 2017, Vol. 37
2017, Vol. 37文章信息
- 冯蕾, 赵运林, 张美文, 王勇, 徐正刚, 彭姣, 王双业
- FENG Lei, ZHAO Yunlin, ZHANG Meiwen, WANG Yong, XU Zhenggang, PENG Jiao, WANG Shuangye.
- 洞庭湖洲滩及滨湖区鼠类分布格局及其多样性
- Distribution pattern and diversity of rodent communities at beach and lakeside areas in the Dongting Lake region
- 生态学报. 2017, 37(17): 5771-5779
- Acta Ecologica Sinica. 2017, 37(17): 5771-5779
- http://dx.doi.org/10.5846/stxb201606061082
-
文章历史
- 收稿日期: 2016-06-06
- 网络出版日期: 2017-04-24

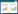




2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 洞庭湖湿地生态系统研究站, 长沙 410125;
3. 中南林业科技大学, 生命科学与技术学院, 长沙 410004
2. Dongting Lake Station for Wetland Ecosystem Research, Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. Central South University of Forestry and Technology, College of Life Science and Technology, Changsha 410004, China
鼠类是自然生态系统中食物链和能量流动的重要环节, 对维持生态平衡具有重要作用, 也与人类的生产、生活及身体健康等有着密切联系。一些鼠类为人类所利用[1];也有些种类能对人类经济和健康造成严重干扰及危害, 如害鼠种群的大暴发[2]。在洞庭湖区, 2007年东方田鼠(Microtus fortis)大暴发曾成为震惊全国的重大生态事件, 其成灾原因与人类社会经济活动密切相关[3]。随着人类社会经济活动对生态环境的影响, 曾经以东方田鼠为绝对优势种的洞庭湖洲滩生境, 其它鼠种已大量进入, 如黑线姬鼠(Apodemus agrarius)和褐家鼠(Rattus norvegicus), 可能的原因是三峡水库运行后, 洞庭湖洲滩受洪水淹没强度降低导致的生态反应[4]。因此, 为了有效预防和控制洞庭湖区害鼠类种群暴发, 有必要对洞庭湖洲滩以及毗邻的滨湖区的鼠类群落有所了解, 即掌握洞庭湖洲滩及其周围生境的鼠类分布格局。
当前涉及洞庭湖区鼠类分布格局已有一些研究, 陈安国等根据湖南全省的鼠类种群数量特征, 就全省鼠类害区类型进行了分析[5]。王勇等、张美文等对洞庭湖区域不同生态类型区的鼠类群落进行分析, 比较平原、丘陵和山区不同生态类型区的鼠类群落[6-7]。张美文等[8]对洞庭湖平原农房家鼠的栖息格局进行了研究。另外, 也有对洞庭湖滨湖区域单一生境, 如稻区[9]和丘岗地[10]鼠类群落结构的分析, 但上述研究均未将洞庭湖洲滩及其滨湖区域作为一个整体进行详细和具体分析。洞庭湖区域每年由于受湖区洲滩被水淹没的影响, 洲滩生境年内会发生较大改变, 进而与该区域鼠类群落有着密切相关。汛期, 随着湖水上涨, 湖滩面积缩小, 东方田鼠在拥挤的的压力下或直接被洪水所迫, 越过防洪堤迁入垸内。湖水回落时, 东方田鼠随湖滩出露而迁回沼泽草地。按迁移动因看, 此种迁移乃洪水逼迫所至, 而由逼迫外迁和自动回迁构成循环, 保证了种群对湖区特殊环境的适应[11]。因此洞庭湖东方田鼠的栖息生境具有季节性。东方田鼠高数量年份, 大量东方田鼠个体汛期受洪水影响从洲滩向滨湖农田迁移[11], 可对当地农作物造成重大为害。与之相反, 三峡水库运行后滨湖区域的黑线姬鼠和褐家鼠在向洲滩逐步渗入[4]。因此, 有必要将洲滩及其环湖滨湖区域作为一个整体进行分析, 为有效控制当地农业鼠害提供参考。
