 2017, Vol. 37
2017, Vol. 37文章信息
- 高韶勃, 刘磊, 王宇坤, 李静鹏, 赵念席, 高玉葆
- GAO Shaobo, LIU Lei, WANG Yukun, LI Jingpeng, ZHAO Nianxi, GAO Yubao.
- 不同利用方式下内蒙古典型草原群落物种相互关系的年际变化
- The annual variation of effects under different grassland utilization types on typical steppe species interactions in Inner Mongolia
- 生态学报. 2017, 37(19): 6562-6570
- Acta Ecologica Sinica. 2017, 37(19): 6562-6570
- http://dx.doi.org/10.5846/stxb201607011355
-
文章历史
- 收稿日期: 2016-07-01
- 网络出版日期: 2017-05-27

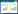




植物群落的空间分布格局是由过去某些生态过程(包括生物过程, 如被捕食, 和非生物过程, 如气候、地形)引发的结果[1-3], 是由一系列复杂植物间相互作用的生态过程所引起[4], 竞争(植物间负相互作用)曾被认为是在高生产力群落当中决定群落结构和多样性的重要驱动因子[5], 并且群落当中竞争作用可以减少竞争者的丰富度, 产生群落中物种之间的负相互关联[6-8], 然而, 竞争对群落结构的重要性, 会随着环境胁迫程度的改变而改变, 在某些环境条件下, 群落当中植物间正相互作用(促进)的影响对群落结构起重要作用[9]。早期的关于植物群落空间格局的实验以邻体移除实验为主, 大多都仅仅关注一对或几对物种对之间的相互关系, 而没有把研究对象放在群落整体的相互作用上。随着研究的深入, 通过物种之间关系来间接地推断其可能潜在影响群落结构及物种多样性的小尺度空间格局关联方法应运而生[10], 这种方法可在一个较小的空间范围内, 通过小尺度空间格局实验, 研究可能发生相互作用的物种之间的关系, 可以有效地对群落整体的物种相互关系进行一个全面的了解, 并且还能够通过已有观测数据来预测将来某一特定时间段群落格局的变化[4], 现已广泛应用于植物群落的格局分析[11-12]。
由于全球气候变化以及20世纪60年代以来长期的过度放牧等不合理利用, 我国北方重要的生态屏障——内蒙古典型草原已发生大面积退化, 其表现为优势种的更替, 物种多样性的减少以及草地生态系统地上净初级生产力的降低。其中, 作为该区域最有代表性的大针茅(Stipa grandis)为建群种的草原群落正在逐步演替为以克氏针茅(Stipa krylovii)和冷蒿(Artemisia frigida)为建群种的草原群落, 群落结构发生了明显变化[13]。自由放牧和刈割草场是内蒙古草原区两种主要的利用方式[14], 在探讨利用方式对群落结构影响的研究方面, 我国学者在物种对于利用方式的响应及物种适应对策等方面做了很多工作[15], 但利用小尺度空间格局法来探索土地利用方式对物种相互关系甚至整个群落格局影响的研究才刚刚起步[16], 而相关研究不仅可以了解和预测群落格局变化, 而且对指导草原的合理利用和有效保护具有重要的理论及实践意义。
因此, 本研究在大针茅草原广泛分布的锡林郭勒草原选择放牧和刈割两种利用方式的样地, 经过3年连续的定点观测群落内小尺度格局, 一方面探讨不同利用方式对大针茅群落植物间相互作用的影响;另一方面探讨群落相互作用关系是否受年际水热变化的影响;所得结果不仅能合理预测利用方式及气候变化对草原演替的影响, 而且可对气候变化下草原的合理利用与有效保护措施的制定提供参考。
1 材料与方法 1.1 研究区域自然概况本研究位于内蒙古自治区锡林郭勒盟锡林浩特市东南55 km处的毛登牧场2.87万hm2生态保护区(116°20.552′E, 44°16.062′N), 海拔910—1377m, 属于典型草原区, 群落年最高高度可达70cm, 盖度约为25%—40%。该区域地形南高北低, 坡度小于10°。利用类型分为放牧(围栏外)与刈割(围栏内)两种方式, 其中放牧方式为夏秋季放牧, 放牧强度为每公顷4只羊, 刈割类型为每年8月中下旬刈割1次。气候类型为典型的大陆性温带草原气候, 四季交替明显, 雨热同期, 冬季寒冷干燥, 夏季较为温暖, 多风, 年平均气温为-0.5℃, 最高气温37.4℃, 最低气温-39.9℃。年平均降雨量280—360mm, 多集中在6—8月, 年蒸发量为降雨量的5.8倍, 年日照约为2800h, 无霜期90—115d。
1.2 研究方法分别在围栏内外设置100m长样线, 沿样线方向, 每隔10m设置调查样方, 每个样方大小为0.5m×0.5m, 然后将样方等分为25个0.1m×0.1m的小样方, 记录每个样方中出现的每种物种, 共计500个小样方。为了便于查找, 每个样方(0.5m×0.5m)的4个顶点用喷过红色漆的筷子定位, 将筷子打进土壤表层15cm, 外露土壤表面1cm, 并用GPS仪对每个样方的经纬度记录以便于定点观测。调查记录时间分别为2013年7月22日, 2014年7月27日和2015年7月24日。植物群落数据于2013年7月测定, 分别测定群落物种数、高度、盖度、生物量(1m2), 土壤样品取自群落0—10cm表层土, 在每个群落当中的5个样方取样。群落生物量在70℃烘箱中烘干至衡重称量, 土壤全碳、全氮用Elementar (德国)进行分析, 土壤全磷用钼锑抗比色法测定。
收集连续3a生长季降雨及平均气温数据, 该数据由研究区工作站提供;从数据可以看出, 在观测的3个年份中, 2014年相对其他两年而言, 更加干旱、高温(图 1)。

|
| 图 1 2013—2015年生长季(6—8月)月降水量和月平均气温 Fig. 1 The three years (2013—2015) monthly precipitation and monthly average temperature in June to August |
