 2017, Vol. 37
2017, Vol. 37文章信息
- 蔡建堤, 徐春燕, 叶孙忠, 庄之栋, 马超, 刘勇, 陈梅芳, 沈长春
- CAI Jiandi, XU Chunyan, YE Sunzhong, ZHUANG Zhidong, MA Chao, LIU Yong, CHEN Meifang, SHEN Changchun.
- 闽东北海域假长缝拟对虾种群分布格局强度
- Pattern intensity of Parapenaeus fissuroides Crosnier in sea area in the northeast of Fujian
- 生态学报. 2017, 37(17): 5790-5797
- Acta Ecologica Sinica. 2017, 37(17): 5790-5797
- http://dx.doi.org/10.5846/stxb201605180959
-
文章历史
- 收稿日期: 2016-05-18
- 网络出版日期: 2017-04-24





2. 福建省海洋生物增养殖与高值化利用重点实验室, 厦门 361013;
3. 霞浦县海洋与渔业局, 霞浦 355100
2. Key Laboratory of Cultivation and High-value Utilization of Marine Organisms in Fujian Province, Xiamen 361013, China;
3. Xiapu Oceanic and Fishery Bureau, Xiapu 355100, China
种群分布格局强度是种群高密区和低密区在密度上的平均差异程度, 反映斑块和斑块间隙之间密度上的差异趋势, 用于测度一个种群在空间的聚集程度[1]。种群分布格局强度的研究对于确定种群特征、种群间相互关系以及种群与环境之间的关系具有非常重要的作用, 是群落空间结构的基本组成要素, 在测定分布格局的基础上进一步揭示格局强度十分必要[2-5]。集群是海洋游泳动物普遍的行为, 种群聚集形成的斑块形状、大小和间隙形式多样, 具有显著的空间异质性, 通过单位捕捞努力量渔获量(CPUE)可以直观地反映海洋游泳动物的时空分布格局[6-8], 但CPUE无法描述空间异质性、种群分布过程中没有涉及聚块间密度差的问题, 因而无法掌握种群的聚块大小差别、聚集强度、斑块组成形式及聚块内个体间的间隙等格局强度生态信息。
海洋作为复杂的生态系统, 海洋游泳生物形成斑块的形状、大小和种群间镶嵌结构等要素错综复杂, 表现出不同特征的格局强度。研究种群分布格局强度不仅仅是对种群聚集特性进行定量描述, 更重要的揭示种群聚集形成的原因, 从而提高渔情预报准确性和捕捞效率[9-13]。海洋生态学家已经对海洋游泳动物的种群分布格局强度进行了初步探索, 主要为应用格局强度指标研究海洋游泳动物聚集强度[14-15], 而对于不同格局强度指标的生态学意义、影响因子、适用性等, 尚缺系统深入研究。为此, 本文根据2008年5、8、11月和2009年2月, 在闽东北海域渔场开展的虾类专项调查研究资料, 以负二项参数、Mrisita指数、平均拥挤度、聚块指数和扩散系数为种群分布格局强度指标, 分析假长缝拟对虾(Parapenaeus fissuroides Crosnier)种群分布格局强度, 并探讨格局强度指标的生态学意义及其影响因子, 旨在探索出海洋游泳生物种群分布格局强度的研究方法, 挖掘假长缝拟对虾种群分布潜在的、有意义的规律, 为闽东北海域假长缝拟对虾渔业资源合理开发和利用探索新视角。
1 材料与方法 1.1 材料 1.1.1 研究对象假长缝拟对虾属于高温、高盐热带暖水性种类, 栖息于底质为沙砾、沙泥的海域。假长缝拟对虾是闽东北海域重要的经济虾类之一, 是拖虾作业重要的捕捞对象, 在东海和南海均有分布, 但主要分布在浙江中南部和福建北部外海60—120 m海域[16-20]。假长缝拟对虾的生命周期为1a, 生殖期为春末和夏季, 越冬期为秋季和冬季, 生殖期有较高的摄食量, 而越冬期减少摄食[17-18]。
