 2017, Vol. 37
2017, Vol. 37文章信息
- 王晶, 吕昭智, 尹传华, 李锦辉, 吴文岳
- WANG Jing, LÜ Zhaozhi, YIN Chuanhua, LI Jinhui, WU Wenyue.
- 塔克拉玛干沙漠北缘柽柳灌丛沙堆对甲虫的庇护作用
- The shelter belt effect: beetles in the litter-layer of Tamarix nebkha in the north rim of Taklamakan
- 生态学报. 2017, 37(19): 6504-6510
- Acta Ecologica Sinica. 2017, 37(19): 6504-6510
- http://dx.doi.org/10.5846/stxb201607091408
-
文章历史
- 收稿日期: 2016-07-09
- 网络出版日期: 2017-05-27





2. 中国科学院新疆生态与地理研究所, 乌鲁木齐 830011;
3. 农一师3团农业科, 阿克苏 843300
2. Xinjiang Institute of Geography and Ecology, Urumqi 830011, China;
3. Agriculture Section of Agricultural Division One Farm Three, Aksu 843300, China
植物群落的物种组成结构和分布特点, 特别是植物的种类数、生物量、高度与昆虫群落的结构和物种多样性密切相关, 并对昆虫群落的稳定性产生深刻的影响[1-2]。在沙漠或者干旱地区地区由于资源匮乏以及环境胁迫植物与昆虫之间的关系更为密切。在索诺兰沙漠中植物群落为昆虫提供大量的资源(叶片和花), 有利于昆虫多样性的增加, 其中叶片生物量在44%和51%以上水平上可以解说物种数及其种群数量变异程度, 花的生物量在32%以上的水平上说明昆虫物种的变异程度[3]。不仅如此, 昆虫多样性也与寄主植物体内的水分含量也存在一定的关系, 在一些类群植物中水分与昆虫群落存在正相关[4]。Wenninger等的工作也支持这一点:昆虫多样性与植物多样性及其灌溉是正相关的[5]。此外, 相同生境中的昆虫群落结构和物种多样性除了受植物群落外还受到土壤硬度和含水量等环境因素的影响[1-2], 还会因季节的推移而发生明显的变化, 从而造成群落内不同昆虫种群季节消长明显[6-10]。
植物群落在沙漠地区分布受多种因子的影响, 如海拔、土壤生物结皮、土壤环境和地理位置[11], 这些因子也可能间接地影响昆虫的多样性[12]。塔里木盆地地处新疆南部, 属极端干旱区, 自然环境恶劣, 严酷的生态环境造成区域内昆虫的物种数量贫乏, 种群的数量极为有限, 生态系统十分脆弱。灌丛沙堆是干旱、半干旱地区的一种典型的风积生物地貌类型[13], 它可以通过富集凋落物、遮荫、收集茎干径流等方式改善局部的微环境条件, 从而吸引植食性昆虫和肉食性动物前来栖息[14], 进而为极端干旱区生物多样性的保护提供庇护场所[13-16]。柽柳包是灌丛沙堆的一种[17-18], 主要分布于山前冲积扇、河岸低阶地绿洲与流动沙丘之间的区域。尽管柽柳的肥岛、盐岛效应已经有所报道[19-20], 但关于它在干旱区生态系统中的功能, 诸如对昆虫群落多样性的维持和保育等鲜有报道发表。本文通过比较典型荒漠区柽柳包及沙漠裸地间地表甲虫的物种组成、丰富度、个体数量和多样性等差异及季节变化动态, 分析地表甲虫对栖息环境的选择性, 探讨柽柳灌丛沙堆在极端干旱区对地表甲虫群落物种多样性维持和保育机制, 进一步理解在沙漠地区动物与植物的复杂生态关系, 从而为脆弱生态系统生物多样性保护与管理提供理论支持。
1 材料和方法 1.1 研究区概况研究区位于新疆维吾尔自治区阿克苏市南130 km处的喀拉库勒镇(40°22′N, 80°03′E), 地处, 天山南麓塔克拉玛干沙漠北缘, 塔里木河的上游段, 为和田河和叶尔羌河的交汇处, 海拔1050 m。属暖温带典型的极端大陆性干旱荒漠气候, 夏季炎热、冬季寒冷、春季升温快而不稳定, 秋季降温迅速, 昼夜温差大。多年平均气温11℃, 极端最高气温43.9℃, 极端最低气温-27.1℃, 年均降水量62.7 mm, 年均蒸发量2337.5 mm。研究区位于典型荒漠区域边缘, 距离绿洲防护林带500 m左右, 地处偏僻, 受人为扰动少, 以起伏连绵的沙地为主要地貌, 生境异质性较低, 区域内分散着少数的柽柳包, 植物种类稀少, 是研究自然生境地表甲虫群落结构和优势种密度的理想场所。