将小样方中物种进行汇总, 构建物种(1表示物种出现, 0表示物种未出现)-样方二元矩阵, 在本研究中, 将总出现次数小于10次的物种进行了删除[12]。运用Ecosim 7.72软件通过零模型(Null Model)对于物种——样方二元矩阵进行5000次蒙特卡洛随机模拟得出的C-simulate值(μ)[17], 后利用C-score和V-ratio 2个指标分别来分析群落中物种的分布格局, 在计算过程中, 考虑了算法(FE, 行固定的而列进行等概率随机转换;FF, 行和列都固定, 进行等概率随机转换;FP, 行固定而列进行非等概率随机转换)对结果的影响, 其中, C-score的计算考虑了3种算法, 而V-ratio的计算仅考虑了FE、FP 2种算法。
1.3.1 C-scoreC-score是物种棋盘分布(Checkerboard unit, CU)的一个指数, 最早由Stone和Roberts[18]提出, 现已广泛应用于植物群落的格局分析[11,19]。计算方法为:

|
(1) |
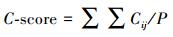
|
(2) |
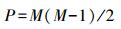
|
(3) |
其中, Ri为物种i出现的样方数, Rj为物种j出现的样方数, S为物种i和j同时出现的样方数, M为群落中的物种数。
1.3.2 V-ratioV-ratio(方差比率)最早由Schluter提出, 是用列的总和的方差与行的总和的方差之比来表示群落结构的一个指标[20], V-ratio不像C-score, 不是矩阵的行和列进行变换, 因而不存在FF算法。
1.3.3 标准效应量通过计算得出的C-score与软件模拟出的C-simulate值进行比较, 即可得出群落中物种的分布情况。为了便于结果的比较, 本研究进一步计算了C-score以及V-ratio矩阵的标准效应量(SES)[7]:
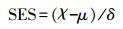
|
(4) |
式中, χ代表C-score或V-ratio的观测值(真实值), μ是运用Ecosim 7.72软件通过零模型(Null Model)对于物种-样方二元矩阵进行5000次蒙特卡洛随机模拟得出的C-score或V-ratio平均值, δ代表C-score或V-ratio的计算模拟平均值的标准差。
1.3.4 统计分析如果SES符合正态分布, 那么对于SES值的95%置信区间应为-2—2。对于C-score, 如果其SES值大于2则表明群落中物种非随机隔离, 即物种之间存在负关联, 说明竞争作用主导群落结构, 反之, 如果SES小于-2, 则表明群落中物种非随机聚集, 即物种之间存在正关联, 说明促进作用主导群落结构[7]。如果SES在-2与2之间, 则表明群落之间物种处于随机分布, 群落中物种之间相互关系不明显, 即中性作用主导群落结构。而对于V-ratio来说, 则恰恰相反, SES大于2时, 群落中物种非随机聚集, 即正相互作用主导群落结构, SES小于-2时, 群落中物种非随机隔离, 竞争作用主导群落结构[3]。运用SPSS 21.0对于SES, 群落数量特征以及土壤特征进行独立样本t检验。
2 结果与分析 2.1 利用方式对群落结构、物种组成及土壤性状的影响不同利用方式对于群落物种数及频度、群落高度、生物量以及土壤全氮的影响具有显著差异), 而对于群落的平均盖度、土壤全碳和全磷的影响则不显著。放牧群落在物种数上显著高于刈割群落, 而刈割群落在群落高度、生物量、和土壤全氮等指标上显著高于放牧群落(表 1, 表 2)。
| 利用方式 Utilization types |
物种数 Species |
群落高度 Height/ cm |
平均盖度 Cover/ % |
生物量 Biomass/ (g/m2) |
土壤全碳 Soil total C/ (mg/g) |
土壤全氮 Soil total N/ (mg/g) |
土壤全磷 Soil total P/ (mg/g) |
| 放牧Grazing | 17.32±0.76* | 45.5±8.32* | 44.75±4.39 | 71.26±4.94* | 12.83±0.28 | 1.52±0.03* | 0.46±0.03 |
| 刈割Mowing | 14.67±0.89* | 61.5±2.39* | 38.25±2.16 | 87.59±2.87* | 13.43±0.27 | 1.64±0.02* | 0.42±0.02 |
| *具有显著差异 | |||||||
| 植物名称 Species | 放牧 Grazing | 刈割 Mowing | |||||
| 2013 | 2014 | 2015 | 2013 | 2014 | 2015 | ||
| 大针茅 Stipa grandis | 144 | 82 | 104 | 158 | 68 | 137 | |
| 羊草 Leymus chinensis | 128 | 201 | 137 | 124 | 114 | 92 | |
| 麻花头 Serratula centauroide | 18 | 41 | 21 | 56 | 21 | 36 | |
| 冷蒿 Artemisia frigida | 55 | 29 | 43 | 70 | 35 | 53 | |
| 糙隐子草 Cleistogenes squarrosa | 182 | 133 | 201 | 147 | 155 | 129 | |
| 阿尔泰狗娃花 Heteropappus altaicus | 22 | 31 | 16 | 6 | 11 | 8 | |
| 二裂委陵菜 Potentilla bifurca | 4 | 17 | 7 | 28 | 13 | 22 | |
| 灰绿藜 Chenopodium glaucum | 33 | 26 | 14 | 12 | 20 | 31 | |