1.1.2 数据来源2008年5、8、11月和2009年2月, 在闽东北海域渔场(26°00′N—28°00′N, 120°00′E—125°30′E)开展虾类的专项调查研究, 共设30个调查渔区(图 1)。调查船为闽霞渔1433桁杆拖虾船(船长36 m, 船宽6 m, 主机功率257 kW; 网具规格:网长32.6 m, 网口高2.7 m, 7个囊, 每囊袋长15 m(网目0.8 cm), 上口网目2.5 cm, 下口网目6 cm, 桁杆长32.4 m), 每航次在每个调查渔区拖曳1次, 拖曳时间均为1 h。调查按《海洋调查规范》(GB/T 12763.6—2007) 方法进行。不同月份各渔区假长缝拟对虾渔获量除以拖曳时间, 获得各月各渔区假长缝拟对虾的单位捕捞努力量渔获量(CPUE, Catch Per Unit Effort), 作为分析其种群分布格局强度的基础数据。

|
| 图 1 渔区图 Fig. 1 Map of fishing zone |
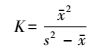
|
(1) |
式中, s2为各个渔区CPUE的方差, x为各个渔区CPUE的均值, K为负二项参数, 用来表示种群的聚集强度。K值愈小, 则种群聚集强度愈强。
1.2.2 Mrisita指数(I)[1-3]
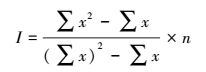
|
(2) |
式中, n为渔区数量, x为各个渔区CPUE, I值为Mrisita指数, 表示种群聚集强度。I值越大, 种群分布越集中。
1.2.3 Lloyd的平均拥挤度指数(m*)[22]
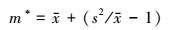
|
(3) |
式中, m*为平均拥挤度指数, 表示每个个体的平均拥挤程度。m*值越大, 则个体的平均拥挤程度越大。
1.2.4 聚块指数[1-3]
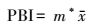
|
(4) |
式中, PBI为聚块指数, 表示斑块组成形式。PBI较大, 则代表主要种群数量集中分布于少数团聚的大斑块, 种群聚集度较高, PBI值较小, 则代表主要种群数量分散分布于多数斑块, 种群聚集度较低。
1.2.5 扩散系数(C)[1-3]
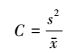
|
(5) |
式中, C为扩散系数, 表示种群扩散程度。C值越小, 种群扩散越明显。
2 结果与分析春季, C06渔区假长缝拟对虾的CPUE最大, 21.79%种群数量集中在该渔区, 种群聚集强度较弱, Mrisita指数较小, 负二项参数较大;夏季, C11渔区假长缝拟对虾的CPUE最大, 54.19%种群数量集中在该渔区, 种群聚集强度最强, Mrisita指数最大, 负二项参数最小;秋季, C03渔区假长缝拟对虾的CPUE最大, 28.10%种群数量集中在该渔区, 种群聚集强度较强, Mrisita指数较大, 负二项参数较小;冬季, C19渔区假长缝拟对虾的CPUE最大, 13.66%种群数量集中在该渔区, 种群聚集强度最弱, Mrisita指数最小, 负二项参数最大(表 1, 图 2)。表明负二项参数和Mrisita指数可用于表征种群聚集强度, Mrisita指数越大, 负二项参数越小, 种群聚集强度越强, 反之, 种群聚集强度越弱, 种群聚集强度夏季最强, 其余依次为秋季、春季和冬季(图 2)。
| 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | ||||
| 渔区Fishing zone | CPUE | 渔区Fishing zone | CPUE | 渔区Fishing zone | CPUE | 渔区Fishing zone | CPUE |
| C01 | C01 | C01 | C01 | 1157.6 | |||
| C02 | 439.5 | C02 | 9330.1 | C02 | 11027.8 | C02 | 2310.0 |
| C03 | 486.5 | C03 | 9465.2 | C03 | 14194.7 | C03 | 2431.2 |