1.2 调查方法和数据处理柽柳包地表甲虫群落多样性调查采用陷阱法(陷阱为塑料杯, 口径7 cm, 深12.5 cm, 埋入土中, 放置时塑料杯口与地面平齐, 由于研究区域蒸发量大, 未使用诱捕剂), 收集地表甲虫, 选取6个柽柳包(表 1)和3块沙漠裸地(对照)作为试验样点, 每个柽柳包沿周长均匀放置9个陷阱, 2个陷阱间隔3m左右, 每个沙漠裸地样点中放置18个陷阱, 排成3排, 2个陷阱之间间隔2m左右, 2排间隔3m左右。2次试验时间间隔为30d, 甲虫标本用75%的酒精溶液浸泡保存。甲虫标本鉴定主要由研究各类群的专家帮助完成, 数量和种类较少没有找专家鉴定的类群按查找相关文献鉴定到科。
| 编号 Number |
基底周长/m Perimeter |
斜边长度/m Hypotenuse length |
表面积/m2 Superficial area |
坡度/(°) Gradient |
植株高度/m Plant height |
表面覆盖物情况 Cladding thickness |
| 1 | 32 | 5.0 | 80 | 25 | 5.2 | 柽柳落叶少 |
| 2 | 26 | 3.6 | 46.8 | 30 | 4.8 | 柽柳落叶层厚 |
| 3 | 35 | 5.4 | 94.5 | 35 | 5.1 | 柽柳落叶少 |
| 4 | 30 | 5.2 | 78 | 40 | 5.0 | 柽柳落叶层稍厚 |
| 5 | 34 | 5.4 | 91.8 | 45 | 5.4 | 柽柳落叶稍多 |
| 6 | 37 | 5.0 | 92.5 | 40 | 4.7 | 柽柳落叶少 |
优势种密度调查时间为2006年7月16日至7月26日, 选取6个两两间距离不小于30m的柽柳包, 用陷阱法(方法同上)收集标本, 在每个柽柳包基部均匀放置30个陷阱, 2个陷阱间隔1m左右;用油漆在捕获的甲虫鞘翅一侧作标记, 连续11d每天对所获标本进行1次标记重捕, 为验证地表甲虫在不同柽柳包间是否存在迁移现象, 同一柽柳包上捕获的标本采用相同颜色油漆标记, 不同柽柳包所用油漆颜色不同, 记录每天标本的捕获数、已标记数和新标记数。
利用Schumacker-Eschmeyer公式估计各柽柳包上甲虫优势种群的大小。
种群大小的估计量(N) 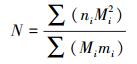
1/N的方差 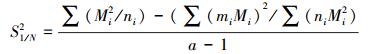
1/N的标准误 
种群大小(N)的95%的置信区间可由1/N的置信区间得来, 1/N的置信区间为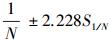
式中, ni是在第i次取样时捕获的甲虫总数, mi在第i次取样时捕获的甲虫中已标记的甲虫总数, Mi是在第i次取样时种群中已标志的甲虫总数。
物种丰富度指群落中的物种数, 以S表示。
群落多样性本文以Shannon-Wiener(1963) 多样性指数(H′)表示:
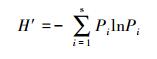
|
式中, 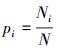
均匀性指数(J)的计算公式: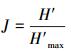
式中H′max为H′的最大理论值, 即假定群落内各个物种均以相同的比例存在时的H′值。
Simpson的优势集中性指数(C)的计算式: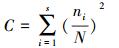
式中, ni为第i个优势种在群落中的重要值, N为群落的总重要值[12]。
2 结果与分析 2.1 柽柳包及沙漠裸地地表甲虫群落结构和数量在2005年和2006年调查过程中发现柽柳包上地表甲虫个体数量和物种数均高于沙漠裸地(表 2)。2005年调查中共捕获地表甲虫23种, 其中柽柳包上21种, 沙漠裸地中10种, 在陷阱设置数量相同的条件下, 在柽柳包上捕获的地表甲虫的个体数量是沙漠裸地的1.7倍;2006年共捕获地表甲虫20种, 在柽柳包上有16种, 沙漠裸地中有12种, 柽柳包上捕获地表甲虫的个体数量是沙漠裸地的2.15倍;通过两年的调查分析比较发现柽柳包上地表甲虫的物种数和个体数量均较高。2005年, 柽柳包地表甲虫的多样性指数和均匀性指数比沙漠裸地中的高, 优势集中性指数较低, 说明柽柳包上地表甲虫群落的稳定性比沙漠裸地中的高。
