 2017, Vol. 37
2017, Vol. 37文章信息
- 孙越, 何怀江, 李良, 宋彩民, 王峰洁, 夏富才
- SUN Yue, HE Huaijiang, LI Liang, SONG Caimin, WANG Fengjie, XIA Fucai.
- 阔叶红松林下6种早夏草本不同生长期生物量分配及模型构建
- Biomass allocation and biomass allometric models of six early-summer herbs under the canopy of broad-leaved Korean pine forest during different growth periods in Jiaohe, Jilin Province
- 生态学报. 2017, 37(19): 6523-6533
- Acta Ecologica Sinica. 2017, 37(19): 6523-6533
- http://dx.doi.org/10.5846/stxb201607071391
-
文章历史
- 收稿日期: 2016-07-07
- 网络出版日期: 2017-05-27





2. 北京林业大学森林资源与生态系统过程北京市重点实验室, 北京 100083;
3. 吉林省蛟河林业实验区管理局, 吉林 132517
2. The Key Laboratory for Silviculture and Conservation of Ministry of Education, Beijing Forestry University, Beijing 100083, China;
3. Administration Bureau for Jiaohe Forestry Experimental Area, Jilin 132517, China
生物量是度量个体、种群在群落中的地位和功能的重要指标, 同时也是反映群落和生态系统功能强弱的重要指标[1]。植物由根、茎、叶、花、果实、种子等器官组成, 植物个体正常的生长发育依赖于各器官间的协调生长, 各器官生物量在个体生物量中所占的比率代表着同化产物向不同器官的分配比例和生长过程中各器官的协调关系[2-3]。生物量测定可以为植物的生物量积累动态过程、生态系统的物质和能量流动过程提供基础资料[4], 在森林生态系统研究中具有十分重要的意义。
阔叶红松林是东北林区常见的森林类型之一[5]。许多学者对其生物量[6-7]、碳储量[8-10]、群落结构[11-13]、空间格局[14-15]、幼苗更新[16-17]等方面进行过大量研究, 但由于林下层草本植物对森林的群落结构无显著影响, 且其生物量相对较小, 因而在以往的研究中往往被忽略。林下草本植物作为森林生态系统的一个特殊层片, 具有增加物种多样性、保持水土、固碳增汇和促进生态系统养分循环和能量流动等重要作用, 有必要对其进行全面详细的研究。当前对林下草本植物的研究主要集中于多样性[18-19]、分布格局[20]、群落特征[21]等方面, 而对林下草本植物生物量和生物量模型方面的研究较少[22-24]。
根据林下草本植物花期差异, 可以将林下草本植物划分为早春植物、早夏植物和晚夏植物3种类型[25]。早夏植物是指在早春开始萌芽、生长并完成开花, 林分郁闭前后开始结果, 林分郁闭后继续生长的一类植物[22]。国内学者对阔叶红松林林下草本植物的研究主要集中在早春植物, 对其物种多样性、数量特征和群落特征进行了详细的研究[26-29], 而常将早夏植物归纳在林下草本层中而未单独进行研究。在国外, 对早夏植物的研究主要集中在传粉和授粉方面, 而对生物量的研究则局限在根、茎和叶等器官的单独分析[30-32], 缺少对早夏植物整株生物量的系统研究。阔叶红松林林下光照条件在落叶树种展叶前后有显著变化, 早夏植物作为林下的优势草本物种, 对整个生长季林下草本物种多样性和生物量具有重要影响。本文通过对张广才岭地区阔叶红松林林下6种常见早夏草本植物物候特征、不同时期生物量分配特征进行研究, 并构建株高级与各组分生物量模型, 旨在了解早夏草本植物的繁殖策略和生长发育特点, 为进一步评价其在森林生态系统中功能地位和碳汇潜力以及生物多样性保护等方面提供理论依据。
1 研究区概况与研究方法 1.1 研究区概况研究区位于吉林省蛟河林业实验区管理局(地理位置:127°35′—127°51′E, 43°51′—44°05′N)内, 辖区森林面积31820hm2, 保留有大量阔叶红松林。该区属长白山系张广才岭支脉断块中心地貌, 海拔459—517 m。气候属受季风影响的温带大陆性山地气候, 年平均气温为3.8 ℃, 最冷月1月份平均气温为-18. 6 ℃, 最热月7月份平均气温为21.7 ℃。年降水量为700—800 mm, 主要集中在6—8月份。土壤为森林暗棕壤, 平均土层厚度20—100 cm, 富含有机质[13]。
该区植被类型属长白山植物区系, 以温带分布型为主, 植物种类分布复杂。群落内垂直成层现象明显, 乔木树种主要包括红松(Pinus koraiensis)、春榆(Ulmus japonica)、色木槭(Acer mono)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、紫椴(Tilia amurensis)、白牛槭(Acer mandshuricum)、蒙古栎(Quercus mongolica)、大青杨(Populus ussuriensis)、裂叶榆(Ulmus laciniata)、怀槐(Maackia amurensis)等。灌木主要包括簇毛槭(Acer barbinerve)、暴马丁香(Syringa reticulata)、毛榛(Corylus mandshurica)、瘤枝卫矛(Euonymus pauciflorus)、东北鼠李(Rhamnus davurica)、金银忍冬(Lonicera maackii)等[33]。林下草本植物丰富, 在不同季节表现出不同的林下景观, 常见的草本植物有黑水银莲花(Anemone amurensis)、多被银莲花(Anemone raddeana)、朝鲜顶冰花(Gagea lutea var.nakaiana)、荷青花(Hylomecon japonicum)、侧金盏(Adonis amurensis)、美汉草(Meehania fargesii)、北重楼(Paris verticillata)、轮叶百合(Lilium distichum)、白花碎米荠(Cardamine leucantha)、山茄子(Anisodus acutangulus)、鹿药(Smilacina japonica)、紫花变豆菜(Sanicula rubriflora)、东北羊角芹(Aegopodium alpestre)、荨麻(Urtica fissa)、宽叶苔草(Carex siderosticta)等。