| C04 | 984.5 | C04 | 59.1 | C04 | 1832.7 | C04 | 4496.5 |
| C05 | 265.0 | C05 | 3600.8 | C05 | 657.8 | C05 | 361.7 |
| C06 | 2979.7 | C06 | 707.1 | C06 | 2496.2 | C06 | 650.6 |
| C07 | 883.5 | C07 | 891.8 | C07 | 117.8 | C07 | 309.6 |
| C08 | 1025.5 | C08 | 551.9 | C08 | C08 | ||
| C09 | C09 | 107.0 | C09 | C09 | |||
| C10 | 593.0 | C10 | 517.0 | C10 | C10 | ||
| C11 | 1145.5 | C11 | 45872.5 | C11 | 10427.5 | C11 | 2910.2 |
| C12 | 805.5 | C12 | 686.3 | C12 | 4770.6 | C12 | 4962.1 |
| C13 | 97.2 | C13 | 1964.9 | C13 | 1385.8 | C13 | 1202.6 |
| C14 | 364.0 | C14 | C14 | 189.4 | C14 | 1028.3 | |
| C15 | 313.5 | C15 | 96.2 | C15 | 728.9 | C15 | 959.8 |
| C16 | 351.0 | C16 | 72.2 | C16 | 127.6 | C16 | 111.4 |
| C17 | C17 | 48.0 | C17 | C17 | |||
| C18 | 427.5 | C18 | 175.1 | C18 | C18 | 1287.2 | |
| C19 | 372.0 | C19 | 244.0 | C19 | 214.2 | C19 | 5248.0 |
| C20 | C20 | C20 | C20 | 1056.9 | |||
| C21 | 1268.5 | C21 | 4448.2 | C21 | 871.9 | C21 | 56.4 |
| C22 | 350.0 | C22 | 1824.2 | C22 | 951.4 | C22 | 3076.9 |
| C23 | 101.5 | C23 | C23 | 86.8 | C23 | 1016.7 | |
| C24 | 216.5 | C24 | C24 | 110.6 | C24 | 125.1 | |
| C25 | C25 | C25 | C25 | ||||
| C26 | C26 | 40.6 | C26 | C26 | |||
| C27 | 121.0 | C27 | 1756.9 | C27 | C27 | 2731.1 | |
| C28 | 70.0 | C28 | 1949.6 | C28 | 326.8 | C28 | 940.3 |
| C29 | C29 | 60.7 | C29 | C29 | |||
| C30 | 12.98 | C30 | 177.5 | C30 | C30 | ||
| 平均CPUE Average CPUE | 455.8 | 2821.6 | 1684.0 | 1281.1 | |||

|
| 图 2 不同季节假长缝拟的种群分布格局强度 Fig. 2 The population distribution pattern intensity Parapenaeus fissuroides in different seasons |