| 物种及群落结构特征值 Species and Characteristic |
2005年柽柳包 2005 Tamarix nebkha |
2005年沙裸地 2005 bare area |
2006年柽柳包 2006 Tamarix nebkha |
2006年沙裸地 2006 bare area |
| 髋胫小土甲Penthicicus koltzei Reitter, 1896 | 229 | 185 | 250 | 102 |
| 笨土甲一种Penthicus Reitter sp. | 1 | 0 | 0 | 0 |
| 光胸宽漠甲Sternoplax costatissima Reitter, 1899 | 25 | 5 | 12 | 0 |
| 光滑胖漠甲Trigonoscelis sublaeuigata Reitter, 1887 | 13 | 1 | 9 | 8 |
| 格氏驼毛甲Cyphostethe grombczewski Semenow, 1891 | 2 | 0 | 6 | 1 |
| 亚皱土甲Gonocephalum subrugulosum Reitter, 1887 | 3 | 0 | 0 | 0 |
| 喀什琵甲Blaps kashgarensis Bates | 3 | 0 | 1 | 0 |
| 肩脊砚甲Cyphogenia humeralis Bates, 1879 | 2 | 0 | 1 | 1 |
| 东憋甲属Anatolica sp.1879 | 4 | 0 | 0 | 0 |
| 高胸鳖甲Colposcelis montivaga Bates, 1879 | 0 | 1 | 26 | 4 |
| Calatus sp. | 9 | 8 | 2 | 33 |
| 柽柳白筒象Lioclenus clathratus (Olivier) | 4 | 0 | 0 | 0 |
| 粉红锥喙象Conorrhynchus conirostris Gyllenhyl | 10 | 3 | 6 | 4 |
| 黄褐纤毛象Tanymecus urbanus Gyllenhyl | 7 | 0 | 0 | 0 |
| 云斑毛足象Phacephorus nebulosus Fahraeus | 1 | 0 | 0 | 0 |
| 横脊象属Platymcteropsis spp. | 14 | 0 | 0 | 0 |
| 阎甲科一种Histeridae | 5 | 1 | 18 | 7 |
| 肖叶甲亚科一种Eumolpidae | 0 | 1 | 0 | 3 |
| 粪金龟科一种Geotrupidae | 0 | 2 | 3 | 3 |
| 叩甲科一种Elateridae | 1 | 0 | 0 | 0 |
| 伪天牛科一种Oedemeridae | 1 | 0 | 0 | 0 |
| 伪叶甲科一种Lagriidae | 12 | 2 | 0 | 0 |
| 伪叶甲科一种Lagriidae | 13 | 0 | 0 | 0 |
| 皮蠹科一种Dermestidae | 1 | 0 | 0 | 1 |
| 蜉金龟科一种Aphodiidae | 0 | 0 | 1 | 1 |
| 苏氏宽漠甲Sternoplax souvorowiana Reitter, 1907 | 0 | 0 | 2 | 0 |
| 大瘤宽漠甲Sternoplax locerta Bates, 1879 | 0 | 0 | 1 | 0 |
| 卓丹楔毛甲Trichosphaena chotanica Semenov, 1903 | 0 | 0 | 1 | 0 |
| 天牛科一种Cerambycidae | 0 | 0 | 1 | 0 |
| 甲虫个体数Individuals | 360 | 209 | 340 | 158 |
| 丰富度(S)Species richness | 21 | 10 | 16 | 12 |