1.2 研究方法 1.2.1 样品采集和处理以蛟河天然阔叶红松林林下自然生长的白花碎米荠、北重楼、山茄子、美汉草、鹿药、紫花变豆菜6种早夏草本植物为研究对象。每种选取10株长势良好, 具有代表性的植株做好标记, 从2015年4月1日开始直到生长期结束, 对其进行物候观测并记录, 当10株被观察样本中有3株出现开花时记为花期开始期, 当剩余不超过3株样本花未凋落时记为花期结束期, 按照相同的方法记录果实开始期和果实结束期;生物量调查采用整株收获法, 特别注意的是对多年生草本植物根进行取样时宿根也应全部取出。从4月20日开始到5月30日, 每隔10d采样1次, 从6月初到8月底, 每隔15d采样1次, 每次每种采样20株, 由于不同物种生长期不同, 具体的采样次数及样本量见表 1。样株采集后洗净根部泥土, 用钢尺测量其株高(从植物根部到顶部最高点, 精度为0.1cm), 将其划分为根、根状茎、茎叶、花、果实等组分, 用烘箱在80℃下分别烘干至恒重, 利用电子天平(精度0.0001g)称取各组分生物量干重, 各组分相加得到整株生物量(TB)。
| 物种 Species |
采样次数 Sample times |
采样株数 Number of Sample |
株高 Height/cm |
地上生物量a Above-ground biomass/g |
地下生物量b Below-ground biomass/g |
生殖部分生物量c Reproduction parts biomass/g |
总生物量 Total biomass/g |
根冠比 Root/shoot |
| 白花碎米荠Cardamine leucantha | 11 | 220 | 6.6—70.6 | 0.0815—6.0445 | 0.056—1.6195 | 0.0083—0.6551 | 0.1490—6.6500 | 0.10—5.29 |
| 北重楼Paris erticillata | 8 | 160 | 6.4—51.1 | 0.0240—2.2234 | 0.104—4.3598 | 0.0108—0.0549 | 0.1404—6.5832 | 0.31—5.32 |
| 鹿药Smilacina japonica | 11 | 220 | 4.6—46.5 | 0.0196—2.4228 | 0.0743—5.7031 | 0.0153—0.9986 | 0.1121—7.8069 | 1.17—12.60 |
| 美汉草Meehania fargesii | 11 | 220 | 6.5—68.3 | 0.0678—6.0526 | 0.0709—2.1719 | 0.0151—0.2261 | 0.1773—8.2245 | 0.12—4.71 |
| 山茄子Anisodus acutangulus | 10 | 200 | 8.2—58.3 | 0.0360—3.7846 | 0.0565—3.0366 | 0.0093—0.1267 | 0.0925—6.8212 | 0.18—3.19 |
| 紫花变豆菜Sanicula rubriflora | 10 | 200 | 7.2—87.6 | 0.0836—5.5764 | 0.1534—7.5879 | 0.0038—0.3678 | 0.2494—13.6143 | 0.06—3.21 |
| a:包括茎、叶、花、果实生物量including stems、leaves、flowers and fruits biomass;b:包括地下茎和根生物量including rhizome and roots biomass;c:包括花和果实生物量including flowers and fruits biomass | ||||||||
利用相对生长关系模型对6种植物生长期内地上生物量(AGB)与地下生物量(BGB)进行相关生长分析[34], 模型形式为:
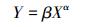
|
(1) |
经对数转化后变为:
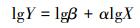
|
(2) |
式中, Y为BGB, X为AGB, β为常数, α为相关生长指数。当α=1时, BGB和AGB为等速生长;当α≠1时, 为异速生长[35]。判断α是否等于1, 利用α的95%置信区间是否包含1来确定。
考虑到同等株高的植株样本因年龄、光照、温湿度以及物候等因素引起的生物量差异, 研究采用4 cm步长(±2 cm)(白花碎米荠、北重楼、美汉草和紫花变豆菜)或2cm步长(±1cm)(鹿药和山茄子)株高分级的平均生物量来构建模型。每种植物选取50株(按照每个株高级内个体数量比例均分分布)作为模型检验样本, 剩余个体作为建模样本。以株高级为自变量, 选取以下五个方程形式构建早夏植物各组分生物量的估测模型, 方程形式为:
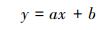
|
(3) |
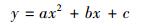
|
(4) |
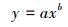
|
(5) |
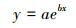
|
(6) |
|
(7) |
式中, y表示各组分生物量(包括TB、AGB和BGB/g), x为株高级(cm), a、b、c为模型参数。
模型建立后, 利用每种剩余的50株作为检验数据对模型精度进行验证, 模型的预估精度利用相对误差(RS)、平均相对误差(EE)、平均相对误差绝对值(RMA)和预估精度(P)进行验证[36], 各估测精度公式为:
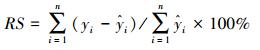
|
(8) |