1 研究地区与方法 1.1 研究区概况洞庭湖位于中国湖南省北部, 长江中游荆江以南(27°39′—29°51′N, 111°19′—113°34′E, 图 1)。属于典型亚热带季风气候, 四季分明, 雨量充沛, 年平均气温16—17℃, 年平均降雨量1200—1550mm。洞庭湖不仅汛期长, 而且水位变幅大, 如城陵矶站多年平均年水位变幅13.35m, 最小10.67m, 最高达17.16m[12]。洞庭湖区是全国重要的商品粮、棉基地及淡水渔业生产基地[13]。长期以来, 由于经济发展和人类对自然生态系统的干扰, 致使长江中上游、四水流域水土流失严重, 湖区泥沙大量淤积, 湖面面积急剧减少。洞庭湖分为东洞庭湖、西洞庭湖、南洞庭湖3个部分[14]。沿洞庭湖区包括9个县, 分别是岳阳县、华容县、南县、安乡县、汉寿县、益阳县、湘阴县、沅江市、汨罗市(图 1)。

|
| 图 1 研究区位置和调查点位置图 Fig. 1 Study area of the survey location |
本研究自2003年起连续10a, 沿洞庭湖区的15个地点进行了鼠类群落调查(图 1), 调查点的设置基本覆盖整个洞庭湖区域, 东洞庭湖、西洞庭湖和南洞庭湖均有调查点分布。研究将调查区域大体可分为5种生境类型, 分别为成熟洲滩、退田还湖洲滩、滨湖农田、滨湖丘岗林地及滨湖农房生境, 每个调查点至少包含1种生境类型。除成熟洲滩外, 四季均有调查, 通常在1月(冬季)、4月(春季)、7月(夏季)、10月(秋季)进行。成熟洲滩汛期会被水淹没(大约是5—9月), 枯水期露出水面(大约是10月—来年4月), 因此夏季的7月一般无法在洲滩进行调查。且淹没时间长达数月, 尽管洲滩高程不一, 但汛期被水淹没后, 适宜鼠类生存的空间会很少。退水后的成熟洲滩主要覆盖有大量的薹草属(Carex Spp.)植物和辣蓼(Polygonum hydropiper)等植物, 或已开发为芦苇场, 种植有大面积的荻(Triarrhena lutarioriparia)与芦苇(Phragmites australis)等。退田还湖洲滩原为农业生产用地, 1998年实施退田还湖后变为洲滩, 种植有大量杨树, 汛期也会被淹没。滨湖农田生境主要种植水稻(夏季)和油菜(冬季), 堤外有大面积的洲滩区域, 汛期会有东方田鼠迁入。滨湖丘岗林地生境植被结构比较复杂, 乔木和灌木类植物占优势。草本类的植物以禾本科的植物较为常见, 如三毛草(Trisetum bifidum)和白茅(Imperuta cylindrica)。汛期常有东方田鼠种群迁入栖息。另外, 在安乡和汉寿的农房也做了少量调查, 由于总体布夹较少, 结果仅供参考。所有调查均采用夹日法, 以生葵花籽为饵, 沿直线每隔5m放置一个大号钢板夹(江西贵溪捕鼠器械厂), 每条调查样线每次至少布夹100个左右, 每次至少调查3条样线。农房每室1—2夹, 置于厨房、卧室、堂屋、畜圈及杂屋等处。每次调查在前1天17:00—19:00布夹, 第2天早上收回, 并记录数据。所捕获的鼠类分类统计, 进行群落结构分析, 并结合此区域历史积累资料进行讨论。
1.3 计算公式研究计算捕获率、优势度、群落多样性Shannon-Wiener指数等指标。
(1) 捕获率=捕获某鼠数目/有效置夹总数×100%
(2) 优势度I= Ni /N;优势集中性指数[15]C=∑(Ni/N)2, 式中Ni为每个种的个体数, N为总个体数。
(3) 群落多样性Shannon-Wiener指数[15]
(4) 相似性指数, 采用两个计算公式, 分别如下:
S1=2c/(a+b), 即Sorenson指数[16], 式中a、b为两群落各自的物种数, c为两群落共有的种数。
S2=1-0.5(∑|ai-bi|), 即Whittaker相似性指数[17], 式中ai为物种i的个体数在群落a中的比例, bi为物种i的个体数在群落物b中的比例。
以上参数计算均采用SPSS 18.0软件包进行。
2 结果与分析调查期间共布放有效夹日数79343只, 野外夹日数为78575只, 农房夹日数为768只。捕获鼠类3769只, 野外捕获3751只, 农房捕获18只。鼠类总捕获率为4.75%, 野外捕获率为4.77%, 农房捕获率为2.34%, 各鼠种在不同生境下的捕鼠数及捕获率见以下详细分析。
2.1 洞庭湖区不同生境鼠种组成现将2003—2013年在洞庭湖区不同生境中捕获的鼠类组成列于表 1中。
| 生境 Habitats | 捕获鼠数/只 No of capture | 种类组成Rodent composition/% | ||||||||
| 褐家鼠 R.norvegicus | 黄胸鼠 R.tanezumi | 小家鼠 M.musculus | 黑线姬鼠 A.agrarius | 东方田鼠 M.fortis | 黄毛鼠 R.losea | 社鼠 N.confucianus | 针毛鼠 N.fulvescens | 巢鼠 M.minutus | ||
| 成熟洲滩 Beach | 2551 | 2.74 | 1.65 | 0.00 | 27.01 | 68.01 | 0.12 | 0.00 | 0.08 | 0.27 |