| 多根葱 Allium polyrhizum | 0 | 0 | 3 | 30 | 17 | 23 | |
| 达乌里芯芭 Cymbaria dahurica | 6 | 11 | 13 | 38 | 29 | 21 | |
| 猪毛菜 Salsolacollina Pall | 4 | 16 | 9 | 28 | 9 | 14 | |
| 洽草 Koeleria glauca | 11 | 14 | 18 | 28 | 11 | 19 | |
| 二色补血草 Limonium bicolor | 4 | 1 | 0 | 12 | 4 | 9 | |
| 瓣蕊唐松草 Thalictrum petaloideum | 21 | 15 | 7 | 4 | 13 | 11 | |
| 冰草 Agropyron cristatum | 126 | 109 | 146 | 76 | 132 | 116 | |
| 蓬子菜 Galium verum | 0 | 1 | 0 | 36 | 25 | 31 | |
| 防风 Saposhnikovia divaricata | 0 | 0 | 2 | 18 | 7 | 4 | |
| 披针叶黄华 Thermopsis lanceolata | 6 | 11 | 0 | 0 | 6 | 15 | |
| 黄花草木樨 Melilotus officinalis | 0 | 0 | 0 | 8 | 19 | 4 | |
在放牧利用方式下, C-score值的SES在FF算法下, 群落物种相互关系与随机模拟出的结果均无显著差异(P>0.05), 而C-score值的SES在FE和FP两种算法下, 其值均显著大于2(P<0.05)(表 3, 图 2), 图形位于双虚线区间上方, 即通过C-score值SES的模拟计算可得知在放牧利用方式下的群落物种之间相互关系由竞争作用所主导, 即群落总体呈现出竞争关系, 并且由于年际间气候的差异, 连续3年呈现出钟形曲线的趋势, 即观测中期(2014年), 群落物种竞争作用最强(图 2), 表明干旱改变了群落物种相互关系。而对于V-ratio值的SES, 与C-score值的SES结果相似, 群落总体呈现出竞争关系(图 3)。
| 群落类型 Community |
年份 Year |
零模型指数 Null index |
零模型指数 Null model |
真实值 Observed index |
计算机模拟值 Mean of simulated index |
平均标准效应量 Average SES |
P |
| 放牧 Grazing |
2013 | C-score | FE | 338.92 | 298.73 | 2.47 | 0.005 |
| FF | 338.92 | 340.93 | -0.57 | NS | |||
| FP | 338.92 | 259.95 | 4.45 | <0.001 | |||
| V-ratio | FE | 0.71 | 0.99 | -2.47 | 0.004 | ||
| FP | 0.71 | 1.28 | -4.26 | <0.001 | |||
| 2014 | C-score | FE | 433.83 | 349.37 | 4.76 | <0.001 | |
| FF | 433.83 | 421.91 | 2.56 | 0.009 | |||
| FP | 433.83 | 314.19 | 6.35 | <0.001 | |||
| V-ratio | FE | 0.52 | 1.00 | -4.07 | <0.001 | ||
| FP | 0.52 | 1.24 | -5.53 | <0.001 | |||
| 2015 | C-score | FE | 429.40 | 370.12 | 4.19 | <0.001 | |
| FF | 429.40 | 424.17 | 1.60 | NS | |||
| FP | 429.40 | 342.60 | 5.44 | <0.001 | |||
| V-ratio | FE | 0.54 | 0.99 | -3.79 | <0.001 | ||
| FP | 0.54 | 1.24 | -4.99 | <0.001 | |||
| 刈割 Mowing |
2013 | C-score | FE | 473.54 | 420.44 | 2.39 | 0.006 |
| FF | 473.54 | 478.28 | -1.13 | NS | |||
| FP | 473.54 | 370.17 | 4.23 | <0.001 | |||
| V-ratio | FE | 0.70 | 1.00 | -2.65 | 0.003 | ||
| FP | 0.70 | 1.27 | -4.16 | <0.001 | |||
| 2014 | C-score | FE | 471.28 | 489.31 | -3.23 | <0.001 | |
| FF | 471.28 | 467.53 | 0.99 | NS | |||
| FP | 471.28 | 345.34 | 5.53 | <0.001 | |||
| V-ratio | FE | 0.57 | 0.99 | -3.68 | <0.001 | ||
| FP | 0.57 | 1.25 | -5.09 | <0.001 | |||
| 2015 | C-score | FE | 400.73 | 352.88 | 3.43 | <0.001 | |
| FF | 400.73 | 395.91 | 1.76 | NS | |||
| FP | 400.73 | 315.04 | 5.40 | <0.001 | |||
| V-ratio | FE | 0.63 | 1.00 | -3.09 | <0.001 | ||
| FP | 0.63 | 1.32 | -4.92 | <0.001 | |||