春季, 83.45%假长缝拟对虾种群数量分散于12个渔区(C02、C03、C04、C06、C07、C08、C10、C11、C12、C18、C19和C21), 即种群主要由较多数目的斑块组成, 种群分布比较分散, 聚块指数较小;12个渔区假长缝拟对虾平均CPUE最小, 为950.9 g/h, 个体平均拥挤的程度最小, 平均拥挤度指数最小, 个体间平均间距最大, 扩散系数最大(表 1, 图 2)。夏季, 81.65%假长缝拟对虾种群数量集中于4个渔区(C2、C3、C11和C21), 即种群主要由最小数目的斑块组成, 种群分布最为集中, 聚块指数最大;4个渔区假长缝拟对虾平均CPUE最大, 为17279.0 g/h, 个体平均拥挤的程度最大, 平均拥挤度指数最大, 个体间平均间距最小, 扩散系数最小(表 1, 图 2)。秋季, 84.95%假长缝拟对虾种群数量集中分布于5个渔区(C02、C03、C06、C11和C12), 即种群主要由较少数目的斑块组成, 种群分布较集中, 聚块指数较大;5个渔区假长缝拟对虾平均CPUE较大, 为8583.4 g/h, 个体平均拥挤的程度较大, 平均拥挤度指数较大, 个体间平均间距较小, 扩散系数较小(表 1, 图 2)。冬季, 90.85%假长缝拟对虾种群数量分散于14个渔区(C01、C02、C03、C04、C11、C12、C13、C14、C18、C19、C20、C22、C23和C27), 即种群主要由最多数目的斑块组成, 种群分布最为分散, 聚块指数最小;14个渔区平均假长缝拟对虾CPUE为2494.0 g/h, 个体平均拥挤的程度较小, 平均拥挤度指数较小, 个体间平均间距较大, 扩散系数较大(表 1, 图 2)。可见, 聚块指数体现了种群斑块的组成形式, 聚块指数大, 则大部分种群数量集中于少数斑块, 反之, 则大部分种群数量分散于多数斑块, 假长缝拟对虾种群聚块性夏季最强, 其余依次为秋季、春季和冬季(图 2);个体平均拥挤度指数反映平均个体实际的拥挤程度, 个体平均拥挤度指数大, 则大部分群体数量的平均个体间拥挤, 假长缝拟对虾个体平均拥挤度夏季最大, 其次是秋季、冬季, 春季最小(图 2);扩散系数反映了个体间平均间距, 个体平均间距大, 则种群扩散明显, 假长缝拟对虾种群扩散程度春季最大, 其次是冬季、秋季, 夏季最小(图 2)。
3 讨论海洋生物的聚集是一种较为复杂海洋生物行为反映的现象, 关于海洋生物聚集以及聚集不同性状, 不同大小的定义, 集群和集聚的命名等字义上的概念, 从20世纪20年代欧美开始有人研究, 并提出不同的定义和看法, 但至今尚没有一致的意见。以集群或者聚集等的单一概念已经难以表达不同性质的聚集特性, 而种群聚集强度、聚块性、平均拥挤度和扩散程度等种群分布格局强度指标能以不同角度在更精细地水平上反映一个种群在空间的不同性质聚集特性。
3.1 种群聚集强度种群聚集强度是度量一个种群分布格局的聚集程度, 是渔业资源学研究的重要内容, 是渔业资源利用和管理的基础, 单一团聚斑块种群的集中度在一定程度上反映种群聚集强度, 通过种群最为集中的斑块所占种群数量的比例数来反映种群聚集强度, 一般比例数越大, 种群就越集中, 种群聚集强度越强, 这种方法直观地反映了种群聚集状况, 但该方法只反映种群在单一斑块聚集度, 因此该方法用于度量种群聚集强度存在一定的片面性, 且不能用于比较不同的种群的聚集状况;而负二项参数从全局的角度反映种群聚集强度, 结果简单明了, 可用于比较同一种群在不同时间或不同生境中聚集强度的变化, 或者比较不同的种群在同时、同类生境中所呈现的聚集状况。负二项参数是通过种群高密区和低密区在密度上的平均差异程度来反映种群聚集强度, 种群分布越均匀, 方差就越小, 种群聚集强度越弱, 负二项参数越大(1) 式, 如冬季, 种群分布最均匀(表 1), 种群聚集强度最弱(图 2), 而方差越大, 种群聚集强度越强, 负二项参数越小(1) 式, 如夏季, 种群分布集中于4个渔区(表 1), 方差最大, 种群聚集强度最强(图 2)。
Mrisita指数计算公式是由渔区数量和各个渔区CPUE两个参数组成(2) 式, 负二项参数的计算公式是由方差和均值两个参数组成(1) 式, 显然两者计算公式的原理不同, 但两者分析种群聚集强度的结果一致, 两种种群聚集强度的计算方法之间既相互印证, 又相互补充, 提高种群分布格局强度适用于研究海洋游泳动物聚集强度结论的准确性和科学性。Mrisita指数不受平均CPUE的影响(2) 式, 种群聚集强度不因平均CPUE的改变而改变, 如平均CPUE春季小于冬季, 但种群聚集强度春季高于冬季。可见, 种群聚集强度是种群固有的特性, 平均CPUE不能准确反映种群聚集强度。