| 退田还湖洲滩 Beach reconverted from farmland | 573 | 1.40 | 0.00 | 0.00 | 80.80 | 17.80 | 0.00 | 0.00 | 0.00 | 0.00 |
| 滨湖农田 Lakeside farmland | 498 | 4.62 | 0.40 | 0.60 | 44.18 | 49.60 | 0.40 | 0.00 | 0.00 | 0.20 |
| 滨湖丘岗林地 Hilly woodland | 129 | 3.10 | 0.00 | 0.00 | 15.50 | 34.88 | 3.10 | 20.93 | 21.71 | 0.78 |
| 滨湖农房 Residential premises | 18 | 0.00 | 66.67 | 33.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 大足鼠(R. nitidus)偶尔捕获, 因此不记录在表内 | ||||||||||
从表 1中可以看出, 在15个调查点5种生境下共捕获到10种鼠, 其中只有东方田鼠属于仓鼠科(Cricetidae), 其余9种都属于鼠科(Muridae)。黑线姬鼠、东方田鼠及褐家鼠3种鼠在除滨湖农房的其他4类生境下均能够捕到, 且黑线姬鼠在退田还湖洲滩生境所占比例达到80%以上, 东方田鼠在成熟洲滩生境所占比例达到60%以上, 2种鼠分别为2种生境下的绝对优势种。在滨湖农田和丘岗林地生境, 东方田鼠所占比例高于黑线姬鼠, 则可能是由于汛期东方田鼠大量迁入导致的。褐家鼠是家野两栖鼠, 可在住宅区及田野间自由迁移, 调查数据显示5种生境下, 褐家鼠在滨湖农田生境所占比例较高, 也主要由于滨湖农田生境下多有人类活动, 通常其数量分布距居民点越远就越少。其他鼠类中, 社鼠(Niviventer confucianus)和针毛鼠(Niviventer fulvescens)在滨湖丘岗林地生境所占比例相对较高, 其中社鼠只出现在滨湖岗地林地生境, 针毛鼠在成熟洲滩生境仅偶尔捕获。滨湖农房生境只捕获到黄胸鼠(Rattus tanezumi)和小家鼠(Mus musculus)。巢鼠(Micromys minutus)仅在滨湖农田、成熟洲滩及滨湖丘岗林地生境少量捕获, 所占比例较低, 而大足鼠(Rattus nitidus)只在成熟洲滩生境捕获到1次。
2.2 洞庭湖区不同生境鼠类捕获率为比较洞庭湖区不同生境各鼠类种群的密度, 列出了不同生境中鼠类捕获率数据见表 2。
| 生境 Habitats | 成熟洲滩 Beach | 退田还湖洲滩 Beach reconverted from farmland | 滨湖农田 >Lakeside farmland | 滨湖丘岗林地 Hilly woodland | 滨湖农房 Residential premises | |
| 夹日数No. of traps/只 | 26821 | 16076 | 31781 | 3897 | 768 | |
| 鼠数No. of number/只 | 2551 | 573 | 498 | 129 | 18 | |
| 总捕获率Trap succession/% | 9.51 | 3.56 | 1.57 | 3.31 | 2.34 | |
| 各鼠种捕获率 | 褐家鼠R. norvegicus | 0.26 | 0.05 | 0.07 | 0.10 | 0.00 |
| Trap succession of | 黄胸鼠R. tanezumi | 0.16 | 0.00 | 0.01 | 0.00 | 1.56 |
| each species/% | 小家鼠M. musculus | 0.00 | 0.00 | 0.01 | 0.00 | 0.78 |
| 黑线姬鼠A. agrarius | 2.57 | 2.88 | 0.69 | 0.51 | 0.00 | |
| 东方田鼠M. fortis | 6.47 | 0.63 | 0.78 | 1.15 | 0.00 | |
| 黄毛鼠R. losea | 0.01 | 0.00 | 0.01 | 0.10 | 0.00 | |
| 社鼠N.confucianus | 0.00 | 0.00 | 0.00 | 0.69 | 0.00 | |
| 针毛鼠N. fulvescens | 0.01 | 0.00 | 0.00 | 0.72 | 0.00 | |
| 巢鼠M. minutus | 0.03 | 0.00 | 0.00 | 0.03 | 0.00 | |
| 大足鼠(R. nitidus)只在成熟洲滩生境出现过1次, 因此不记录在表内 | ||||||
