 2017, Vol. 37
2017, Vol. 37文章信息
- 金艳强, 李敬, 张一平, 刘运通, 费学海, 李培广
- JIN Yanqiang, LI Jing, ZHANG Yiping, LIU Yuntong, FEI Xuehai, LI Peiguang.
- 元江干热河谷稀树灌草丛植被碳储量及净初级生产力
- Carbon storage and net primary productivity of a savanna ecosystem in a dry-hot valley in Yuanjiang, Yunnan
- 生态学报. 2017, 37(17): 5584-5590
- Acta Ecologica Sinica. 2017, 37(17): 5584-5590
- http://dx.doi.org/10.5846/stxb201606011053
-
文章历史
- 收稿日期: 2016-06-01
- 网络出版日期: 2017-04-24





2. 中国科学院西双版纳热带植物园, 元江干热河谷生态站, 元江 653300;
3. 中国科学院大学, 北京 100049
2. Yuanjiang Savanna Ecosystem Research Station, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Yuanjiang 653300, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
森林固碳能力或碳储量是衡量森林生态功能的关键指标, 也是研究碳循环的基础。尤其是在全球变暖大背景下, 森林在调节全球碳平衡的主导地位显得尤为突出。然而, 近年来针对森林碳储量的研究主要集中于乔木林和灌木林, 对不同林型、不同起源林分的碳储量及生产力进行了大量研究[1-3]。但是, 稀树灌草丛(萨王纳, Savanna)作为一类特殊分布的森林类群, 其碳储量及生产力受到的关注较少。另外, 稀树灌草丛植被受气候变化的影响较为敏感。根据Zhao等[4]的模型研究认为受到气候变化的影响, 我国稀树灌草丛植被的面积有增大的趋势, 而且其分布的界限也正逐渐北移。因此, 研究稀树灌草丛碳储量及分配特征显得更为迫切。
分布于我国的稀树灌草丛植被中, 尤以元江干热河谷的最具代表性[5]。但是, 现有的研究主要针对元江稀树灌木草丛的群落特征[6-11]、植物生理[12-14]等方面展开, 而针对元江干热河谷稀树灌草丛的碳储量及生产力的研究较少。
元江干热河谷分布的稀树灌草丛作为世界萨王纳植被的重要组成部分, 其植被的碳储量及其分配特点是怎样的, 该植被类型的生产力和碳汇能力又是如何, 这些问题都急需回答。因此, 本研究选择元江干热河谷稀树灌草丛群落为对象, 旨在研究稀树灌木草丛群落碳储量及生产力特征, 为该区碳储量估算及碳汇功能评估提供基础, 同时可为该区植被的保护提供理论。
1 研究区概况研究地点位于中国科学院西双版纳热带植物园元江干热河谷生态站(102°10′40″E, 23°27′56″N), 海拔553 m, 该区植被以稀树灌木草丛为代表, 总体上植被覆盖较好, 乔木层一般较为低矮, 主要优势种为厚皮树(Lannea coromandelica)、细基丸(Polyalthia cerasoides);灌木多以宿萼木(Strophioblachia fimbricalyx)、虾子花(Woodfordia fruticosa)、清香木(Pistacia weinmanniifolia)、霸王鞭(Euphorbia royleana)等为优势, 灌木成丛现象比较明显, 平均高度80 cm左右, 盖度40%;草本以扭黄茅(Heteropogon contortus)为优势, 高度在10—60 cm之间, 盖度达50%以上。该区多年平均降雨量700—800 mm左右, 年平均温度24.9℃, 年日照时数2292 h, 年蒸发量2750 mm, 总体气候特点为降水量少, 平均温度高, 日照充足, 干湿季明显, 终年无霜[15]。生境中裸岩率高(达61.2%), 土层较薄, 土壤以燥红土为主。