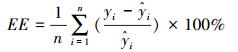
|
(9) |
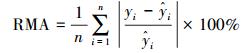
|
(10) |
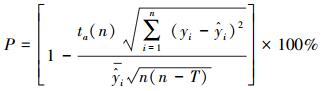
|
(11) |
式中, yi和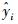
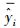
本文采用Excel 2013进行数据处理和制表, 采用R-3.2.3进行统计分析, 采用SigmaPlot 10.0进行制图。
2 结果与分析 2.1 6种早夏植物花、果实物候期由图 1可知, 从6种早夏植物可以看出, 在阔叶红松林下早夏植物花期一般开始于4月底, 结束于6月中旬, 其中紫花变豆菜开花时间最早(4月24日), 北重楼开花时间最晚(5月14日), 紫花变豆菜开花持续时间最长(49 d), 美汉草和山茄子开花持续时间最短(22 d)。6种早夏植物果期开始时间差异明显, 山茄子结果时间最早(5月26日), 其次是美汉草(5月28日)和鹿药(6月6日), 北重楼结果时间最晚(6月20日), 结果开始时间相较于花期开始时间差异较大;从果期持续时间长短来看, 鹿药果期持续时间最长, 长达73d, 美汉草果期最短仅15 d。从整个繁殖期来看, 鹿药的花果期时间最长(104 d), 美汉草的花果期最短(37 d), 6种早夏植物在8月中下旬(8月20日)全部完成生殖过程。

|
| 图 1 6种早夏植物的开花时间, 开花持续时间和结果持续时间 Fig. 1 The Flowering time, Florescence and Fruit period for six early summer herbaceous plants |
表 1列出了6种早夏草本植物整个生长期的采样状况、株高的分布范围、各组分生物量范围及地下/地上生物量(R/S)。从表中可知, 整个生长期内植株高度、生物量以及根冠比变化较大, 6个物种株高最大时是株高最小时的7—10倍;生长末期植株生物量可达生长初期的100倍以上;根冠比最大时也可达到最小时的50倍左右, 且同一物种不同生长期根冠比差异也较大。
6种早夏植物株高随生长期逐渐增加, 随后均出现降低的趋势, 但不同物种降低趋势不同(图 2)。6种早夏植物除紫花变豆菜在7月15日达到株高最大值后出现显著的下降趋势外, 其它5种均在6月30日达到株高的最大值, 随后株高均有轻微的降低趋势;6种植物在生长初期株高无明显差异, 随着生长期增长物种间株高差异增大, 其中紫花变豆菜和白花碎米荠株高最大, 其次是山茄子和美汉草, 最小的是北重楼和鹿药。6种植物在5月30日之前, 株高增长率达到最大后逐渐下降, 说明在林分内落叶树种未展叶前, 林下光照资源充足, 早夏植物会快速进行高生长, 在此期间主要进行营养生长和完成开花过程;林分内所有树种完全展叶后(6月30日前后)后, 光照成为最主要的限制因子, 此时植物的高生长停止, 在此过程中地上部分会完成结实并逐渐枯萎死亡。

|
| 图 2 不同生长时期6种早夏草本植物株高、根冠比及生物量变化 Fig. 2 The height, R/S and biomass variation of six early-summer herbs with growth periods |
6种植物的总生物量与株高随生长期的变化表现出大致相同的变化趋势(图 2), 但不同的是总生物量在达到最大值后会随着生长期的增加呈现显著的降低趋势。在整个生长阶段, 紫花变豆菜具有最高的单株总生物量, 显著高于其它5种;其它5种差异相对较小, 在生长早期阶段(6月15日之前)北重楼总生物量最小, 后期(6月15日之后)鹿药总生物量最小。
6种植物繁殖器官(花、果实)生物量在整个生长阶段均表现出不同的趋势(图 2), 说明6种植物具有不同的繁殖策略, 这些繁殖策略是植物对环境的长期适应的结果。具体来看, 紫花变豆菜最早(4月30日)出现繁殖器官生物量的积累随后生物量逐渐增大, 在6月15日出现显著的增大趋势, 在6月30日达到最大随后显著降低, 7月15日后凋落, 说明紫花变豆菜在6月15日后花逐渐转变为果实, 6月30日果实基本成熟随后开始凋落, 这与其花、果实的物候期是相对应的。山茄子、白花碎米荠、美汉草和鹿药4种在5月10日出现花, 其中山茄子在整个期间繁殖器官生物量无明显变化趋势;白花碎米荠与紫花变豆菜表现出相似趋势;美汉草在整个繁殖过程中繁殖器官生物量逐渐增大, 5月30日后即不再出现, 说明白花碎米荠果实在很短时间内就能够成熟并凋落;而鹿药的繁殖器官持续时间最长, 其生物量在7月15日之前表现为缓慢增长趋势, 7月15日后显著增加直到8月15日后凋落。北重楼出现繁殖器官生物量积累时间最晚, 在6月15日时才开始出现, 并且凋落的时间最早, 在6月30日后即凋落, 说明北重楼会在极短时间内完成整个生殖过程, 这种繁殖策略可能是为了避免较长时间的生殖过程带来的外界干扰, 而在极短时间内完成整个生殖过程会大大降低外界对其造成伤害的几率。
6种植物R/S随着不同的生长时期表现出不同的变化趋势(图 2), 其中白花碎米荠、美汉草、山茄子和紫花变豆菜表现为生长初期最大, 随着生长时间逐渐降低, 到5月30日后基本保持不变;北重楼的R/S生长初期和生长末期基本保持不变, 整个生长期表现为偏“V”字型, 且地下生物量高于地上生物量(R/S>1);鹿药R/S表现为生长末期显著高于生长初期, 虽然在整个生长阶段会出现忽高忽低的现象, 但整体是逐渐增高的趋势, 这主要是由于鹿药具有明显的根状茎, 林分郁闭后植株地上生物量生长速率降低而根状茎会继续生长, 从而造成生长末期R/S较大。
2.3 不同时期6种早夏草本植物生物量分配及地上、地下部分相关生长关系从6种草本植物各组分生物量分配比例来看(图 3), 营养部分和地下部分所占比例远远大于繁殖部分, 繁殖部分生物量分配比例在整个生长期内维持较低水平;不同物种各组分生物量分配比例存在一定差异, 且同一物种在不同生长期生物量分配比例有所不同。所有物种不同生长期内繁殖部分生物量比例均较小(不超过5%), 说明在整个生长期内早夏草本植物用于开花、结实的生物量是极少的。除鹿药和北重楼营养部分生物量分配比例较低外(不超过40%), 其它4种均较高, 特别是在繁殖期营养部分生物量比例大都超过60%, 随后会保持不变或稍有降低。从同一物种不同生长期来看, 大都表现为生长初期营养生长部分较低随后增大到繁殖期后又稍降低或保持不变的趋势。而地下部分生物量分配比例与营养部分成相反的规律。