从表 2中可以看出, 5个调查生境的总捕获率从高到低依次为成熟洲滩、退田还湖洲滩、滨湖丘岗林地、滨湖农房、滨湖农田。成熟洲滩生境共捕获到8种鼠, 是捕获鼠种最多的生境, 捕获率最高为东方田鼠, 其次为黑线姬鼠。成熟洲滩作为东方田鼠的最适生境, 东方田鼠为绝对优势种。但与20世纪90年代[18-19]比较, 有明显变化, 即黑线姬鼠可在洲滩普遍捕获。其他几种鼠在成熟洲滩捕获率仍较低。在退田还湖洲滩生境(由农田被放弃耕作后演替而来)共捕获到3种鼠, 捕获率最高的为黑线姬鼠, 黑线姬鼠本身是生活在稻作区数量最多的野栖鼠, 退田还湖后在退田还湖洲滩捕获率仍较高;同时, 东方田鼠也在这一区域有较多栖息。滨湖农田生境共捕获到7种鼠, 捕获率最高的为东方田鼠, 其次为黑线姬鼠。农田是人类活动特别频繁的生境, 黑线姬鼠在当地农田的比例均较高, 在滨湖区域, 由于汛期有东方田鼠迁移, 改变了物种组成比例, 导致黑线姬鼠捕获率降低, 较高的东方田鼠捕获率具有临时特征;褐家鼠、黄胸鼠、小家鼠均是家野两栖鼠种, 能在农房与农田间来回迁移, 但在农田捕获率均较低。滨湖丘岗林地生境共捕获到7种鼠, 捕获率较高的有东方田鼠、针毛鼠和社鼠。东方田鼠主要是汛期洲滩迁入所致。针毛鼠和社鼠是长期栖息在山区和丘陵区林地鼠种, 因此在滨湖岗地林地生境这2种鼠的捕获率也较高。在滨湖农房生境共捕获到2种鼠, 为黄胸鼠和小家鼠。这2种鼠多栖息在农房区, 除此之外还有褐家鼠也喜欢栖息在农房区, 但由于布夹较少, 调查期间未捕获到褐家鼠。
2.3 洞庭湖区鼠类群落优势度及优势集中性指数现将各种鼠类群落优势度及优势集中性指数列出见表 3。
| 鼠种 Rodent species | 生境Habitats | |||||||||
| 滨湖农田 Lakeside farmland | 成熟洲滩 Beach | 退田还湖洲滩 Beach reconverted from farmland | 滨湖丘岗林地 Hilly woodland | 滨湖农房 Residential premises | ||||||
| 指标 Index | 序列 Sequence | 指标 Index | 序列 Sequence | 指标 Index | 序列 Sequence | 指标 Index | 序列 Sequence | 指标 Index | 序列 Sequence | |
| 褐家鼠R. norvegicus | 0.0462 | 3 | 0.0274 | 3 | 0.0140 | 3 | 0.0310 | 5 | ||
| 黄胸鼠R. tanezumi | 0.0040 | 6 | 0.0165 | 4 | 0.6667 | 1 | ||||
| 小家鼠M. musculus | 0.0060 | 4 | 0.3333 | 2 | ||||||
| 黑线姬鼠A. agrarius | 0.4418 | 2 | 0.2701 | 2 | 0.8080 | 1 | 0.1550 | 4 | ||
| 东方田鼠M. fortis | 0.4960 | 1 | 0.6801 | 1 | 0.1780 | 2 | 0.3488 | 1 | ||
| 黄毛鼠R. losea | 0.0040 | 5 | 0.0012 | 6 | 0.0310 | 6 | ||||
| 社鼠N.confucianus | 0.2093 | 3 | ||||||||
| 针毛鼠N. fulvescens | 0.0008 | 7 | 0.2171 | 2 | ||||||
| 巢鼠M. minutus | 0.0020 | 7 | 0.0027 | 5 | 0.0078 | 7 | ||||
| 大足鼠R. nitidus | 0.0004 | 8 | ||||||||
| C值 | 0.4434 | 4 | 0.5365 | 3 | 0.6847 | 1 | 0.2386 | 5 | 0.5556 | 2 |
| 指标表示优势度; C值为优势集中性指数 | ||||||||||