| 多样性指数(H′)Shanon-Weiner index | 1.5995 | 0.5743 | 1.1399 | 1.2963 |
| 均匀性指数(J)Evenness index | 0.5254 | 0.2494 | 0.4111 | 0.5217 |
| 优势集中性指数(C)Simpson | 0.4172 | 0.7860 | 0.5521 | 0.3933 |
按54个陷阱中地表甲虫的数量为标准处理2年的甲虫数据(图 1)。2005年柽柳包和沙漠裸地中的地表甲虫的数量在6月份最高, 7、8、9月份依次减少。2006年柽柳包上地表甲虫的数量从4月份开始上升, 7月份地表甲虫的数量最多;沙漠裸地中地表甲虫的季节动态基本上与柽柳包中的一致, 但甲虫的数量在6月份最多。造成甲虫数量峰值出现的月份不同的原因可能与年际间最高气温出现时间不同有关系:2005年气温峰值可能在7月, 而2006年则提前到了6月。而气温峰值的出现也是造成柽柳包与沙漠裸地甲虫数量差距加大的原因。因为环境越极端, 柽柳包的微环境效应就越突出。沙漠地区7、8月份地表温度到达50℃以上, 由于柽柳的遮荫作用和相对丰富的食物资源, 可以吸引更多的地表甲虫在柽柳包栖息。荒漠优势种髋胫小土甲与地表甲虫季节动态相似(图 2), 这表明沙漠地区地表甲虫在适应极端环境中具有相同的趋势。

|
| 图 1 柽柳包与沙漠裸地中地表甲虫的季节动态 Fig. 1 Seasonal dynamics of the litter-layer beetles between the Tamarix nebkha and bare desert |

|
| 图 2 柽柳包与沙漠裸地中地表甲虫优势种(髋胫小土甲)的季节动态 Fig. 2 Seasonal dynamics of the dominant species between the Tamarix nebkha and bare desert |
柽柳包上髋胫小土甲标记过程及其密度见表 3和表 4。对柽柳包上优势种甲虫髋胫小土甲的个体数量的调查发现, 优势种甲虫髋胫小土甲在柽柳包上数量相对较大, 每平方米密度达到13头以上。同时, 2, 4, 5号3个柽柳包上髋胫小土甲密度相对较高, 数量较多, 与调查柽柳包基本特征(表 1)对比发现, 2, 4, 5号3个柽柳包落叶层较厚, 植物有机质等积累较多, 有利于甲虫的生存与食物的获取, 而1, 3, 6号柽柳包落叶少, 甲虫数量相对较少。
| 取样日期 Date |
捕获数ni Catches |
已标记数mi Marker |
新标记数Ui New marker |
已标记总数Mi Total marker |
niMi | Mimi | niMi2 | mi2/ni |
| 07-16 | 40 | 0 | 40 | 0 | 0 | 0 | 0 | 0 |
| 07-17 | 33 | 1 | 32 | 40 | 1320 | 40 | 52800 | 0.0303 |
| 07-18 | 42 | 6 | 36 | 72 | 3024 | 432 | 217728 | 0.8571 |
| 07-19 | 61 | 4 | 57 | 108 | 6588 | 432 | 711504 | 0.2623 |
| 07-20 | 45 | 2 | 43 | 165 | 7425 | 330 | 1225125 | 0.0889 |
| 07-21 | 39 | 5 | 34 | 208 | 8112 | 1040 | 1687296 | 0.6410 |
| 07-22 | 31 | 7 | 24 | 242 | 7502 | 1694 | 1815484 | 1.5806 |
| 07-23 | 49 | 9 | 40 | 266 | 13034 | 2394 | 3467044 | 1.6531 |
| 07-24 | 30 | 8 | 22 | 306 | 9180 | 2448 | 2809080 | 2.1333 |
| 07-25 | 51 | 6 | 45 | 328 | 16728 | 1968 | 5486784 | 0.7059 |
| 07-26 | 24 | 2 | 0 | 373 | 8952 | 746 | 3339096 | 0.1667 |
| 总和Sum | 445 | 50 | 373 | 2108 | 81865 | 11524 | 20811941 | 8.1192 |