2 研究方法 2.1 样方调查2014年10月通过踏勘, 并考虑林分的特点及代表性, 在研究区内林分特征及立地条件相对一致的地段选取了6块典型样地, 样地面积不低于600 m2 (20 m×30 m), 样地情况见表 1。将样地划分为100 m2 (10 m×10 m)的乔木样方, 分别对乔木(树高>3 m)进行每木检尺, 测定每株的树高、胸径、冠幅。在样地中随机的设置3个灌木样方, 大小为2 m×2 m, 布设于对角线上, 测定灌木样方的物种组成、灌木的平均基径、平均高度等。另外, 在3个灌木样方的左上角, 分别设置了1个草本样方(1 m × 1 m), 测定草本样方的总盖度、物种组成、平均高度等指标。最后, 采用收获法分别对灌木、草本样方中所有植物按器官(灌木分叶、枝干、根系三部分, 草本分地上、地下两部分)全部收获, 先将灌草地上部分齐地剪断收获;再用全挖法将灌草的根系全部挖出, 由于该区土层较薄, 挖掘深度直至岩层为止, 然后将灌木和草本各部分器官分别进行称鲜重, 同时每个部分取150 g左右的样品带回实验室, 在烘箱中用105℃杀青30min后, 恒温70℃下烘干至衡重后测定各部分器官的含水率, 根据灌草各部分器官的含水率, 将林下植被(灌木、草本)各部分的鲜重换算为干重生物量。最终, 将各部分器官的干重生物量统一乘以转换系数0.5得到各部分的碳储量(t C/hm2)。
| 样地 Site |
坡位 Slope position |
样地面积/m2 Stand area |
郁闭度 Coverage |
林分密度/ (株/hm2) Density |
平均树高/m Average height of trees |
| P1 | 中 | 1000 | 0.5 | 1560 | 4.4 |
| P2 | 中 | 1000 | 0.5 | 1460 | 4.0 |
| P3 | 中 | 600 | 0.45 | 1800 | 3.9 |
| P4 | 中 | 600 | 0.4 | 1766 | 3.8 |
| P5 | 中 | 600 | 0.45 | 1833 | 3.9 |
| P6 | 中 | 600 | 0.5 | 2100 | 3.9 |
乔木层生物量估算使用每木检尺的树高、胸径数据, 通过异速生长方程计算得到。考虑研究区域的气候、植被、树种组成及模型使用范围, 乔木层生物量估算选用的异速生长方程[16]:

|
(1) |
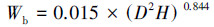
|
(2) |
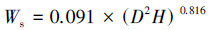
|
(3) |

|
(4) |
式中, Wl指叶生物量(kg), Wb指枝生物量(kg), Ws指茎干生物量(kg), Wr指根生物量(kg), D为乔木1.3 m处胸径(cm), H为树高(m)。将计算得到乔木、各部分生物量统一乘以转换系数0.5得到各部分的碳储量(t C/hm2)。
净初级生产力指的是生物体在单位面积、单位时间内有机物的净生产量。本研究中净初级生产力(Net primary productivity, NPP)使用林分中各组分碳储量的年变化量(t C hm-2 a-1)来表示, 其计算公式为:
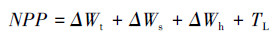
|
(5) |
式中, ΔWt、ΔWs、ΔWh分别指代乔木、灌木、草本碳储量年净增量, TL为凋落物年掉落量。乔木的增量依据2a的群落调查数据通过模型计算得到, 灌木、草本的增量依据2a通过全挖法的数据计算得到;凋落物量(TL)通过凋落物筐收集得到, 乘以转换系数0.5得到凋落物的碳储量。
3 结果与分析 3.1 稀树灌草丛乔木层碳储量稀树灌草丛群落乔木层的总碳储量为(26.68±0.88) t C/hm2 (图 1)。乔木层11种木本植物的碳储量在(0.05±0.03) —(16.47±2.01) t C/hm2之间, 物种间碳储量最大值比最小值高出339倍。碳储量最大的物种是厚皮树, 为16.47 t C/hm2, 占到乔木层总碳储量的61.71%, 其次为霸王鞭, 它的碳储量为5.58 t C/hm2, 占到20.90%;乔木层中碳储量最低的物种为疏序黄荆(Vitex negundo form. laxipaniculata), 为0.04 t hm-2, 仅占乔木层总碳储量的0.18%。这表明乔稀树灌草丛乔木层中优势物种比较突出, 碳储量在各物种间分布不均匀。

|
| 图 1 元江干热河谷稀树灌草丛乔木层碳储量(均值+标准误) Fig. 1 Carbon storage of tree layer for Savanna in Yuanjiang dry-hot valley (Values represent means ± SE) HC:心叶木Haldina cordifolia;PW:清香木Pistacia weinmanniifolia;SI:异序乌桕Sapium insigne;VA:深裂山葡萄Vitis amurensis var. dissecta;LC:厚皮树Lannea coromandelica;ER:霸王鞭Euphorbia royleana;PC:老人皮Polyalthia cerasoides;BB:鞍叶羊蹄甲Bauhinia brachycarpa;DY:云南柿Diospyros yunnanensis;PT:余甘子Phyllanthus taxodiifolius;VN:疏序黄荆Vitex negundo form. laxipaniculata |