从表 3可以看出, 洞庭湖区鼠类群落在5种生境类型区的优势集中性差别相对较大。其中滨湖丘岗林地的优势集中性指数最低, 为0.2386, 退田还湖洲滩最高, 达到0.6847, 优势种为黑线姬鼠。成熟洲滩生境和滨湖农房生境的优势集中性指数较为相近, 分别为0.5365和0.5565, 成熟洲滩生境优势种为东方田鼠, 滨湖农房生境优势种为黄胸鼠。在滨湖丘岗林地生境, 各种类的优势度都不是很高, 最高的是东方田鼠种群, 为0.3488, 其次为针毛鼠和社鼠种群, 分别为0.2171和0.2093, 说明其优势种不突出, 因此群落优势集中性指数最低。成熟洲滩生境和滨湖农房生境, 虽然优势集中性指数接近, 但在优势种上却有差别, 成熟洲滩生境以东方田鼠为绝对优势种, 优势度为0.6801, 随后为黑线姬鼠0.2701, 这与以往相关报道相吻合[20-21], 即在成熟洲滩生境, 优势种都较为突出, 皆为东方田鼠。在滨湖农房生境, 以黄胸鼠为绝对优势种, 优势度达0.6667, 小家鼠为第二优势种, 优势度为0.3333, 也与之前的调查情况基本相符。在洞庭湖区, 黄胸鼠、小家鼠都以农房为最适栖息地。在滨湖农田生境中, 优势集中性指数为0.4434, 其中优势种为东方田鼠, 第二优势种为黑线姬鼠, 优势度分别为0.4960和0.4418, 优势度十分相近, 也可看出2种鼠竞争较为激烈, 由于东方田鼠的入侵, 对黑线姬鼠为优势的农田鼠类种群产生了较大影响。
2.4 洞庭湖区不同生境鼠类群落多样性不同生境鼠类群落多样性分析结果见表 4。从表 4可以看出, 滨湖丘岗林地生境物种多样性指数最高, 为1.5685, 这与一般情况下滨湖岗地林地植被覆盖度好, 受人类活动影响较少, 鼠类栖息条件好有关, 因此多样性指数高。其它生境要么受汛期洪涝淹没干扰, 要么受人类活动的干扰, 导致植被多样性低, 形成了优势种突出的群落, 农田和成熟洲滩生境的物种数量也不少, 但优势种突出, 因此与林地生境相比, 群落多样性也就有所降低。
| 评价指标 Evaluation index | 生境Habitats | 合计 Total | ||||
| 滨湖农田 Lakeside farmland | 成熟洲滩 Beach | 退田还湖洲滩 Beach reconverted from farmland | 滨湖丘岗林地 Hilly woodland | 滨湖农房 Residential premises | ||
| 物种数Numbers of species | 7 | 8 | 3 | 7 | 2 | 10 |
| 密度Trap success/% | 1.57 | 9.51 | 3.56 | 3.31 | 2.34 | 4.75 |
| 多样性指数H′Shannon-Weiner | 0.9380 | 0.8149 | 0.5392 | 1.5685 | 0.6365 | 0.8994 |
为了比较不同生境物种分隔的程度, 以了解各生境鼠类群落整体多样性, 通过比较不同生境鼠类群落的相似性指数, 对各个鼠类群落多样性差异进行定量分析, 进一步揭示鼠类群落的反应规律, 分析结果见表 5。
| Sorenson指数 Sorenson similarity index | 生境 Habitats | 1滨湖农田 Lakeside farmland | 2成熟洲滩 Beach | 3退田还湖洲滩 Beach reconverted from farmland | 4滨湖丘岗林地 Hilly woodland | 5滨湖农房 Residential premises | Whittaker相似性指数 Whittaker similarity index |
| S1 | 1滨湖农田 | 0.8013 | 0.6338 | 0.5408 | 0.0100 | S2 | |
| 2成熟洲滩 | 0.8000 | 0.4627 | 0.5365 | 0.0171 | |||
| 3退田还湖洲滩 | 0.6000 | 0.5455 | 0.3470 | 0.0000 | |||
| 4滨湖丘岗林地 | 0.7143 | 0.8000 | 0.6000 | 0.0000 | |||
| 5滨湖农房 | 0.4444 | 0.2000 | 0.0000 | 0.0000 |
表 5中, S1是以物种数为基础计算相似性指数, S2则将各鼠种比例也考虑到其中。总体来看, 相似性指数S1为0.0000—0.8000, S2的波动范围为0.0000—0.8013, 没有完全相似的两生境群落, 但存在完全的不相似的两生境群落的情况。
从相似性指数S1看, 生境2(成熟洲滩)分别和生境1(滨湖农田)及生境4(滨湖丘岗林地)两类生境的相似性指数最高, 达0.8000。而生境5(滨湖农房)分别和生境3(退田还湖洲滩)及生境4(滨湖丘岗林地)两类生境的相似性指数最低, 为0.0000。从之前分析已经可以看出, 由于退田还湖工程的实施, 使生境3中的2种家鼠已从该生境退出(即使有栖息, 也因种群数量特低而未捕获), 同时, 生境4物种多样性指数最高, 人类活动影响较少, 导致2种家鼠捕获率极低(或未捕获), 在这两方面的作用下, 导致了这两类生境与生境5的相似性最低。生境2(成熟洲滩)分别与生境1(滨湖农田)和生境4(滨湖丘岗林地)的相似性最高, 也是由于洲滩为东方田鼠栖息地, 而汛期大量东方田鼠迁入农田, 导致其具有较高的相似性。而在生境4中, 东方田鼠已经是该生境群落的优势种之一, 且为第一优势种, 因此也是其具有较高相似性的原因之一。