| FE, 行固定的而列进行等概率随机转换Fixed-Equiprobable; FF, 行和列都固定, 进行等概率随机转换Fixed-Fixed; FP, 行固定, 而列按照比例进行非等概率随机转换Fixed-Proportional; NS表示不显著 | |||||||

|
| 图 2 不同利用方式下植物群落C-score的3种算法标准效应量的年际变化 Fig. 2 The interannual variance of three algorithms on standard effect size of communities C-score under different grassland utilization types FE:行是固定的, 而列进行等概率随机转换Fixed-Equiprobable; FF:行和列都固定, 进行等概率随机转换Fixed-Fixed; FP:行固定, 列按照比例进行非等概率转换Fixed-proportional; SES:Standard Effect Size |

|
| 图 3 不同利用方式下植物群落V-ratio的2种算法标准效应量的年际变化 Fig. 3 The interannual variance of two algorithms on standard effect size of communities V-ratio under different grassland utilization types FE:行是固定的, 而列进行等概率随机转换Fixed-equiprobable; FP:行固定, 列按照比例进行非等概率转换Fixed-proportional |
在刈割利用方式下, 对于C-score值的SES, 同样在FF算法下, 从2013年到2015年之间的群落物种相互关系与随机模拟结果无显著差异(P>0.05)。而C-score值的SES在FE和FP两种算法下, 从2013年到2015年有较为明显的变化, 两者趋势相同, 在2013年和2015年其值都落在了双虚线区间的上方, 表明群落物种之间相互关系由竞争所主导, 相反, 2014年的SES值则落在双虚线下方, 表明群落物种间相互作用关系发生明显改变, 由2013年较弱的竞争转变为促进作用, 群落总体格局发生明显变化, 而2015年又转变为较强的竞争(图 2)。对于刈割条件下的V-ratio(图 3), FE与FP两种算法对其SES得出的结果, 表明在2013至2015连续3a内群落物种总体相互作用关系均为竞争关系, 并且随着年际间气候变化, 竞争作用先增强后减弱。
3 讨论与结论 3.1 两种利用方式对于气候变化的不同响应1994年, Callaway和Bertness提出了著名的“胁迫梯度假说Stress Gradient Hypothesis, ”(SGH)[21], 即随着胁迫程度的增加, 物种之间的相互关系会由竞争转变为促进。在一些亚北极高山、苔原以及地中海生态系统的研究结果均支持该假说[22-25]。本研究发现, 两种利用方式对于群落物种相互关系的影响较小, 而气候变化对于群落物种相互关系的影响较大, 尤其在2014年最为明显。在放牧利用方式下, 不论是C-score还是V-ratio指数的标准效应量(SES)均表明该群落植物相互作用由竞争作用主导(FF算法例外), 并且在干旱高温的2014年竞争作用增强(图 2), 这一结果并不支持胁迫梯度假说;而在刈割利用方式下, 群落C-score与V-ratio的SES值则出现了两种相反的趋势(图 2, 图 3), 即C-score的SES值表明极端干旱的2014年竞争作用减弱, 主导群落相互作用关系转变为促进作用, 而其他两年则为竞争作用, 此结果支持胁迫梯度假说。SGH自提出之后, 就受到生态学研究者的广泛关注, 随着研究的逐渐深入, SGH的适应范围不断变化, Maestre在2009年对SGH进行了改进, 将传统的SGH线性模型精炼为单峰模型[11], 而Michalet, Fabio等人的研究结果则发现环境胁迫程度达到一定的强度时, 促进作用消失[26-29]。本研究在放牧利用下, 干旱年份竞争作用增强的可能原因是, 由于牛羊啃食, 减少建群种以及优势种的竞争优势地位, 从而使得群落当中的其他植物得以快速的生长, 并引起群落整体表现出竞争的结果[30,31], 即在相对高生产力的环境, 干扰能减轻高大优势植物对光、水分以及养分的竞争优势, 平衡了群落当中物种的相对地位[32]。
3.2 不同利用方式对于气候波动响应不同的内在机制很多学者认为非生物因素是改变群落格局的重要原因[4,7,33], 且有研究支持气候的年际波动对群落格局的影响大于利用方式, 如Kimberley对澳大利亚半干旱区灌木草原的研究发现, 干旱对于群落物种关系的影响要远大于放牧干扰[34], 与本研究所得结果类似, 2014年的气候波动对群落格局具有显著的影响。同时本研究还发现, 利用方式不同, 群落对气候波动的响应是不同的, 如2014年极端干旱的气候条件下, 放牧利用方式的响应为竞争加剧, 而刈割利用方式的响应为竞争减弱(促进作用), Fabio基于双晶格的模型推测、Bennett对于加拿大普列里草原的研究结果也有类似的报道[26,35-38]。在2014年, 群落建群种大针茅种群数量明显减少, 而克氏针茅、冷蒿和糙隐子草种群均增加调查数据, 未发表, 然而, 放牧群落和刈割群落对于气候干旱的响应发生了明显的不同, 植物群落种间关系表现出两种相反的结果, 支持温度与降水可能会改变群落物种相互关系的方向和强度[39]。刈割利用方式仅仅是在生长季结束时, 一次性对牧草进行收获。而在次年牧草返青直到下次刈割这一漫长的生长季当中, 整个群落几乎处于无干扰状态[35](除少部分小型动物, 如达乌里鼠兔Ochotona daurica、布氏田鼠Lasiopodomys brandtii等), 大针茅在群落当中的优势地位凸显, 而且刈割群落, 减少了放牧家畜对于植物以及土壤的践踏, 更有利于保持土壤原始的团粒结构, 并且能够增强土壤保水性[40]。Smit[41]的研究结果也发现, 在干扰(如放牧)存在的情况下, 物种间的促进作用消失, 表现出中性作用或竞争作用。而在生境相对温湿的钙质草原群落当中, 维管植物和苔藓植物也正是因为由于干扰的存在, 表现出较强的竞争作用[42]。