另外, 乔木层碳储量中地上部分(叶、枝、茎)的总碳储量为(17.80±0.64) t C/hm2, 占总碳储量的66.70%, 地下部分碳储量为(8.89±0.26) t C/hm2, 占到33.30%。而且, 从地上部分碳储量上看, 枝和茎的碳储量(17.06 t C/hm2)就占到了95.79%, 但叶的生物量仅为0.54 t C/hm2, 只占到3.05%, 减少了叶片中的碳储量分配。
3.2 稀树灌草丛林下植被碳储量元江稀树灌草丛林下植被层总碳储量为(5.44±0.99) t C/hm2, 其中灌木层的总碳储量为(4.04±0.84) t C/hm2, 占林下植被层总碳储量的74.32%, 草本层碳储量为(1.40±0.28) t C/hm2, 占到25.70%(图 2)。

|
| 图 2 元江干热河谷稀树灌草丛林下植被碳储量(均值+标准误) Fig. 2 Carbon storage of understory layer for Savanna in Yuanjiang dry-hot valley (mean+SE) |
具体看来, 灌木层地上部分的叶、枝干碳储量0.29、2.43 t C/hm2, 分别占到灌木层碳储量的7.21%、60.12%。而灌木层的地下碳储量为(1.32±0.32) t C/hm2, 占灌木层碳储量的32.67%。草本层地上部分碳储量为(1.12±0.24) t C/hm2, 占草本层碳储量的79.89%;地下部分碳储量为(0.28±0.05) t C/hm2, 仅占草本层碳储量的20.04%。可知稀树灌草丛林下植被碳储量主要分配在地上部分。
3.3 稀树灌草丛林分净初级生产力元江稀树灌草丛净初级生产力为3.88 t C hm-2 a-1, 其中林分的净初级生产力为1.90 t C hm-2 a-1, 凋落物量为1.98 t C hm-2 a-1 (表 2)。从林分的净初级生产力看, 地上部分为1.33 t C hm-2 a-1, 占到了林分总净初级生产力的70.04%;而林分地下部分的净初级生产力是0.57 t C hm-2 a-1, 占到林分的29.96%。表明林分中地上净初级部分生产力更为明显, 当然, 29.96%比率也显示了地下部分的净初级生产力所占比重较大, 凸显了稀树灌草丛的特征。
| 层次Layer | 地上部分 Aboveground/(t C hm-2 a-1) |
地下部分 Belowground/(t C hm-2 a-1) |
合计 Totalt/(C hm-2 a-1) |
| 林分Forest | |||
| 乔木Tree | 0.62 | 0.39 | 1.01 (53.08) |
| 灌木Shrub | 0.51 | 0.05 | 0.56 (29.49) |
| 草本Herb | 0.20 | 0.13 | 0.33 (17.43) |
| 小计Total | 1.33 (70.04) | 0.57 (29.96) | 1.90 |
| 凋落物Litter | - | - | 1.98 |
| 总共Total | - | - | 3.88 |
| 括号内数字指代逻辑单元占林分总生产力的百分比 | |||
从林分的不同层次上看, 林分中乔木层净初级生产力为1.01 t C hm-2 a-1、灌木层0.56 t C hm-2 a-1、草本层0.33 t C hm-2 a-1, 分别占到林分总净初级生产力的53.08%、29.49%、17.43%。而林分中林下植被层(灌木层和草本层)的净初级生产力达到了46.92%, 接近于乔木层的净初级生产力, 说明在稀树灌草丛中林下植被层的净初级生产力对生态系统净初级生产力的贡献不可忽视。
4 讨论 4.1 稀树灌草丛碳储量森林生态系统作为一个动态的碳库, 准确估计森林的碳储量是正确评价森林碳汇功能的基础[17]。元江稀树灌草丛林分碳储量为32.13 t C/hm2, 在乔、灌、草3个层次中, 稀树灌草丛往地下部分分配了较多的碳储量(均占到总碳储量的30%以上), 反映了稀树灌草丛地下根系统较为发达。水分是限制稀树灌草丛生态系统生长的关键因子[15], 发达的根系有利于该植被中植物能从更深更广的区域获取水分, 这是对干旱胁迫的一种适应[18-19]。相反, 乔木层和灌木层中叶碳储量的分配较少, 其在总碳储量中的比例不足10%, 由于元江干热河谷水热矛盾突出, 较少的叶碳储量分配说明植物叶片的叶面积及叶数量的减少, 叶片也变薄[12], 这有利于减少水分的散失, 是适应干旱的一种策略[20]。