从相似性指数S2看, 与S1结果基本一致, 生境5(滨湖农房)分别和生境3(退田还湖洲滩)及生境4(滨湖丘岗林地)两类生境的相似性指数最低, 为0.0000, 生境2(成熟洲滩)与生境1(滨湖农田)的相似性指数最高, 为0.8013。而生境2和生境4的相似性指数与之前S1的结果相比有所下降, 主要原因可能是由于生境4的多样性指数较高, 物种数较多, 而在生境2中东方田鼠为绝对优势种所导致的结果。
3 讨论洞庭湖区域栖息的主要鼠种已有较多研究, 如黑线姬鼠[22]、东方田鼠[22]、褐家鼠[22]、黄胸鼠[23]、小家鼠[24]、社鼠[25]等的一些基本生物学特性和生态学特性均有一些报道。从群落来看, 不同生境的群落结构和演替也有一些报道, 如湖滩生境[4]、农田稻作区[9]、农房[8, 26]和丘岗林地[10]生境等, 还对平原、丘陵和山地生境的鼠类生物群落进行过较详细的比较研究[6-7]。但缺少将洞庭湖洲滩及其比邻的滨湖区域作为一个整体研究, 本文统计了这类区域调查的历史数据。
从种群密度、鼠种组成比例以及各项多样性指标来看, 东方田鼠和黑线姬鼠是洞庭湖洲滩和滨湖区域重要组分, 除农房外, 其它生境均有栖息。在整个洞庭湖区15个调查点5个生境中, 东方田鼠虽主要栖息于成熟洲滩生境, 但在除滨湖农房生境的其他3个生境均有捕获, 东方田鼠的栖息生境是有季节性的, 它们在洲滩与滨湖农田或岗地之间来回迁移[11]。说明滨湖区域是汛期洲滩被淹后东方田鼠的主要栖息生境。由于东方田鼠以洞庭湖洲滩为主要繁殖基地[22], 冬春季在湖滩大量繁殖, 汛期进入农田、岗地栖息, 具有明显的“避难”性质。洞庭湖区的东方田鼠种群就在洲滩与滨湖区域来回迁移以保障种群的顺利繁衍。在食物选择上, 东方田鼠适应能力强, 能依不同栖息地的植被结构调整摄食对象, 能适应湖区生活环境变化[27], 这也是其种群在洞庭湖区域能够繁殖发展的重要保障。
洞庭湖区黑线姬鼠栖息地广泛, 主要生活在农业区, 是长江流域稻作区数量最多的野栖鼠, 也是滨湖区域生境的主要优势鼠种。对洞庭湖区的黑线姬鼠种群已有较充分的研究, 已掌握洞庭湖区该鼠种的形态、栖息地、食物组成、繁殖特性、数量动态与预测模型, 以及为害特征、防治措施等[22, 28]。最新研究显示, 随着三峡工程对洞庭湖小兽群落影响效应的显现, 即洞庭湖洲滩汛期淹没强度的降低, 黑线姬鼠已大量侵入洲滩生境[4]。本研究调查结果也是对此现象的进一步印证, 即可以确认枯水季节黑线姬鼠大量迁入湖滩栖息已是即成事实, 且洲滩生境黑线姬鼠的密度已较高, 但从鼠种组成上看, 仍未超过东方田鼠。黑线姬鼠最高捕获率出现在退田还湖洲滩生境, 可能是退田还湖后任由其自由发展的结果。在退田还湖洲滩生境还捕到东方田鼠和褐家鼠, 由于退田还湖洲滩生境均不是这2种鼠的最适生境, 黑线姬鼠数量远远超过这2种鼠, 因此优势种突出。但随着退田还湖区域生境向成熟洲滩演替以及三峡水库调度对洞庭湖洲滩的影响, 此生境的黑线姬鼠和东方田鼠种群动态有可能发生相应的变化[4, 15]。
在洞庭湖区, 褐家鼠、小家鼠、黄胸鼠这3种鼠为家野两栖鼠, 通常栖息于滨湖农房生境中, 但野外也可捕到。滨湖农房常成为他们越冬和向农田扩散的基地[22-24]。本研究在滨湖农房生境只捕获到小家鼠及黄胸鼠, 并没有捕获到褐家鼠, 可能是其种群数量较低和布放夹日数较少的结果, 但它们在野外生境也均有捕获。在滨湖丘岗林地生境, 捕获率最高的为东方田鼠, 其次为社鼠和针毛鼠, 东方田鼠是汛期迁入的, 占用了滨湖丘岗林地的大量资源, 抑制了社鼠和针毛鼠的种群数量。一些研究发现, 社鼠在洞庭湖区主要栖息在山区林地, 此外仅局限于山边农田、农房偶尔捕获[29]。一般情况下, 对农业生产不会有很大危害, 而栖息于林地对维护生态平衡有一定益处, 如社鼠有利于一些植物种子的扩散与萌发、植被的演替等[30-33]。一般情况下社鼠在林地内密度不会太高, 但需要防范鼠传疾病, 社鼠是山区主要的疫源动物, 如在大别山岳西县境内的调查发现, 社鼠为肾综合症出血热主要宿主动物等[34]。针毛鼠主要栖息在林地, 是林业中的主要害鼠之一。与洞庭湖其他鼠相比, 针毛鼠栖息的林地, 生活环境受人类干扰相对较少, 林地的物种多样性较高, 种群的数量相对稳定。除以上几种害种之外, 仅在成熟洲滩、滨湖农田和丘岗林地生境内捕获到极少数的巢鼠, 巢鼠虽在洞庭湖平原及山地都有分布, 但数量极少, 虽对农业林危害不大, 但在某些疾病传播上有一定负面作用。