3.3 小尺度格局的优点及本研究的意义小尺度空间格局法与邻体移除法相比具有不破坏植物本身完整性, 将植物群落作为一个有机整体本身进行研究等优点[3,43-44], 减少了仅关注几个物种对的相互关系而缺少偶见种以及伴生种分析而造成的信息丢失。此外, 中期或长期的群落小尺度格局监测, 更能反映气候变化对于群落的影响[25]。本研究结果表明在极端干旱和高温气候年份, 放牧利用将加剧群落植物间的竞争而刈割利用方式下群落植物间的关系为促进, 支持已有的关于一年一次刈割方式较放牧更利于对大针茅草原的恢复[45];但本研究中2013和2015年并非极端降水和温度年份, 放牧和刈割两种利用方式对群落格局的影响基本一致, 所得结果不仅为SGH的完善提供了实验数据, 而且可为该区域草原的合理利用和保护措施的制定提供理论指导。
致谢: 内蒙古自治区锡林郭勒盟毛登牧场工作人员提供支持, 特此致谢。| [1] | Reitalu T, Prentice H C, Sykes M T, Lonn M, Johansson L J, Hall K. Plant species segregation on different spatial scales in semi-natural grasslands. Journal of Vegetation Science, 2008, 19(3): 407–416. DOI:10.3170/2008-8-18381 |
| [2] | Pottier J, Marrs R H, Bédécarrats A. Integrating ecological features of species in spatial pattern analysis of a plant community. Journal of Vegetation Science, 2007, 18(1): 223–230. |
| [3] | Badano E I, Cavieres L A, Molina-Montenegro M A, Quiroz C L. Slope aspect influences plant association patterns in the Mediterranean matorral of central Chile. Journal of Arid Environments, 2005, 62(1): 93–108. DOI:10.1016/j.jaridenv.2004.10.012 |
| [4] | Forrester D I. The spatial and temporal dynamics of species interactions in mixed-species forests: from pattern to process. Forest Ecology and Management, 2014, 312: 282–292. DOI:10.1016/j.foreco.2013.10.003 |
| [5] | Grubb P J. The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biological Reviews, 1977, 52(1): 107–145. DOI:10.1111/brv.1977.52.issue-1 |
| [6] | Pacala S W, Levin S A. Biologically generated spatial pattern and the coexistence of competing species//Tilman D, Kareiva P, eds. Spatial Ecology: the Role of Space in Population Dynamics and Interspecific Interactions. Princeton:Princeton University Press, 1997: 204–232. |
| [7] | López R P, Valdivia S, Rivera M L, Rios R S. Co-occurrence patterns along a regional aridity gradient of the subtropical Andes do not support stress gradient hypotheses. PLoS One, 2013, 8(3): e58518. DOI:10.1371/journal.pone.0058518 |
| [8] | Travis J M J, Brooker R W, Clark E J, Dytham C. The distribution of positive and negative species interactions across environmental gradients on a dual-lattice model. Journal of Theoretical Biology, 2006, 241(4): 896–902. DOI:10.1016/j.jtbi.2006.01.025 |
| [9] | Callaway R M, Walker L R. Competition and facilitation: a synthetic approach to interactions in plant communities. Ecology, 1997, 78(7): 1958–1965. DOI:10.1890/0012-9658(1997)078[1958:CAFASA]2.0.CO;2 |