从同一地区来看, 位于哀牢山河谷的元江干热河谷稀树灌草丛的碳储量(32.13 t C/hm2)要远低于哀牢山山顶地区中山湿性常绿阔叶林的碳储量(257.90 t C/hm2)[30], 这主要是因为哀牢山山顶具有较好的水分条件(降雨量1879.5 mm)和土壤状况, 森林生长茂密, 树高可达20 m以上;而元江干热河谷稀树灌草丛分布于较为恶劣的石生环境中(裸岩率达到61.6%), 严重的限制了植被的生长, 植被一般较为低矮(乔木层平均树高4.0 m左右, 表 1), 制约了元江干热河谷稀树灌草丛植被的碳积累。
元江干热河谷稀树灌草丛植被的生长环境与喀斯特区灌木林生长环境极为相似, 都生长于石生环境中。比较两种生境中植被的碳储量发现, 元江稀树灌草丛的碳储量为32.13 t C/hm2, 稍高于贵州茂兰喀斯特区灌木林的碳储量(27.61 t C/hm2)[16], 但低于贵州普定喀斯特次生乔木林的碳储量(44.35 t C/hm2)[21]。这说明, 元江干热河谷稀树灌草丛生态系统碳储量与喀斯特区次生乔木林的碳储量相当。虽然, 在贵州喀斯特区平均年降雨量较为充足(1200 mm), 然而生境恶劣及土层浅薄也是限制喀斯特植被的生长的重要因素[16]。
与国内其他干热河谷区植被相比, 元江干热河谷稀树灌草丛的碳储量要高(表 3)。主要原因是在金沙江干热河谷水土流失严重, 土层中养分流失严重, 限制了植被的发育, 再加上该区多为人工林分, 人为干扰严重, 原生植被破坏严重, 影响植被的碳积累[11, 23-24]。相反, 元江干热河谷稀树灌草丛群落发育相对完善, 可以明显分出乔、灌、草三层[6, 11], 有利于植被的保护及碳积累。
| 地点 Location |
降雨量/ mm Precipitation |
林型 Forest type |
碳储量/(t C/hm2) Carbon storage |
文献 References |
|
| 喀斯特区Karst | 贵州普定 | 1390 | 常绿落叶阔叶混交林 | 44.35a | [21] |
| 贵州茂兰 | 1321 | 乔灌Trees and shrubs | 27.61 | [16] | |
| 干热河谷区 Dry-hot valley |
四川凉山 | 970 | 赤桉+合欢混交林 | 13.59a | [22] |
| 云南元谋 | 614 | Savanna桉树 | 24.73a | [23] | |
| 云南元阳 | 800—1000 | 膏桐人工林 | 11.91 | [24] | |
| 稀树灌草 丛Savanna |
巴西东南部Southeastern Brazil | 1550 | Cerrado savanna | 24.20 | [25] |
| 美国南部southern U. S. | 665 | temperate savanna | 19.0—43.0 | [26] | |
| 非洲南部Southern Africa | 850 | tropical savanna | 33.3 | [27] | |
| 澳洲北部Northern Australia | 1190—1695 | tropical savanna(桉树) | 31.75b | [28] | |
| 全球平均Global average | - | savanna | 28.40 | [29] | |
| 云南元江Yuanjiang, Yunnan | 700—800 | 稀树灌草丛savanna | 32.13 | 本研究 | |
| 碳储量一列中上标a表示该值由生物量乘以转换系数0.5得到;b表示该值为地上部分碳储量 | |||||
而与其它区域稀树灌草丛相比, 元江干热河谷稀树灌草丛碳储量(32.12 t C/hm2)要稍高于全球稀树灌草丛主要分布区的平均碳储量(28.40 t C/hm2)(表 3)和巴西东南部稀树灌草丛(Cerrado savanna)的24.20 t C/hm2, 而与美国南部(19.0—43.0 t C/hm2)、非洲南部(33.3 t C/hm2)、澳洲北部(31.75 t C/hm2)稀树灌草丛的碳储量比较接近。这表明该区稀树灌草丛植被的碳储量与世界其它稀树灌草丛分布区域的碳储量是相当的, 其碳储量不容忽视, 应得到重视与保护。
4.2 净初级生产力元江干热河谷稀树灌草丛总的净初级生产力为3.88 t C hm-2 a-1, 其中林分的净初级生产力为1.90 t C hm-2 a-1, 林分中林下植被的净初级生产力在林分净初级生产力的贡献达到了47%, 接近乔木层净初级生产力所占比例(53%)(表 2)。由于上层乔木通过控制林内小气候与资源利用率影响着林下植被的组成与生长[31-32], 元江稀树灌草丛乔木层较为开阔的林冠(Open-canopy, 郁闭度0.5左右), 为林下植被层获取光资源和空间生态位的占领提供了便利, 同时林下植物的多分枝(或分蘖)现象也会提高植物的生产力[33], 提高了植物的固碳能力。
