496. DOI:10.1111/gcb.2011.17.issue-3 |
| [6] | Wu J S, Zhou P, Li L, Su Y R, Yuan H Z, Syers J K. Restricted mineralization of fresh organic materials incorporated into a subtropical paddy soil. Journal of the Science of Food and Agriculture, 2012, 92(5): 1031–1037. DOI:10.1002/jsfa.v92.5 |
| [7] | 王嫒华, 苏以荣, 李杨, 吴金水, 郑华, 朱捍华, 胡乐宁. 稻草还田条件下水田和旱地土壤有机碳矿化特征与差异. 土壤学报, 2011, 48(5): 979–987. DOI:10.11766/trxb201011030456 |
| [8] | 周萍, 宋国菡, 潘根兴, 李恋卿, 张旭辉, WuL S. 南方三种典型水稻土长期试验下有机碳积累机制研究I.团聚体物理保护作用. 土壤学报, 2008, 45(6): 1063–1071. |
| [9] | Zhou P, Song G H, Pan G X, Li L Q, Zhang X H. Role of chemical protection by binding to oxyhydrates in SOC sequestration in three typical paddy soils under long-term agro-ecosystem experiments from South China. Geoderma, 2009, 153(1/2): 52–60. |
| [10] | Cotrufo M F, Wallenstein M D, Boot C M, Denef K, Paul E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter?. Global Change Biology, 2013, 19(4): 988–995. DOI:10.1111/gcb.12113 |
| [11] | Spaccini R, Piccolo A, Haberhauer G, Gerzabek M H. Transformation of organic matter from maize residues into labile and humic fractions of three European soils as revealed by 13C distribution and CPMAS-NMR spectra. European Journal of Soil Science, 2000, 51(4): 583–594. DOI:10.1111/ejs.2000.51.issue-4 |
| [12] | Song X Y, Li L Q, Zheng J F, Pan G X, Zhang X H, Zheng J W, Hussain Q, Han X J, Yu X Y. Sequestration of maize crop straw C in different soils: role of oxyhydrates in chemical binding and stabilization as recalcitrance. Chemosphere, 2012, 87(6): 649–654. DOI:10.1016/j.chemosphere.2012.01.038 |
| [13] | 周萍, 宋国菡, 潘根兴, 李恋卿, 张旭辉, WuL S. 三种南方典型水稻土长期试验下有机碳积累机制研究Ⅱ.团聚体内有机碳的化学结合机制. 土壤学报, 2009, 46(2): 263–273. |
| [14] | 吴金水, 林启美, 黄巧云, 肖和艾. 土壤微生物生物量测定方法及其应用. 北京: 气象出版社, 2006: 57–60. |
| [15] | 周萍, 张旭辉, 潘根兴. 长期不同施肥对太湖地区黄泥土总有机碳及颗粒态有机碳含量及深度分布的影响. 植物营养与肥料学报, 2006, 12(6): 765–771. DOI:10.11674/zwyf.2006.0602 |
| [16] | 徐建明, 赛夫, 袁可能. 土壤有机矿质复合体研究IX:钙键复合体和铁铝键复合体中腐殖质的性状特征. 土壤学报, 1999, 36(2): 168–178. DOI:10.11766/trxb199708050204 |
| [17] | 窦森, 于水强, 张晋京. 不同CO2浓度对玉米秸秆分解期间土壤腐殖质形成的影响. 土壤学报, 2007, 44(3): 458–466. DOI:10.11766/trxb200601280312 |
| [18] | Cambardella C A, Elliott E T. Methods for physical separation and characterization of soil organic matter fractions. Geoderma, 1993, 56(1/4): 449–457. |
| [19] | 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999. |
| [20] | 刘光崧. 土壤理化分析与剖面描述. 北京: 中国标准出版社, 1996. |
| [21] | Phillips D L, Gregg J W. Uncertainty in source partitioning using stable isotopes. Oecologia, 2001, 127(2): 171–179. DOI:10.1007/s004420000578 |
| [22] | Kasel S, Bennett L T. Land-use history, forest conversion, and soil organic carbon in pine plantations and native forests of south eastern Australia. Geoderma, 2007, 137(3/4): 401–413. |
| [23] | 张文娟, 廖洪凯, 龙健, 李娟, 刘灵飞. 贵州喀斯特山区土地利用对土壤有机碳及其周转速率的影响. 生态学杂志, 2014, 33(5): 1297–1303. |
| [24] | 李忠佩, 吴晓晨, 陈碧云. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化. 中国农业科学, 2007, 40(8): 1712–1721. |
| [25] | Yuan H Z, Ge T D, Wu X H, Liu S L, Tong C L, Qin H L, Wu M N, Wei W X, Wu J S. Long-term field fertilization alters the diversity of autotrophic bacteria based on the ribulose-1, 5-biphosphate carboxylase/oxygenase (RubisCO) large-subunit genes in paddy soil. Applied Microbiology and Biotechnology, 2012, 95(4): 1061–1071. DOI:10.1007/s00253-011-3760-y |
| [26] | 黄东迈, 朱培立, 王志明, 余晓鹤. 旱地和水田有机碳分解速率的探讨与质疑. 土壤学报, 1998, 35(4): 482–492. DOI:10.11766/trxb199712060407 |