| [10] | Tewksbury J J, Lloyd J D. Positive interactions under nurse-plants: spatial scale, stress gradients and benefactor size. Oecologia, 2001, 127(3): 425–434. DOI:10.1007/s004420000614 |
| [11] | Maestre F T, Callaway R M, Valladares F, Lortie C J. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. Journal of Ecology, 2009, 97(2): 199–205. DOI:10.1111/jec.2009.97.issue-2 |
| [12] | López R P, Squeo F A, Armas C, Kelt D A, Gutiérrez J R. Enhanced facilitation at the extreme end of the aridity gradient in the Atacama Desert: a community-level approach. Ecology, 2016, 97(6): 1593–1604. DOI:10.1890/15-1152.1 |
| [13] | Chen L P, Zhao N X, Zhang L H, Gao Y B. Responses of two dominant plant species to drought stress and defoliation in the Inner Mongolia steppe of China. Plant Ecology, 2013, 214(2): 221–229. DOI:10.1007/s11258-012-0161-y |
| [14] | 杨浩, 白永飞, 李永宏, 韩兴国. 内蒙古典型草原物种组成和群落结构对长期放牧的响应. 植物生态学报, 2009, 33(3): 499–507. |
| [15] | Pamela G, Martin R. Do species' strategies and type of stress predict net positive effects in an arid ecosystem?. Ecology, 2017, 98(3): 794–806. DOI:10.1002/ecy.1703 |
| [16] | Zhang J, Hao Z Q, Song B, Li B H, Wang X G, Ye J. Fine-scale species co-occurrence patterns in an old-growth temperate forest. Forest Ecology and Management, 2009, 257(10): 2115–2120. DOI:10.1016/j.foreco.2009.02.016 |
| [17] | Jiménez J J, Decaëns T, Rossi J P. Soil environmental heterogeneity allows spatial co-occurrence of competitor earthworm species in a gallery forest of the Colombian' Llanos'. Oikos, 2012, 121(6): 915–926. DOI:10.1111/more.2012.121.issue-6 |
| [18] | Stone L, Roberts A. The checkerboard score and species distributions. Oecologia, 1990, 85(1): 74–79. DOI:10.1007/BF00317345 |
| [19] | Luzuriaga A L, Sánchez A M, Maestre F T, Escudero A. Assemblage of a semi-arid annual plant community: abiotic and biotic filters act hierarchically. PLoS One, 2012, 7(7): e41270. DOI:10.1371/journal.pone.0041270 |
| [20] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998–1005. DOI:10.2307/1938071 |
| [21] | Bertness M D, Callaway R. Positive interactions in communities. Trends in Ecology & Evolution, 1994, 9(5): 191–193. |
| [22] | He Q, Bertness M D. Extreme stresses, niches, and positive species interactions along stress gradients. Ecology, 2014, 95(6): 1437–1443. DOI:10.1890/13-2226.1 |
| [23] | Reisner M D, Doescher P S, Pyke D A. Stress-gradient hypothesis explains susceptibility to Bromus tectorum invasion and community stability in North America's semi-arid Artemisia tridentata wyomingensis ecosystems. Journal of Vegetation Science, 2015, 26(6): 1212–1224. DOI:10.1111/jvs.2015.26.issue-6 |