本研究中稀树灌草丛总的净初级生产力要低于喀斯特森林的净初级生产力(4.34 t C hm-2 a-1)[34]。相比之下, 元江稀树灌草丛的净初级生产力是较低的, 然而, 元江干热河谷植被降水偏少, 土层浅薄等严酷环境会影响植物的发育, 间接影响到植被的固碳潜力[35]。与其它区域稀树灌草丛相比, 研究区稀树灌草丛的净初级生产力仅达到全球平均值(7.2 t C hm-2 a-1)[29]的一半左右。这主要是因为Grace等[29]在综合全球稀树灌草丛净初级生产力的数据中, 印度稀树灌草丛的净初级生产力普遍偏高(最高值达到22.8 t C hm-2 a-1), 拉高了全球的平均值, 而印度这些稀树灌草丛多为热带雨林退化形成, 其主要树种贡献了较高生产力。而与其它类型稀树灌草丛净初级生产力相比, 元江稀树灌草丛的净初级生产力处于要远低于澳洲北部桉树为主要优势种的稀树灌草丛(11.0 t C hm-2 a-1)[36], 由于桉树作为一种速生树种, 具有较高的生产力, 再加上澳洲北部较好的水分条件(平均年降雨量1200—1400 mm)促进了澳洲稀树灌草丛较高的生产力;其值要高于非洲西部(2.2 t C hm-2 a-1)[37]和美国南部(2.22 t C hm-2 a-1)[26]稀树灌草丛的净初级生产力, 更接近于南美洲稀树灌草丛的净初级生产力(3.9 t C hm-2 a-1)[38]。通过比较可以看出, 元江稀树灌草丛具有一定的净初级生产力, 其固碳潜力是不可忽视的。
5 结论(1) 元江稀树灌草丛植被的碳储量为32.13 t C/hm2。乔木层碳储量为26.70 t C/hm2, 主要树种间碳储量的分配不均匀。
(2) 林下植被层碳储量为5.44 t C/hm2, 其中灌木占74.3%, 草本层碳储量占25.7%。
(3) 元江稀树灌草丛植被总的净初级生产力为3.88 t C hm-2 a-1。林分中林下植被层的净初级生产力达到46.92%。林分的地上部分净初级生产力所占比重较大(70.04%), 但地下部分净初级生产力(29.96%)也较为明显。
(4) 元江干热河谷区稀树灌草丛植被的碳储量和固碳潜力不可忽视, 理应受到重视与保护。
致谢: 中国科学院西双版纳热带植物园元江干热河谷生态站和中心实验室大力协助, 刀万有、封乾元、刀新明帮助凋落物收集,特此致谢。| [1] | 胡会峰, 王志恒, 刘国华, 傅伯杰. 中国主要灌丛植被碳储量. 植物生态学报, 2006, 30(4): 539–544. |
| [2] | Zhang Y D, Gu F X, Liu S R, Liu Y C, Li C. Variations of carbon stock with forest types in subalpine region of southwestern China. Forest Ecology and Management, 2013, 300: 88–95. DOI:10.1016/j.foreco.2012.06.010 |
| [3] | Fang J Y, Chen A P, Peng C H, Zhao S Q, Ci L J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 2001, 292(5525): 2320–2322. DOI:10.1126/science.1058629 |
| [4] | Zhao D S, Wu S H. Responses of vegetation distribution to climate change in China. Theoretical and Applied Climatology, 2014, 117(1/2): 15–28. |
| [5] | 《云南植被》编写组. 云南植被. 北京: 科学出版社, 1987. |
| [6] | 金振洲. 云南元江干热河谷半萨王纳植被的植物群落学研究. 广西植物, 1999, 19(4): 289–302. |
| [7] | 刘方炎, 朱华. 元江干热河谷植被数量分类及其多样性分析. 广西植物, 2005, 25(1): 22–25. |
| [8] | 刘方炎, 朱华, 施济普, 陈晓鸣. 元江干热河谷植物群落特征及土壤肥力研究. 应用与环境生物学报, 2007, 13(6): 782–787. |
| [9] | 沈蕊, 张建利, 何彪, 李峰, 张志明, 周睿, 欧晓昆. 元江流域干热河谷草地植物群落结构特征与相似性分析. 生态环境学报, 2010, 19(12): 2821–2825. DOI:10.3969/j.issn.1674-5906.2010.12.009 |
| [10] | 朱华. 元江干热河谷肉质多刺灌丛的研究. 云南植物研究, 1990, 12(3): 301–310. |
| [11] | 金振洲, 欧晓昆. 干热河谷植被:元江、怒江、金沙江、澜沧江. 昆明: 云南大学出版社, 云南科技出版社, 2000. |