|
| 图 3 6种早夏草本植物不同生长期各组分生物量分配比例 Fig. 3 The biomass allocation proportion of six early-summer herbs plant with growth periods 营养部分包括茎和叶;繁殖部分包括花和果实;地下部分包括地下茎和根 |
从表 2可知, 6种早夏草本植物地下、地上生物量之间均具有明显的相关生长关系(P < 0.0001), 且均表现为异速生长关系。从相关生长指数α和其95%的置信区间来看, 白花碎米荠、美汉草、山茄子和紫花变豆菜的相关生长指数α均小于1, 同时其95%置信区间上限也均小于1, 说明这4种植物地下部分生长速率小于地上部分;而北重楼和鹿药相关生长指数α和95%置信区间下限均大于1, 说明这两种植物地下生长速率要高于地上部分, 这可能是由于北重楼和鹿药均具有根状茎存在, 根状茎和根的共同生长速率超过了地上部分的增长速率, 特别是在生长后期, 地上生物量的增加量降低而地下根状茎为下一年无性繁殖会快速储存营养物质和能量, 从而导致地下部分增长速率较高。
| 物种 Species |
样本量 Number of samples |
相关生长指数 Allometric scaling exponent α |
截距Intercept lgβ | R2 | P | F | 生长关系 Allometric relationships |
|||
| 平均值±标准误差 Mean±SD |
95%置信区间 95% confidence interval |
平均值±标准误差 mean±SD |
95%置信区间 95% confidence interval |
|||||||
| 白花碎米荠 | 220 | 0.1755±0.0148 | 0.1463—0.2047 | 0.2539±0.0260 | 0.2027—0.3051 | 0.392 | < 0.0001 | 140.5 | 异速生长 | |
| 北重楼 | 160 | 1.9127±0.0731 | 1.7683—2.0571 | 0.0868±0.0536 | -0.0190—0.1927 | 0.8124 | < 0.0001 | 684.4 | 异速生长 | |
| 鹿药 | 220 | 2.4187±0.1108 | 2.2004—2.6370 | 0.4800±0.0845 | 0.3135—0.6466 | 0.6862 | < 0.0001 | 476.6 | 异速生长 | |
| 美汉草 | 220 | 0.2638±0.0171 | 0.2301—0.2974 | 0.2572±0.0264 | 0.2052—0.3092 | 0.5225 | < 0.0001 | 238.6 | 异速生长 | |
| 山茄子 | 200 | 0.5915±0.0286 | 0.5351—0.6480 | 0.0058±0.0462 | -0.0852—0.0967 | 0.6835 | < 0.0001 | 427.7 | 异速生长 | |
| 紫花变豆菜 | 200 | 0.4754±0.0245 | 0.4261—0.5247 | 0.5906±0.0718 | 0.4489—0.7322 | 0.6463 | < 0.0001 | 361.7 | 异速生长 | |
以株高级为自变量, 利用方程(3)—(7) 构建了AGB、BGB和TB的估测模型, 并根据模型的SEE和R2选择最佳模型(选择标准为SEE较小, 且R2较大)。模型形式及其参数见表 3。
| 物种 Species |
组分 Components |
最佳模型 Optimal model |
模型参数Model parameter | R2 | SEE | P | CF* | ||
| a** | b** | c** | |||||||
| 白花碎米荠 C. leucantha |
AGB | y=ax2+bx+c | 0.0014 | -0.045 | 0.666 | 0.986 | 0.176 | < 0.001 | — |
| BGB | y=ax2+bx+c | 0.0005 | -0.019 | 0.447 | 0.906 | 0.128 | < 0.001 | — | |
| TB | y=ax2+bx+c | 0.0018 | -0.064 | 1.112 | 0.981 | 0.266 | < 0.001 | — | |
| 北重楼 P. verticillata |
AGB | y=axb | 0.0029 | 1.637 | — | 0.982 | 0.135 | < 0.001 | 1.009 |
| BGB | y=axb | 0.0057 | 1.647 | — | 0.977 | 0.154 | < 0.001 | 1.012 | |
| TB | y=axb | 0.0087 | 1.643 | — | 0.984 | 0.130 | < 0.001 | 1.008 | |
| 鹿药 S. japonica |
AGB | y=axb | 0.0014 | 1.918 | — | 0.993 | 0.114 | < 0.001 | 1.006 |
| BGB | y=axb | 0.0067 | 1.765 | — | 0.968 | 0.225 | < 0.001 | 1.026 | |
| TB | y=axb | 0.0080 | 1.805 | — | 0.985 | 0.157 | < 0.001 | 1.012 | |
| 美汉草 M. urticifolia |
AGB | y=axb | 0.0133 | 1.286 | — | 0.949 | 0.195 | < 0.001 | 1.019 |
| BGB | y=aebx | 0.2783 | 0.020 | — | 0.737 | 0.233 | < 0.001 | 1.028 | |
| TB | y=axb | 0.0536 | 1.009 | — | 0.905 | 0.214 | < 0.001 | 1.023 | |
| 山茄子 B. paridiformis |