在不同生境, 每种鼠的习性与食物组成的差异也造成了它们选择不同的生境, 导致优势种不同。且每个生境条件下, 鼠类的物种多样性指数也存在不同。已有分析表明, 人类的干扰可降低鼠种丰富度和鼠类群落的多样性[35-38], 本文结果也与此一致。在洞庭湖滨湖区域除了人类活动的干扰外, 还有洪水对洲滩的影响, 因此在5类生境中, 干扰程度最低的丘岗林地鼠类群落多样性指最高, 其它受干扰较多生境的多样性均较低。在受干扰的生境种, 鼠类优势种突出, 进而为一些物种的种群爆发成灾创造了条件。特别值得注意的是湖滩生境由于受三峡工程的影响而导致每年的淹没强度减轻, 枯水期滨湖农田的一些种类大量进入湖滩[4], 物种数已达到与农田生境和丘岗林地生境相当, 但多样性指数仍较林地低, 优势集中度指数也较林地高, 说明其受洪涝的干扰, 仍具有优势种突出的特征。
| [1] | 陈安国. 第十二章啮齿动物的资源功能与保护//郑智民, 姜志宽, 陈安国. 啮齿动物学(第二版). 上海: 上海交通大学出版社, 2012: 619-713. |
| [2] | 夏武平. 害鼠与生态平衡//王祖望, 张知彬. 鼠害治理的理论与实践. 北京: 北京科学出版社, 1996: 2-18. |
| [3] | 张美文, 李波, 王勇. 洞庭湖区东方田鼠2007年暴发成灾的原因剖析. 农业现代化研究, 2007, 28(5): 601–605. |
| [4] | Zhang M W, Wang Y, Li B, Guo C, Huang G X, Shen G, Zhou X J. Small mammal community succession on the beach of Dongting Lake, China after the three gorges project. Integrative Zoology, 2014, 9(3): 294–308. DOI:10.1111/1749-4877.12073 |
| [5] | 陈安国, 袁主中, 张建云, 王勇, 郭聪, 程敬葆, 谭树林. 湖南农业鼠害防治技术研究Ⅰ.害鼠的种类、害区和与防治有关的生物学特性. 兽类学报, 1988, 8(3): 215–223. |
| [6] | 王勇, 张美文, 李波, 王凯荣. 洞庭湖地区不同生态类型区鼠类群落组成及其演替趋势. 农村生态环境, 2003, 19(1): 13–17. |
| [7] | 张美文, 王凯荣, 王勇, 李波. 洞庭湖区鼠类群落的物种多样性分析. 生态学报, 2003, 23(11): 2260–2270. DOI:10.3321/j.issn:1000-0933.2003.11.008 |
| [8] | 张美文, 王勇, 郭聪, 李波. 洞庭平原农房家鼠的栖息格局. 兽类学报, 2003, 23(2): 145–152. |
| [9] | 李波, 王勇, 张美文, 陈安国, 郭聪. 洞庭湖滨湖稻区鼠类群落基本特征研究. 中国生态农业学报, 2005, 13(1): 158–161. |
| [10] | 王勇, 赵尔宓, 郭聪, 李波, 张美文, 王广力. 洞庭湖滨湖丘岗地鼠类群落演替. 武汉大学学报:理学版, 2006, 52(2): 252–256. |
| [11] | 郭聪, 王勇, 陈安国, 李波, 张美文, 武正军. 洞庭湖区东方田鼠迁移的研究. 兽类学报, 1997, 17(4): 279–286. |
| [12] | 谢永宏, 张琛, 蒋勇. 洞庭湖湿地生态环境演变. 长沙: 湖南科学技术出版社, 2014. |
| [13] | 张光贵. 洞庭湖演变对农业生态环境的影响. 长江流域资源与环境, 1997, 6(4): 76–80. |
| [14] | 陆胤昊. 洞庭湖的演变及其驱动因子研究[D]. 武汉: 华中师范大学, 2009. |
| [15] | 张美文, 王勇, 李波, 郭聪, 谢永宏. 洞庭湖区退田还湖工程后小型兽类群落演替. 北京: 科学出版社, 2016. |
| [16] | Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213–251. DOI:10.2307/1218190 |
| [17] | Whittaker R H. Vegetation of the Siskiyou mountains, Oregon and California. Ecological Monographs, 1960, 30(3): 279–338. DOI:10.2307/1943563 |
| [18] | 王勇, 陈安国, 郭聪, 李波, 李世斌. 洞庭湖稻区黑线姬鼠种群数量预测. 兽类学报, 1997, 17(2): 125–130. |