| [12] | 宋富强, 曹坤芳. 元江干热河谷植物叶片解剖和养分含量特征. 应用生态学报, 2005, 16(1): 33–38. |
| [13] | 张树斌, 张教林, 曹坤芳. 干旱对干热河谷优势木本植物虾子花光能分配的影响. 云南大学学报:自然科学版, 2014, 36(5): 774–780. |
| [14] | Zhang J L, Zhu J J, Cao K F. Seasonal variation in photosynthesis in six woody species with different leaf phenology in a valley savanna in southwestern China. Trees, 2007, 21(6): 631–643. DOI:10.1007/s00468-007-0156-9 |
| [15] | 张一平, 段泽新, 窦军霞. 岷江上游干暖河谷与元江干热河谷的气候特征比较研究. 长江流域资源与环境, 2005, 14(1): 76–82. |
| [16] | 黄宗胜, 喻理飞, 符裕红, 杨瑞. 茂兰退化喀斯特森林植被自然恢复中生态系统碳吸存特征. 植物生态学报, 2015, 39(6): 554–564. DOI:10.17521/cjpe.2015.0053 |
| [17] | 杨洪晓, 吴波, 张金屯, 林德荣, 常顺利. 森林生态系统的固碳功能和碳储量研究进展. 北京师范大学学报:自然科学版, 2005, 41(2): 172–177. |
| [18] | Santiago L S, Bonal D, De Guzman M E, Ávila-Lovera E. Drought survival strategies of tropical trees//Goldstein G, Santiago L S, eds. Tropical tree physiology:adaptations and responses in a changing environment. Cham:Springer International Publishing, 2016:243-258. |
| [19] | Brunner I, Herzog C, Dawes M A, Arend M, Sperisen C. How tree roots respond to drought. Frontiers in Plant Science, 2015, 6: 547. |
| [20] | Farooq M, Wahid A, Kobayashi N, Fujita D, Basra S M A. Plant drought stress:effects, mechanisms and management. Agronomy for Sustainable Development, 2009, 29(1): 185–212. DOI:10.1051/agro:2008021 |
| [21] | 刘长成, 魏雅芬, 刘玉国, 郭柯. 贵州普定喀斯特次生林乔灌层地上生物量. 植物生态学报, 2009, 33(4): 698–705. |
| [22] | 林伟宏, 陈克明, 刘照光. 川西南干热河谷赤按人工林生物量和营养元素含量. 山地研究, 1994, 12(4): 251–255. |
| [23] | 杨忠, 张建平, 王道杰, 张信宝. 元谋干热河谷桉树人工林生物量初步研究. 山地学报, 2001, 19(6): 503–510. |
| [24] | 郭永清. 云南干热河谷膏桐人工林生态系统碳库特征[D]. 南京: 南京林业大学, 2010. http://cdmd.cnki.com.cn/Article/CDMD-10298-2010253487.htm |
| [25] | Lilienfein J, Wilcke W. Element storage in native, agri-, and silvicultural ecosystems of the Brazilian savanna. Plant and Soil, 2003, 254(2): 425–442. DOI:10.1023/A:1025579932395 |
| [26] | Hughes R F, Archer S R, Asner G P, Wessman C A, McMurtry C, Nelson J, Ansley R J. Changes in aboveground primary production and carbon and nitrogen pools accompanying woody plant encroachment in a temperate savanna. Global Change Biology, 2006, 12(9): 1733–1747. DOI:10.1111/gcb.2006.12.issue-9 |