AGB | y=axb | 0.0015 | 1.907 | — | 0.917 | 0.321 | < 0.001 | 1.053 |
| BGB | y=axb | 0.0054 | 1.402 | — | 0.851 | 0.329 | < 0.001 | 1.056 | |
| TB | y=axb | 0.0057 | 1.660 | — | 0.917 | 0.280 | < 0.001 | 1.040 | |
| 紫花变豆菜 S. rubriflora |
AGB | y=aebx | 0.4523 | 0.034 | — | 0.952 | 0.187 | < 0.001 | 1.018 |
| BGB | y=aebx | 0.6580 | 0.020 | — | 0.700 | 0.321 | < 0.001 | 1.053 | |
| TB | y=axb | 0.1373 | 0.913 | — | 0.768 | 0.346 | < 0.001 | 1.062 | |
| 混合物种 all species |
AGB | y=axb | 0.0035 | 1.656 | — | 0.901 | 0.359 | < 0.001 | 1.067 |
| BGB | y=axb | 0.0314 | 0.981 | — | 0.443 | 0.721 | < 0.001 | 1.297 | |
| TB | y=axb | 0.0222 | 1.324 | — | 0.780 | 0.461 | < 0.001 | 1.112 | |
| AGB:地上生物量above-ground biomass;BGB:地下生物量below-ground biomass;TB:整株生物量total biomass;SEE:估计标准误standard error estimate;CF:校正因子correction factor; *为校正系数, 计算公式为:CF=exp(SEE2/2);**在P < 0.001水平下均差异显著; —表示无数据 | |||||||||
从表 3可知, 5种模型中有幂函数方程、一元二次方程和指数方程3种模型被选用, 其中以幂函数形式使用次数最多, 其次是指数方程形式和一元二次方程。除混合物种BGB方程R2较低外(0.443), 其它方程R2均大于0.70且SEE均较低, 说明模型拟合效果较好, 株高级可以很好的表征6种草本植物生物量。从6个物种不同组分生物量来看, AGB方程的拟合效果最好R2均大于0.900;TB方程的拟合效果也较好R2介于0.768—0.985之间, 两者均要好于BGB(R2介于0.443—0.977), 这可能是由于地下生物量在获取过程中会有损失存在较大的不确定性造成的。从单种模型和混合物种模型来看, 混合物种模型效果相对较差(特别是BGB), 说明6个物种不同种间株高(株高级)与生物量间存在较大差异, 在估测物种生物量时尽量利用单种生物量方程进行计算。
将预留的每种50株样本株高的实测数据转化为株高级带入到各自最佳模型中(混种模型把6种300株样本株高转化为株高级后带入), 得出各组分生物量的预测值, 根据公式(6)—(9) 计算得出各拟合方程的总相对误差(RS)、平均相对误差(EE)、平均相对误差绝对值(RMA)、预估精度(P), 结果见表 4.从检验结果来看, 所有方程的RS、EE和RMA分别介于-9.465%—3.405%、-12.459%—6.180%和10.470%—30.697%之间, 各模型相对误差均较小, 所有方程的预估精度P值均大于80%。
| 物种 Species |
组分 Components |
最佳模型 Optimal models |
RS/% | EE/% | RMA/% | P/% |
| 白花碎米荠 C. leucantha |
AGB | y=1.4×10-3x2-0.045x+0.666 | -7.370 | -3.252 | 19.864 | 89.995 |
| BGB | y=4.5×10-4x2-0.019x+0.447 | -8.896 | -5.096 | 10.470 | 91.032 | |
| TB | y=1.8×10-3x2-0.064x+1.112 | 3.405 | 4.071 | 14.720 | 91.869 | |
| 北重楼P.verticillata | AGB | y=2.9×10-3x1.637 | 0.877 | -5.909 | 14.472 | 89.789 |
| BGB | y=5.8×10-3x1.647 | -1.395 | -3.548 | 13.644 | 90.345 | |
| TB | y=8.7×10-3x1.643 | -1.562 | -5.238 | 11.286 | 92.418 | |
| 鹿药S. japonica | AGB | y=1.4×10-3x1.918 | 1.611 | 4.173 | 14.074 | 91.779 |
| BGB | y=6.8×10-3x1.765 | 1.297 | -1.191 | 18.578 | 90.196 | |
| TB | y=8.1×10-3x1.805 | 0.534 | -1.158 | 15.435 | 90.669 | |
| 美汉草M. urticifolia | AGB | y=1.4×10-3x1.286 | -9.465 | -9.922 | 16.519 | 87.373 |
| BGB | y=0.286×e0.020x | 2.597 | 1.518 | 10.943 | 91.306 | |
| TB | y=5.5×10-2x1.009 | -5.563 | -5.324 | 13.591 | 91.253 | |
| 山茄子 B. paridiformis |
AGB | y=1.5×10-3x1.907 | -8.083 | -12.459 | 14.147 | 89.683 |
| BGB | y=5.7×10-3x1.402 | -3.559 | -4.101 | 14.055 | 90.498 | |
| TB | y=5.9×10-3x1.660 | -4.508 | -7.634 | 12.766 | 91.278 | |
| 紫花变豆菜 S. rubriflora |
AGB | y=0.460×e0.034x | -3.776 | -0.768 | 24.242 | 88.834 |