| 在调查过程中, 在有些柽柳包上捕获的甲虫出现个别死亡, 所以每天的ni并不一定等于mi和ui之和 | ||||||||
| 序号 Number |
捕获总数ni Total capture |
累计标记数mi Total marker |
种群大小估计量N Population Size |
N的95%的置信区间 Confidence interval of 95% |
平均密度/(头/m2) Mean density |
| 1 | 445 | 50 | 1806 | 1320—2856 | 17—36 |
| 2 | 230 | 9 | 2873 | 1693—9473 | 36—202 |
| 3 | 181 | 7 | 1870 | 1192—4326 | 13—46 |
| 4 | 235 | 6 | 3901 | 2354—11384 | 30—146 |
| 5 | 567 | 32 | 4544 | 3549—6313 | 39—69 |
| 6 | 376 | 27 | 2126 | 1625—3074 | 18—33 |
在调查中发现7月24号在3号柽柳包上捕到1头用绿色油漆(4号柽柳包上标记所用)标记的标本, 3号和4号柽柳包距离32m, 表明甲虫在相似的生境之间存在一定的迁移现象。
3 讨论与结论 3.1 柽柳包在干旱区生态系统中的功能在于为地表甲虫群落物种多样性的保育提供了庇护所在本研究中, 柽柳包上地表甲虫群落的物种多样性和优势种种群数量要明显高于沙漠裸地(图 1)。这表明在干旱区生态系统中, 柽柳包对地表甲虫起着庇护所的作用。柽柳包之所以有这样的庇护功能在于灌丛沙堆的(柽柳包)的形成有利于降低风速, 遮蔽阳光, 同时地面凋落物的积累, 为食草和腐食性昆虫提供丰富的食物资源, 又有利于改善沙地土壤特性, 增加土壤养分含量, 进而改善局地小环境, 而灌丛周围相对沙漠裸地的裸露高温条件, 其相对较低的温度、优良的保水条件可为地表甲虫提供安全的产卵场所[21-27], 这些作用通常被称作灌丛的“肥岛及虫岛效应”。同时, 柽柳包生境又具有较好的地面覆盖层, 能够为甲虫栖息、食物获取、藏身、躲避天敌等活动提供适宜的场所和条件[23]。本研究中地表甲虫在生境选择过程中更趋向于选择地面覆盖物积累较高的柽柳包环境也证实了这一点。植食性的昆虫(象甲类)主要取食柽柳植物及其柽柳包上其他植物, 白天可以观察到这些昆虫取食植物叶片和花;拟步甲除了取食植物之外, 在柽柳包相互追逐完成交配。柽柳包在沙漠地区存在不仅对昆虫物种多样性的保育有着积极的意义, 同时也观察到蜥蜴数量也是明显高于裸地。
本研究中没有系统地监测其他类群昆虫, 但观察到在柽柳开花期间大量的授粉昆虫(食蚜蝇、蜂类和蛾类)访问柽柳包, 需要在以后的研究中加强对授粉昆虫的研究, 通过更严格的控制试验(铲除柽柳包植物、选取死亡的柽柳包, 不同尺度下的比较)来研究柽柳包对生物多样性维持过程及其内在机制。
3.2 柽柳包对地表甲虫庇护作用的强弱有明显的季节变化本研究结果表明地表甲虫的数量和种类存在明显的季节变化, 2005—2006年甲虫数量高峰分别出现在6月和7月(图 1, 图 2)。这可能与两年的气候变化和洪水有很大关系, 2005年降水量相对较少, 地表径流也相对小, 而2006年降水量相对较大, 地表径流也相对较大[28-29], 为躲避洪水的影响或者更好利用土壤水分, 甲虫会涌向柽柳包避难或者种群在洪水后维持相对高的数量, 但洪水与动物种群是十分复杂, 需要在以后的研究注意。Wenninger等也研究表明在夏季末期, 灌溉作用比植物多样性更影响昆虫多样性[5]。本研究表明了在沙漠地区柽柳包对维持地表甲虫群落物种多样性、物种丰富度等有利作用及荒漠地区柽柳包对地表甲虫的庇护作用。人们在开荒屯田过程中, 一定要注重对柽柳包的保护, 以更好维持荒漠地区昆虫多样性和荒漠生态系统的平衡。
| [1] | 高宝嘉, 张执中, 李镇宇. 封山育林对昆虫群落结构及多样性稳定性影响的研究. 生态学报, 1992, 12(1): 1–7. |
| [2] | 于晓东, 周红章, 罗天宏. 鄂尔多斯高原地区昆虫物种多样性研究. 生物多样性, 2001, 9(4): 329–335. |