| [24] | Brooker R W, Maestre F T, Callaway R M, Lortie C L, Cavieres L A, Kunstler G, Liancourt P, TielböRger K, Travis J M J, Anthelme F, Armas C, Coll L, Corcket E, Delzon S, Forey E, Kikvidze Z, Olofsson J, Pugnaire F, Quiroz C L, Saccone P, Schiffers K, Seifan M, Touzard B, Michalet R. Facilitation in plant communities: the past, the present, and the future. Journal of Ecology, 2008, 96(1): 18–34. |
| [25] | Schöb C, Michalet R, Cavieres L A, Pugnaire F I, Brooker R W, Butterfield B J, Cook B J, Kikvidze Z, Lortie C J, Xiao S, Al Hayek P, Anthelme F, Cranston B H, García M C, Le Bagousse-Pinguet Y, Reid A M, le Roux P C, Lingua E, Nyakatya M J, Touzard B, Zhao L, Callaway R M. A global analysis of bidirectional interactions in alpine plant communities shows facilitators experiencing strong reciprocal fitness costs. New Phytologist, 2014, 202(1): 95–105. DOI:10.1111/nph.12641 |
| [26] | Bulleri F, Xiao S, Maggi E, Benedetti-Cecchi L. Intensity and temporal variability as components of stress gradients: implications for the balance between competition and facilitation. Oikos, 2014, 123(1): 47–55. DOI:10.1111/more.2014.123.issue-1 |
| [27] | Michalet R, Schöb C, Lortie C J, Brooker R W, Callaway R M, Bailey J K. Partitioning net interactions among plants along altitudinal gradients to study community responses to climate change. Functional Ecology, 2014, 28(1): 75–86. DOI:10.1111/fec.2014.28.issue-1 |
| [28] | Michalet R, Brooker R W, Cavieres L A, Kikvidze Z, Lortie C J, Pugnaire F I, Valiente-Banuet A, Callaway R M. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities?. Ecolgy Letttters, 2006, 9(7): 767–773. DOI:10.1111/ele.2006.9.issue-7 |
| [29] | Michalet R, Pugnaire F I. Facilitation in communities: underlying mechanisms, community and ecosystem implications. Functional Ecology, 2016, 30(1): 3–9. DOI:10.1111/1365-2435.12602 |
| [30] | Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302–1310. DOI:10.1126/science.199.4335.1302 |
| [31] | Bruno J F, Stachowicz J J, Bertness M D. Inclusion of facilitation into ecological theory. Trends in Ecology & Evolution, 2003, 18(3): 119–125. |
| [32] | Zeidler M, Duchoslav M, Banaš M, Lešková M. Impacts of introduced dwarf pine (Pinus mugo) on the diversity and composition of alpine vegetation. Community Ecology, 2012, 13(2): 213–220. DOI:10.1556/ComEc.13.2012.2.11 |
| [33] | Alados C L, Navarro T, Komac B, Pascual V, Martinez F, Cabezudo B, Pueyo Y. Do vegetation patch spatial patterns disrupt the spatial organization of plant species?. Ecological Complexity, 2009, 6(2): 197–207. DOI:10.1016/j.ecocom.2008.10.015 |