| [19] | 李波, 陈安国, 郭聪, 王勇, 武正军, 张美文, 刘辉芬. 洞庭湖稻区鼠类群落及其种间关系初探//中国有害生物综合治理论文集. 长沙: 农业部全国农业技术推广服务中心, 中国昆虫学会, 1996: 11. |
| [20] | 王勇, 郭聪, 张美文, 李波, 陈安国. 洞庭湖区东方田鼠种群动态及其危害预警. 应用生态学报, 2004, 15(2): 308–312. |
| [21] | 张美文, 王勇, 李波, 黄国鲜, 郭聪. 三峡工程和退田还湖对洞庭湖区东方田鼠种群的潜在影响. 应用生态学报, 2012, 23(8): 2100–2106. |
| [22] | 陈安国, 郭聪, 王勇, 张美文, 刘辉芬, 李波. 第三章长江流域稻作区重要害鼠的生态学及控制对策//张知彬, 王祖望. 农业重要害鼠的生态学与控制对策. 北京: 海洋出版社, 1998: 114-174. |
| [23] | 张美文, 陈安国, 王勇, 郭聪, 刘辉芬, 李波. 长江流域黄胸鼠生物学特性观察. 兽类学报, 2000, 20(3): 200–211. |
| [24] | 郭聪, 陈安国, 王勇, 李世斌, 李波. 华中地区小家鼠生物学特性观察. 兽类学报, 1994, 14(1): 51–56. |
| [25] | 张美文, 黄璜, 王勇, 李波. 洞庭湖区社鼠的繁殖生态. 生态学报, 2006, 26(3): 884–894. |
| [26] | 王勇, 张美文, 李波. 洞庭湖区农村住宅鼠类群落结构和演替. 农业现代化研究, 2008, 19(6): 688–691. |
| [27] | 吴林, 张美文, 李波. 洞庭湖区东方田鼠的食物组成调查. 兽类学报, 1998, 18(4): 282–291. |
| [28] | 雍仲禹, 张美文, 郭聪, 王勇, 李波, 朱俊霞, 杨玉超, 徐正刚. 洞庭湖区黑线姬鼠食性调查. 动物学杂志, 2012, 47(3): 115–121. |
| [29] | 张美文, 黄璜, 王勇, 李波. 洞庭湖区社鼠(Niviventer confucianus)的外形特征及栖息地选择. 湖南农业大学学报, 2007, 33(1): 53–56. |
| [30] | 王巍, 马克平. 东灵山地区动物对辽东栎坚果的捕食和传播Ⅰ.排除啮齿目动物对坚果丢失的影响. 生态学报, 2001, 21(2): 204–210. |
| [31] | 张知彬, 王福生. 鼠类对山杏种子存活和萌发的影响. 生态学报, 2001, 21(11): 1761–1768. |
| [32] | 肖治术, 张知彬, 王玉山, 程瑾瑞. 都江堰地区灌丛内迺栎橡子的捕食和搬运. 动物学报, 2004, 50(4): 535–540. |
| [33] | Li H J, Zhang Z B. Effect of rodents on acorn dispersal and survival of the Liaodong oak (Quercus liaotungensis Koidz.). Forest Ecology and Management, 2003, 176(1/3): 387–396. |
| [34] | 柳燕, 吴松榆, 王俊, 张志平, 张福安, 王玉林, 刘红. 安徽省大别山区肾综合征出血热微小疫源地调查及分析. 安徽预防医学杂志, 2003, 9(5): 272–273, 276. |
| [35] | Stephenson P J. The small mammal fauna of Réserve Spécialed'Analamazaotra, Madagascar:the effects of human disturbance on endemic species diversity. Biodiversity and Conservation, 1993, 2(6): 603–615. DOI:10.1007/BF00051961 |
| [36] | Soul M E, Alberts A C, Bolger D T. The effects of habitat fragmentation on chaparral plants and vertebrates. Oikos, 1992, 63(1): 39–47. DOI:10.2307/3545514 |
| [37] | 吴德林, 罗成昌. 人类活动对云南哀牢山小型兽类群落结构的影响. 动物学研究, 1993, 14(1): 35–41. |
| [38] | 陈志平, 王应祥, 冯庆, 蒋学龙, 林苏. 云南西双版纳片断热带雨林鼠形啮齿类的物种多样性研究. 动物学研究, 1996, 17(4): 451–458. |