| [3] | De Alckmin Marques E S, Price P W, Cobb N S. Resource abundance and insect herbivore diversity on woody fabaceous desert plants. Environmental Entomology, 2000, 29(4): 696–703. DOI:10.1603/0046-225X-29.4.696 |
| [4] | Schowalter T D, Lightfoot D C, Whitford W G. Diversity of arthropod responses to host-plant water stress in a desert ecosystem in southern New Mexico. The American Midland Naturalist, 1999, 142(2): 281–290. DOI:10.1674/0003-0031(1999)142[0281:DOARTH]2.0.CO;2 |
| [5] | Wenninger E J, Inouye R S. Insect community response to plant diversity and productivity in a sagebrush-steppe ecosystem. Journal of Arid Environments, 2008, 72(1): 24–33. DOI:10.1016/j.jaridenv.2007.04.005 |
| [6] | Davies K F, Margules C R. Effects of habitat fragmentation on carabid beetles: experimental evidence. Journal of Animal Ecology, 1998, 67(3): 460–471. DOI:10.1046/j.1365-2656.1998.00210.x |
| [7] | 于晓东, 罗天宏, 周红章. 东灵山地区地表甲虫群落组成及季节变化. 昆虫学报, 2002, 45(6): 785–793. |
| [8] | 康乐. 放牧干扰下的蝗虫——植物相互作用关系. 生态学报, 1995, 15(1): 1–11. |
| [9] | 黄保宏, 邹运鼎, 毕守东, 李恒奎. 梅园昆虫群落的时间结构及动态. 应用与环境生物学报, 2005, 11(2): 187–191. |
| [10] | Tengberg A. Nebkha dunes as indicators of wind erosion and land degradation in the Sahel zone of Burkina Faso. Journal of Arid Environments, 1995, 30(3): 265–282. DOI:10.1016/S0140-1963(05)80002-3 |
| [11] | 张荣, 刘彤. 古尔班通古特沙漠南部植物多样性及群落分类. 生态学报, 2012, 32(19): 6056–6066. |
| [12] | Li X R, Chen Y W, Su Y G, Tan H J. Effects of biological soil crust on desert insect diversity: evidence from the Tengger desert of northern China. Arid Land Research and Management, 2006, 20(4): 263–280. DOI:10.1080/15324980600940985 |
| [13] | Tengberg A, Chen D L. A comparative analysis of nebkhas in central Tunisia and northern Burkina Faso. Geomorphology, 1998, 22(2): 181–192. DOI:10.1016/S0169-555X(97)00068-8 |
| [14] | Langford R P. Nabkha (coppice dune) fields of south-central New Mexico, U.S.A. Journal of Arid Environments, 2000, 46(1): 25–41. DOI:10.1006/jare.2000.0650 |