| BGB | y=0.693×e0.020x | -0.121 | 1.650 | 14.273 | 90.690 | |
| TB | y=0.146×e0.913x | -2.921 | -0.895 | 24.778 | 83.406 | |
| 混合物种 All species |
AGB | y=3.7×10-3x1.656 | 1.963 | 6.180 | 28.160 | 93.782 |
| BGB | y=4.1×10-2x0.981 | -0.991 | 0.272 | 30.697 | 91.339 | |
| TB | y=2.5×10-2x1.324 | -4.198 | -2.194 | 25.029 | 93.775 | |
| RS:相对误差relative error; EE:平均相对误差mean relative error; RMA:平均相对误差绝对值mean absolute relative error; P:预估精度prediction precision | ||||||
植物体不同器官生物量分配策略是植物对环境条件长期自然选择的结果, 植物通过调节自身资源配置以适应复杂的环境变化, 在不同的环境条件下表现出不同的生长特性和生物量分配规律[37-38]。生物量分配策略使植物对环境条件的利用达到最大化, 但不同类型物种生物量分配模式存在较大差异[39], 东北阔叶红松林林下早夏草本植物为了获得更多的光照资源一般表现为具有更高的地上生物量(图 3), 但在不同的生长阶段, 植物各器官生物量的分配规律不同, 如何调整各器官生物量分配规律的变化是研究植物生存策略的重要内容[37]。从本研究可知, 6种植物中4种(白花碎米荠、美汉草、山茄子和紫花变豆菜)表现为生长初期R/S最大(R/S大于1), 并随着植物生长R/S逐渐降低并在中后期趋于稳定(R/S小于1), 从各组分生物量分配比例也可知, 这4种植物生长早期营养部分比例较地下部分低, 随着植物的生长, 营养部分逐渐增大到整株植物的60%—80%。这是由于4种植物均属于多年生草本, 生长初期地下部分存在上一年保留的老根而地上部分刚开始萌发生长, 地上生物量相对较低, 同时也是植物为了获取更多的水分、养分等地下资源, 把更多的生物量分配到地下部分, 以便释放随之而来的生存压力[37];随着落叶树种展叶林冠闭合, 林下空间和光照资源缺乏, 植物把更多的生物量分配到地上部分争夺空间和光照资源, 以满足自身生长和繁殖需要。北重楼和鹿药两种植物的分配规律与其它4种不同, 在整个生长季的R/S均大于1, 北重楼呈“V”型分布, 鹿药呈随生长时期逐渐升高的趋势。这可能是由于这两种植物有明显的根状茎, 随着植株生长根状茎同时生长, 根状茎在储存能量的同时可以进行无性繁殖, 为了维持生长和繁殖会把更多的生物量分配在地下部分, 同时在林冠闭合后, 地上生长停止而地下根状茎会继续增加为下一年植物的生长和繁殖作准备[23]。
根据代谢理论[40]和维管网络模型[35]植物器官生物量间存在相对生长关系(幂指数关系), 相对生长关系与个体大小(尺度)无关, 可以揭示不同物种间生物量分配的基本速率问题[34-35]。对全球植物AGB和BGB数据库(包括不同生境条件下的草本与木本植物)的研究表明, 自然条件下种内和种间个体水平的AGB和BGB分配速率是等速的(即幂指数a为1.0, 为等速生长关系)[34-35]。但本研究的6种植物AGB和BGB之间的生长关系均是异速的(所有幂函数指数a均不等于1), 可能是由于研究对象所处的生长阶段不同造成的, 本研究是对植物从幼苗到成熟整个生长阶段进行的, 表征的是整个生长期内AGB和BGB的平均水平, 而Nnquist&Niklas[35]和Niklas[34]的研究对象是成熟植株。肖遥等[37]对4种荒漠草本植物不同生长期的生物量分配进行研究, 得出不同生长期4种植物相关生长指数是动态变化的, 且变化趋势均不同, 说明在不同的生长阶段AGB和BGB表现出不同的生长关系(等速和异速交替出现)。所以为了解释整个生长阶段植物的相对生长关系, 应进一步对每个物种不同生长阶段的AGB和BGB的生长速率进行分析研究。
3.2 生物量模型的建立和检验生物量模型方法是基于易测变量评价生物量的较准确方法[41], 对草本植物而言, 株高是较为易测的因子之一, 可以利用株高(或转化为株高级)作为变量建立草本植物生物量方程。有研究利用地径和冠幅作为自变量进行拟合生物量方程[41], 但本研究中6种草本植物地径较小且在整个生长期变化不明显, 用地径来拟合方程效果不好, 而冠幅测量相对麻烦且同一生长期冠幅变化差异较大也不适宜用来拟合方程, 所以本研究选用株高(株高级)作为唯一变量进行生物量模型的拟合。从研究结果可知, 利用株高级建立单种地上生物量、地下生物量和总生物量方程的R2均较高且SEE较低, 说明株高(或株高级)可以很好的表征草本植物各组分生物量。以往许多研究也常会利用株高或株高级作为变量建立草本植物生物量方程[23-24, 42]。利用不同模型形式建立生物量模型, 不同物种所选择的最优模型形式不同。本研究中以幂函数模型被选用次数最多, 不管是对乔木、灌木植物还是对草本植物进行生物量模型构建时[43-44], 这种模型形式经常被用到。相比于单种生物量模型, 混种模型拟合效果相对较差, 这是因为6种草本植物自身属性差异较大(属于不同的科属), 造成株高生长速率存在差异, 按照单一变量(株高级)建立生物量方程势必会增大预测误差, 拟合效果相对较差。相比于地上生物量和总生物量, 地下生物量拟合效果较差, 可能是因为地下生物量在挖取和清洗过程中会造成细根的损失, 存在较大的不确定性。对乔木和灌木建立地下生物量模型时, 也经常会遇到这一问题[43-44]。
有研究表明, 当模型验证的RS, EE和RMA<30%, P>80%时, 模型拟合结果比较符合实际值[36, 45]。从本研究结果来看, 除混合物种BGB方程的RMA(30.679%)稍大于30%外, 其它所有方程的RS、EE和RMA均小于30%, 且所有P值均大于80%, 说明所选择的最优模型符合建模的标准可以用来估测当地各物种的生物量。混种模型对BGB的估测误差相对较大但基本符合建模要求, 在无单种生物量方程的情况下, 可以选用混种模型估测草本物种生物量。本研究6种早夏植物单种生物量模型和混种模型是在阔叶红松林林下进行取样建立的, 是否适用于其它林分或者非林地还有待进一步验证。使用本研究中生物量方程进行生物量估算前, 应进行验证符合标准后再使用, 否则要对模型进行校正后再使用。