| [27] | Ryan C M, Williams M, Grace J. Above-and belowground carbon stocks in a miombo woodland landscape of Mozambique. Biotropica, 2011, 43(4): 423–432. DOI:10.1111/btp.2011.43.issue-4 |
| [28] | Scheiter S, Higgins S I, Beringer J, Hutley L B. Climate change and long-term fire management impacts on Australian savannas. New Phytologist, 2015, 205(3): 1211–1226. DOI:10.1111/nph.13130 |
| [29] | Grace J, José J S, Meir P, Miranda H S, Montes R A. Productivity and carbon fluxes of tropical savannas. Journal of Biogeography, 2006, 33(3): 387–400. DOI:10.1111/jbi.2006.33.issue-3 |
| [30] | 张鹏超, 张一平, 杨国平, 郑征, 刘玉洪, 谭正洪. 哀牢山亚热带常绿阔叶林乔木碳储量及固碳增量. 生态学杂志, 2010, 29(6): 1047–1053. |
| [31] | Augusto L, Dupouey J L, Ranger J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Annals of Forest Science, 2003, 60(8): 823–831. DOI:10.1051/forest:2003077 |
| [32] | Zapfack L, Engwald S, Sonke B, Achoundong G, Madong B A. The impact of land conversion on plant biodiversity in the forest zone of Cameroon. Biodiversity & Conservation, 2002, 11(11): 2047–2061. |
| [33] | Sah J P, Ross M S, Koptur S, Snyder J R. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. Forest Ecology and Management, 2004, 203(1/3): 319–329. |
| [34] | 于维莲, 董丹, 倪健. 中国西南山地喀斯特与非喀斯特森林的生物量与生产力比较. 亚热带资源与环境学报, 2010, 15(2): 25–30. |
| [35] | 田汉勤, 徐小锋, 宋霞. 干旱对陆地生态系统生产力的影响. 植物生态学报, 2007, 31(2): 231–241. DOI:10.17521/cjpe.2007.0027 |
| [36] | Chen X Y, Hutley L B, Eamus D. Carbon balance of a tropical savanna of northern Australia. Oecologia, 2003, 137(3): 405–416. DOI:10.1007/s00442-003-1358-5 |
| [37] | Hanan N P, Kabat P, Dolman A J, Elbers J A. Photosynthesis and carbon balance of a Sahelian fallow savanna. Global Change Biology, 1998, 4(5): 523–538. DOI:10.1046/j.1365-2486.1998.t01-1-00126.x |
| [38] | San José J J, Montes R A. Management effects on carbon stocks and fluxes across the Orinoco savannas. Forest Ecology and Management, 2001, 150(3): 293–311. DOI:10.1016/S0378-1127(00)00588-0 |