| [34] | Howard K S C, Eldridge D J, Soliveres S. Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic and Applied Ecology, 2012, 13(2): 159–168. DOI:10.1016/j.baae.2012.02.008 |
| [35] | Maalouf J P, Le Bagousse-Pinguet Y, Marchand L, Touzard B, Michalet R. The interplay of stress and mowing disturbance for the intensity and importance of plant interactions in dry calcareous grasslands. Annals of Botany, 2012, 110(4): 821–828. DOI:10.1093/aob/mcs152 |
| [36] | Le Bagousse-Pinguet Y, Maalouf J P, Touzard B, Michalet R. Importance, but not intensity of plant interactions relates to species diversity under the interplay of stress and disturbance. Oikos, 2014, 123(7): 777–785. DOI:10.1111/more.2014.123.issue-7 |
| [37] | Martorell C, Almanza-Celis C A I, Pérez-García E A, Sánchez-Ken J G. Co-existence in a species-rich grassland: competition, facilitation and niche structure over a soil depth gradient. Journal of Vegetation Science, 2015, 26(4): 674–685. DOI:10.1111/jvs.12283 |
| [38] | Bennett J A, Cahill J F Jr. Evaluating the relationship between competition and productivity within a native grassland. PLoS One, 2012, 7(8): e43703. DOI:10.1371/journal.pone.0043703 |
| [39] | Baldwin A H, Jensen K, Schönfeldt M. Warming increases plant biomass and reduces diversity across continents, latitudes, and species migration scenarios in experimental wetland communities. Global Change Biology, 2014, 20(3): 835–850. DOI:10.1111/gcb.2014.20.issue-3 |
| [40] | 张成霞, 南志标. 放牧对草地土壤理化特性影响的研究进展. 草业学报, 2010, 19(4): 204–211. DOI:10.11686/cyxb20100426 |
| [41] | Smit C, Vandenberghe C, den Ouden J, Müller-Schärer H. Nurse plants, tree saplings and grazing pressure: changes in facilitation along a biotic environmental gradient. Oecologia, 2007, 152(2): 265–273. DOI:10.1007/s00442-006-0650-6 |
| [42] | Jeschke M, Kiehl K. Effects of a dense moss layer on germination and establishment of vascular plants in newly created calcareous grasslands. Flora-Morphology, Distribution, Functional Ecology of Plants, 2008, 203(7): 557–566. DOI:10.1016/j.flora.2007.09.006 |
| [43] | Hacker S D, Gaines S D. Some implications of direct positive interactions for community species diversity. Ecology, 1997, 78(7): 1990–2003. DOI:10.1890/0012-9658(1997)078[1990:SIODPI]2.0.CO;2 |
| [44] | Chu C J, Maestre F T, Xiao S, Weiner J, Wang Y S, Duan Z H, Wang G. Balance between facilitation and resource competition determines biomass-density relationships in plant populations. Ecology Letters, 2008, 11(11): 1189–1197. |
| [45] | 张建丽, 张丽红, 陈丽萍, 李军鹏, 赵念席, 高玉葆. 不同管理方式对锡林郭勒大针茅典型草原退化群落的恢复作用. 中国草地学报, 2012, 34(6): 81–88. |