| [1] | Odum E P, Barrett G W. Fundamentals of Ecology. Philadelphia: Saunders, 1971. |
| [2] | 苏智先, 钟章成. 缙云山慈竹种群生物量结构研究. 植物生态学与地植物学学报, 1991, 15(3): 240–252. |
| [3] | 孙儒泳, 李庆芬, 牛翠娟, 娄安如. 基础生态学. 北京: 高等教育出版社, 2002. |
| [4] | 李雪华, 李晓兰, 蒋德明, 骆永明, 王红梅. 科尔沁沙地70种草本植物个体和构件生物量比较研究. 干旱区研究, 2009, 26(2): 200–205. |
| [5] | 张春雨, 赵秀海, 赵亚洲. 长白山温带森林不同演替阶段群落结构特征. 植物生态学报, 2009, 33(6): 1090–1100. |
| [6] | 杨松, 侯继华, 赵秀海, 魏丽萍, 李颖, 姚静, 代海军, 何怀江. 吉林蛟河天然阔叶红松林下5种灌木生物量估算模型. 广东农业科学, 2013(9): 36–41. |
| [7] | 代海军, 何怀江, 赵秀海, 张春雨, 汪金松, 杨松. 阔叶红松林两种主要树种的生物量分配格局及异速生长模型. 应用与环境生物学报, 2013, 19(4): 718–722. |
| [8] | 范春楠. 吉林省森林植被碳估算及其分布特征[D]. 哈尔滨: 东北林业大学, 2014: 137. http://cdmd.cnki.com.cn/Article/CDMD-10225-1014410161.htm |
| [9] | 齐麟, 于大炮, 周旺明, 周莉, 赵福安, 王长海, 代力民. 采伐对长白山阔叶红松林生态系统碳密度的影响. 生态学报, 2013, 33(10): 3065–3073. |
| [10] | 王新闯, 齐光, 于大炮, 周莉, 代力民. 吉林省森林生态系统的碳储量、碳密度及其分布. 应用生态学报, 2011, 22(8): 2013–2020. |
| [11] | 徐丽娜, 金光泽. 小兴安岭凉水典型阔叶红松林动态监测样地:物种组成与群落结构. 生物多样性, 2012, 20(4): 470–481. |
| [12] | 李良, 夏富才, 杨晓燕, 张显龙, 孙国文, 吴相菊, 王贵春, 林天喜. 张广才岭阔叶红松林林分结构研究. 林业资源管理, 2014(1): 56–61. |
| [13] | 何怀江, 张春雨, 赵秀海, 倪瑞强, 高海涛, 王贵春. 吉林蛟河次生针阔混交林群落结构与微地形关系. 东北林业大学学报, 2014, 42(5): 13-17, 33-33. |
| [14] | 赵中华, 惠刚盈, 胡艳波, 周红敏, 张连金. 2种类型阔叶红松林优势种群空间分布格局及其关联性. 林业科学研究, 2011, 24(5): 554–562. |
| [15] | 王蕾, 张春雨, 赵秀海. 长白山阔叶红松林的空间分布格局. 林业科学, 2009, 45(5): 54–59. DOI:10.11707/j.1001-7488.20090508 |
| [16] | 陈香茗, 赵秀海, 夏富才, 王蕾, 陈贝贝. 长白山紫椴种子雨的时空分布格局. 东北林业大学学报, 2011, 39(1): 7–10. |
| [17] | 姚杰, 闫琰, 张春雨, 邳田辉, 赵秀海. 吉林蛟河针阔混交林乔木幼苗组成与月际动态. 植物生态学报, 2015, 39(7): 717–725. DOI:10.17521/cjpe.2015.0068 |
| [18] | 李步杭, 张健, 姚晓琳, 叶吉, 王绪高, 郝占庆. 长白山阔叶红松林草本植物多样性季节动态及空间分布格局. 应用生态学报, 2008, 19(3): 467–473. |
| [19] | 夏富才, 赵秀海, 潘春芳, 赵晓静, 王玉红. 长白山红松阔叶林草本植物物种多样性季节动态. 吉林农业大学学报, 2009, 31(1): 27-31, 44-44. |
| [20] | 张姗, 蔺菲, 原作强, 匡旭, 贾仕宏, 王芸芸, 索炎炎, 房帅, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林草本层物种多度分布格局及其季节动态. 生物多样性, 2015, 23(5): 641–648. DOI:10.17520/biods.2015089 |
| [21] | 夏富才, 张春雨, 赵秀海, 潘春芳. 早春草本植物群落结构及其聚类分析. 东北师大学报:自然科学版, 2008, 40(4): 109–114. |
| [22] | 李良, 孙悦, 夏富才, 王戈戎, 张英莉, 张骁. 轮叶百合生物量分配分析. 北方园艺, 2015(1): 87–89. |
| [23] | 范春楠, 程岩, 郑金萍, 刘彬, 刘鹤. 早春类短命植物生物量研究(Ⅰ)——齿瓣延胡索生物量特征及模型构建. 北华大学学报:自然科学版, 2015, 16(6): 790–795. |
| [24] | 范春楠, 程岩, 郑金萍, 杨学东, 王月. 早春类短命植物生物量研究(Ⅱ)——多被银莲花生物量及其分配特征. 北华大学学报:自然科学版, 2016, 17(1): 24–29. |
| [25] | Kudo G, Ida T Y, Tani T. Linkages between phenology, pollination, photosynthesis, and reproduction in deciduous forest understory plants. Ecology, 2008, 89(2): 321–331. DOI:10.1890/06-2131.1 |
| [26] | 吴刚, 尹若波, 周永斌, 郝占庆, 梁秀英. 长白山红松阔叶林林隙动态变化对早春草本植物的影响. 生态学报, 1999, 19(5): 659–663. |
| [27] | 叶万辉, 姜会武, 董世林. 不同林型下早春植物层片生物量的分析研究. 哈尔滨师范大学自然科学学报, 1992, 8(4): 87–95. |