| [15] | Nickling W G, Wolfe S A. The morphology and origin of Nabkhas, region of Mopti, Mali, West Africa. Journal of Arid Environments, 1994, 28(1): 13–30. DOI:10.1016/S0140-1963(05)80017-5 |
| [16] | Brown G, Porembski S. The maintenance of species diversity by miniature dunes in a sand -depleted Haloxylon salicornicum community in Kuwait. Journal of Arid Environments, 1997, 37(3): 461–473. DOI:10.1006/jare.1997.0286 |
| [17] | 刘铭庭. 新疆柽柳属植物研究及推广应用. 中国沙漠, 1996, 16(4): 428–429. |
| [18] | 穆桂金. 塔克拉玛干沙漠灌草丘的发育特征及环境意义. 干旱区研究, 1994, 11(1): 34–41. |
| [19] | 尹传华, 冯固, 田长彦, 白灯莎, 张福锁. 塔克拉玛干沙漠边缘柽柳对土壤水盐分布的影响. 中国环境科学, 2007, 27(5): 670–675. |
| [20] | 尹传华, 冯固, 田长彦, 张福锁. 干旱区柽柳灌丛下土壤有机质、盐分的富集效应研究. 中国生态农业学报, 2008, 16(1): 263–265. |
| [21] | Stapp P. Microhabitat use and community structure of darkling beetles (Coleoptera: Tenebrionidae) in shortgrass prairie: effects of season shrub and soil type. The American Midland Naturalist, 1997, 137(2): 298–311. DOI:10.2307/2426849 |
| [22] | Mazía C N, Chaneton E J, Kitzberger T. Small-scale habitat use and assemblage structure of ground-dwelling beetles in a Patagonian shrub steppe. Journal of Arid Environments, 2006, 67(2): 177–194. DOI:10.1016/j.jaridenv.2006.02.006 |
| [23] | 杭佳, 石云, 刘文惠, 贺达汉. 宁夏黄土丘陵区不同生态恢复生境地表甲虫多样性. 生物多样性, 2014, 22(4): 516–526. |
| [24] | 赵哈林, 郭轶瑞, 周瑞莲. 灌丛对沙质草地土壤结皮形成发育的影响及其作用机制. 中国沙漠, 2011, 31(5): 1105–1111. |
| [25] | 赵哈林, 刘任涛, 周瑞莲, 赵学勇, 张铜会. 科尔沁沙地灌丛的"虫岛"效应及其形成机理. 生态学杂志, 2012, 31(12): 2990–2995. |
| [26] | 苏永中, 赵哈林, 张铜会. 几种灌木、半灌木对沙地土壤肥力影响机制的研究. 应用生态学报, 2002, 13(7): 802–806. |
| [27] | 岳兴玲, 哈斯, 庄燕美, 张军. 沙质草原灌丛沙堆研究综述. 中国沙漠, 2005, 25(5): 738–743. |
| [28] | 孙本国, 沈永平, 王国亚. 1954-2007年叶尔羌河上游山区径流和泥沙变化特征分析. 冰川冻土, 2008, 30(6): 1068–1072. |
| [29] | 高鑫, 张世强, 叶柏生, 谯程骏. 1961-2006年叶尔羌河上游流域冰川融水变化及其对径流的影响. 冰川冻土, 2010, 32(3): 445–453. |