饵料浮游动物指使用浅水I型浮游生物网进行采样所获的浮游动物(剔除水母类和海搏类), 是经济海产动物(包括须鲸类、鱼类、虾类等)的饵料基础[23]。丰富的饵料浮游动物是假长缝拟对虾种群聚集的主要原因[20], 东海大陆架渔业资源调查资料显示[24-25], 闽东北海域, 夏季饵料浮游动物生物量为最高峰, 其次是秋季, 而春冬季生物量较低。饵料浮游动物生物量和负二项参数的关系为:
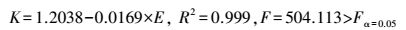
|
(6) |
式中, K为负二项参数, E为饵料浮游动物生物量(mg/m3)。R2为0.999, 说明饵料浮游动物生物量和负二项参数线性相关程度极高, 模型的拟合优度较好, 而F检验表明在Fα=0.05水平上模型回归效果显著。饵料浮游动物生物量越高, 负二项参数越小, 种群聚集度越强, 表明假长缝拟对虾具有集中索饵的特征, 饵料浮游动物是影响种群聚集强度的主要因子之一。
3.2 种群平均拥挤度平均拥挤度是指每个个体的平均邻居数, 可以通过种群主要数量的平均CPUE大小表达平均拥挤程度, 但在不同时间种群主要数量所占全体数量比例并不一致, 如夏季81.65%假长缝拟对虾种群数量(分布于4个渔区)为种群主要数量, 而秋季84.95%假长缝拟对虾种群数量(分布于5个渔区)为种群主要数量, 因此该方法用于度量种群平均拥挤度可能存在一定的偏颇。尽管平均CPUE可以作为度量周围种群个体拥挤度的一个相对指标, 但基于所有样方(包括个体数目为零的样方)的平均值而获得, 为样方的平均, 受CPUE为零的样方影响很大, 容易给出有偏差的信息, 如种群主要数量集中于少数样方时, CPUE为零的样方很多, 虽然平均数很小, 但实际上在种群内部的个体间很拥挤, 种内竞争激烈, 因此, 平均资源密度难以真正反映生物因素的影响效应, 有大量的CPUE为零的样方的情况下, 平均拥挤度的效果更好, 它可以比较真实地反映出种内竞争和生物因素的作用, 在格局分析中能提供更可靠的信息。各个季节假长缝拟对虾分布渔区数量较多, 出现频率大于60%, 受资源密度为零的渔区的影响较小, 故平均拥挤度和平均CPUE呈现出夏季>秋季>冬季>春季的一致趋势(表 1, 图 2)。
个体平均拥挤度高, 鱼群比较密集, 捕捞效率高, 鱼群具有捕捞价值。因此, 平均拥挤度在渔业资源开发和渔业管理具有较直接的指导意义。
3.3 种群聚块性聚块性是衡量种群聚集形成斑块的特征, 可以采用种群主要数量所占据的斑块数量表述种群聚块性, 但在不同时间种群主要数量所占全体数量比例往往不一致, 因此该方法只能定向描述种群聚块性, 用于定量分析种群聚块性存在一定的误差, 而聚块指数能真实体现种群斑块组成形式, 可用于比较同一种群或者不同种群斑块的组成形式。聚块指数考虑了空间格局本身的性质, 两个种群虽然平均CPUE不同, 可能显出同样的聚块性, 而两个种群虽然平均CPUE相同, 但是可能显出不同的聚块性(4) 式。由(1) 式、(3) 式和(4) 式可得PBI=1+1/K, 因此聚块指数也能度量种群聚集强度。
3.4 种群扩散程度扩散是生物个体之间相互远离的单线性运动, 是生物的基本特征之一, 对种群的分布、动态及遗传结构等方面均有重要影响[26-27]。扩散是研究种群数量变动的重要因子, 是生物寻找更广阔生活区域的一种方式, 扩散与聚集的强度不为反比关系, 即使聚集强度较强的种群可能存在扩散行为, 如春季种群聚集强度高于冬季, 而春季种群扩散程度大于冬季。说明扩散程度并非聚集强度简单的反义词, 种群扩散程度也是描述种群分布格局强度的指标之一。
扩散不仅有利于生物个体扩展其分布区, 寻找合适的生活环境, 还避免了种内竞争以及近缘个体之间的竞争, 有益于保持种群结构的稳定。种群竞争是影响种群分布格局强度最主要的生物学因素[26-29]。扩散系数和平均拥挤度(种内竞争)的关系为:

|
(7) |
式中, C为扩散系数, m*为平均拥挤度。R2为0.999, 说明扩散系数和平均拥挤度线性相关程度极高, 模型的拟合优度较好, 而F检验表明在Fα=0.05水平上模型回归效果显著。扩散系数越小, 种群扩散越明显, 则平均拥挤度越低, 种内竞争越小。种群扩散减少了种内竞争, 种内竞争是假长缝拟对虾种群扩散的主要因子之一。