| [28] | 赵秀海, 戚继忠, 孙广仁, 刘青山, 陈喜林, 于民. 长白山阔叶红松林早春草本层植物的数量特征. 吉林林学院学报, 1997, 13(1): 7-11, 16-16. |
| [29] | 周永斌, 殷有, 于凤英, 郝占庆, 叶吉. 长白山阔叶红松林早春植物的养分吸收与归还. 生态学杂志, 2007, 26(6): 793–796. |
| [30] | Helenurm K, Barrett S C H. The reproductive biology of boreal forest herbs. Ⅱ. Phenology of flowering and fruiting. Canadian Journal of Botany, 1987, 65(10): 2047–2056. DOI:10.1139/b87-279 |
| [31] | Ida T Y, Kudo G. Seasonal patterns of carbon assimilation and allocation of a summer-green forest herb, Parasenecio auriculata (Senecioneae; Asteraceae). Plant Ecology, 2010, 210(1): 181–193. DOI:10.1007/s11258-010-9748-3 |
| [32] | Bonser S P, Aarssen L W. Allometry and development in herbaceous plants: functional responses of meristem allocation to light and nutrient availability. American Journal of Botany, 2003, 90(3): 404–412. DOI:10.3732/ajb.90.3.404 |
| [33] | 丁胜建, 张春雨, 夏富才, 赵秀海, 倪瑞强, 范娟, 何怀江. 老龄阔叶红松林下层木空间分布的生境关联分析. 生态学报, 2012, 32(11): 3334–3342. |
| [34] | Niklas K J. Modelling below-and above-ground biomass for non-woody and woody plants. Annals of Botany, 2005, 95(2): 315–321. DOI:10.1093/aob/mci028 |
| [35] | Enquist B J, Niklas K J. Global allocation rules for patterns of biomass partitioning in seed plants. Science, 2002, 295(5559): 1517–1520. DOI:10.1126/science.1066360 |
| [36] | 唐守正, 张会儒, 胥辉. 相容性生物量模型的建立及其估计方法研究. 林业科学, 2000, 36(S1): 19–27. |
| [37] | 肖遥, 陶冶, 张元明. 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征. 植物生态学报, 2014, 38(9): 929–940. |
| [38] | 何玉惠, 赵哈林, 刘新平, 张铜会, 岳广阳. 不同类型沙地狗尾草的生长特征及生物量分配. 生态学杂志, 2008, 27(4): 504–508. |
| [39] | 郝虎东, 田青松, 石凤翎, 卞晓燕, 李芳. 无芒雀麦地上生物量及各构件生物量分配动态. 中国草地学报, 2009, 31(4): 85–90. |
| [40] | West G B, Brown J H, Enquist B J. A general model for the structure and allometry of plant vascular systems. Nature, 1999, 400(6745): 664–667. DOI:10.1038/23251 |
| [41] | Oliveras I, van der Eynden M, Malhi Y, Cahuana N, Menor C, Zamora F, Haugaasen T. Grass allometry and estimation of above-ground biomass in tropical alpine tussock grasslands. Austral Ecology, 2014, 39(4): 408–415. DOI:10.1111/aec.2014.39.issue-4 |
| [42] | 李良, 孙悦, 夏富才, 王戈戎, 张英莉. 美汉草(Meehania fargesii Levl)生物量分配分析. 北华大学学报:自然科学版, 2014, 15(6): 804–807. |
| [43] | Rojas-García F, De Jong B H J, Martínez-Zurimendí P, Paz-Pellat F. Database of 478 allometric equations to estimate biomass for Mexican trees and forests. Annals of Forest Science, 2015, 72(6): 835–864. DOI:10.1007/s13595-015-0456-y |
| [44] | Conti G, Enrico L, Casanoves F, Díaz S. Shrub biomass estimation in the semiarid Chaco forest: a contribution to the quantification of an underrated carbon stock. Annals of Forest Science, 2013, 70(5): 515–524. DOI:10.1007/s13595-013-0285-9 |
| [45] | 范春楠, 庞圣江, 郑金萍, 李兵, 郭忠玲. 长白山林区14种幼树生物量估测模型. 北京林业大学学报, 2013, 35(2): 1–9. |