| [27] | Inubushi K, Furukawa Y, Hadi A, Purnomo E, Tsuruta H. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use change in tropical peatlands located in coastal area of south Kalimantan. Chemosphere, 2003, 50(3): 603–608. |
| [28] | 赵次娴, 陈香碧, 黎蕾, 肖和友, 刘坤平, 何寻阳, 苏以荣. 添加蔗渣生物质炭对农田土壤有机碳矿化的影响. 中国农业科学, 2013, 46(5): 987–994. |
| [29] | 简燕, 葛体达, 吴小红, 周萍, 王久荣, 袁红朝, 邹冬生, 吴金水. 稻田与旱地土壤自养微生物同化碳在土壤中的矿化与转化特征. 应用生态学报, 2014, 25(6): 1708–1714. |
| [30] | 李玲, 肖和艾, 吴金水. 红壤旱地和稻田土壤中有机底物的分解与转化研究. 土壤学报, 2007, 44(4): 669–674. DOI:10.11766/trxb200604290413 |
| [31] | Hao R J, Li Z P, Che Y P. Differences in organic c mineralization between aerobic and submerged conditions in paddy soils of southern Jiangsu Province, China. Agricultural Sciences in China, 2011, 10(9): 1410–1418. DOI:10.1016/S1671-2927(11)60134-X |
| [32] | Xu X, Shi Z, Li D J, Rey A, Ruan H H, Craine J M, Liang J Y, Zhou J Z, Luo Y Q. Soil properties control decomposition of soil organic carbon: Results from data-assimilation analysis. Geoderma, 2016, 262: 235–242. DOI:10.1016/j.geoderma.2015.08.038 |
| [33] | 李忠佩, 刘明, 江春玉. 红壤典型区土壤中有机质的分解、积累与分布特征研究进展. 土壤, 2015, 47(2): 220–228. |
| [34] | Paterson E, Sim A. Soil specific response functions of organic matter mineralization to the availability of labile carbon. Global Change Biology, 2013, 19(5): 1562–1571. DOI:10.1111/gcb.12140 |
| [35] | Kuzyakov Y. Priming effects: interactions between living and dead organic matter. Soil Biology and Biochemistry, 2010, 42(9): 1363–1371. DOI:10.1016/j.soilbio.2010.04.003 |
| [36] | Ge T D, Yuan H Z, Zhu H H, Wu X H, Nie S A, Liu C, Tong C L, Wu J S, Brookes P C. Biological carbon assimilation and dynamics in a flooded rice-soil system. Soil Biology and Biochemistry, 2012, 48: 39–49. DOI:10.1016/j.soilbio.2012.01.009 |
| [37] | Shahbaz M, Kuzyakov Y, Heitkamp F. Decrease of soil organic matter stabilization with increasing inputs: Mechanisms and controls. Geoderma, 2016. DOI:10.1016/j.geoderma.2016.05.019 |
| [38] | Kaschuk G, Alberton O, Hungria M. Three decades of soil microbial biomass studies in Brazilian ecosystems: lessons learned about soil quality and indications for improving sustainability. Soil Biology and Biochemistry, 2010, 42(1): 1–13. DOI:10.1016/j.soilbio.2009.08.020 |
| [39] | Kemmitt S J, Lanyon C V, Waite I S, Wen Q, Addiscott T M, Bird N R A, O'Donnell A G, Brookes P C. Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass-a new perspective. Soil Biology and Biochemistry, 2008, 40(1): 61–73. DOI:10.1016/j.soilbio.2007.06.021 |
| [40] | Strickland M S, Callaham Jr M A, Davies C A, Lauber C L, Ramirez K, Richter Jr D D, Fierer N, Bradford M A. Rates of in situ carbon mineralization in relation to land-use, microbial community and edaphic characteristics. Soil Biology and Biochemistry, 2010, 42(2): 260–269. DOI:10.1016/j.soilbio.2009.10.026 |
| [41] | Ball B C. Soil structure and greenhouse gas emissions: a synthesis of 20 years of experimentation. European Journal of Soil Science, 2013, 64(3): 357–373. DOI:10.1111/ejss.2013.64.issue-3 |
| [42] | Cui J, Li Z X, Liu Z T, Ge B M, Fang C M, Zhou C L, Tang B P. Physical and chemical stabilization of soil organic carbon along a 500-year cultived soil chronosequence originating from estuarine wetlands: Temporal patterns and land use effects. Agriculture, Ecosystems & Environment, 2014, 196: 10–20. |
| [43] | 雷敏, 周萍, 黄道友, 刘守龙, 朱捍华, 童成立. 长期施肥对水稻土有机碳分布及化学结合形态的影响. 生态学杂志, 2012, 31(4): 967–974. |