林龙山[14]等采用平均值、丛生指数、扩散型指数、负二项参数和平均拥挤度5个格局强度指标反映台湾海峡夏季游泳动物优势种聚集强度;陈亚瞿等[15]采用丛生指数、聚块性指标、扩散指标、平均拥挤度和负二项参数5个格局强度指标反映南黄海、东海鲐鱼鲹鱼索铒场浮游动物聚集度。不同的种群分布格局强度指标具有不同的生态学意义, 其研究侧重点不同, 如负二项参数侧重于研究种群聚集强度、平均拥挤度侧重于表征个体平均拥挤程度、扩散系数主要用于体现种群扩散程度, 种群分布格局强度各指标的计算结果往往存在一定的偏差, 如在计算春季和冬季种群分布格局强度时, 其中负二项参数的值与平均拥挤度的值出现了不一致的结果, 平均拥挤度冬季大于春季, 而种群聚集强度春季大于冬季。可见, 以不同生态学意义的格局强度指标研究种群聚集强度有失偏颇, 甚至可能导致错误结果。
种群分布格局强度研究方法简化了集群行为研究的复杂性, 能细致地反映种群聚集强度、斑块的组成形式、个体平均拥挤度、种群内部的竞争程度和种群扩散程度等生态学信息, 且所需数据简单, 是一种帮助认识鱼群行为内在规律的有效手段。这些模型均是简化且突出重点的模型, 对模型的结果需要用实验观察测量来核实, 通过考察模型结果与实际观察资料比较, 用其相似程度来判断、推测和分析鱼群行为内在的机制, 筛选出具有重要影响的因子, 这种鱼类行为观察与数学模型相结合的方法, 有助于揭示鱼类和鱼群行为规律, 是现代鱼类行为科学研究的重要途径。
| [1] | 张金屯. 数量生态学. 北京: 科学出版社, 2011: 269–269. |
| [2] | 兰国玉, 雷瑞德. 植物种群空间分布格局研究方法概述. 西北林学院学报, 2003, 18(2): 17–21. |
| [3] | 杨持. 植物种群分布格局的强度与纹理. 内蒙古大学学报:自然科学版, 1988, 19(4): 662–668. |
| [4] | 刘金福, 洪伟. 格氏栲种群生态学研究Ⅶ.格氏栲种群分布格局的强度与纹理分析. 中南林学院学报, 1999, 19(1): 59–63. |
| [5] | 武玉珍, 张峰. 山西桑干河流域湿地植被优势种群分布格局研究. 植物研究, 2006, 26(6): 735–741. |
| [6] | 刘守海, 王金辉, 刘材材, 秦玉涛, 刘志国, 邓邦平. 长江口水域夏季鱼卵和仔稚鱼年间变化. 生态学报, 2015, 35(21): 7190–7197. |
| [7] | 杨胜龙, 马军杰, 张禹, 化成君, 戴阳. 大西洋中部延绳钓黄鳍金枪鱼渔场时空分布与温跃层的关系. 生态学报, 2013, 33(19): 6345–6353. |
| [8] | 陈云龙, 单秀娟, 周志鹏, 戴芳群, 李忠炉, 金显仕. 黄海细纹狮子鱼种群特征的年际变化. 生态学报, 2013, 33(19): 6227–6235. |
| [9] | 冯春雷, 李志国, 黄洪亮, 张勋, 陈雪忠. 鱼类行为研究在捕捞中的应用. 大连水产学院学报, 2009, 24(2): 167–170. |
| [10] | 周应祺, 王军, 钱卫国, 曹道梅, 张仲秋, 柳玲飞. 鱼类集群行为的研究进展. 上海海洋大学学报, 2013, 22(5): 734–743. |
| [11] | Breder C M. Studies of social grouping in fishes. Bulletin of the American Museum of Natural History, 1959, 117(6): 393–482. |
| [12] | Shaw E. Some news thoughts on the schooling of fishes. FAO Fisheries Reports, 1967, 62(2): 217–232. |
| [13] | 蔡建堤, 马超, 姜双城, 吴建绍, 蔡玉婷, 刘勇. 闽南-台湾浅滩渔场二长棘鲷群体集群行为宏观量化与分析. 水生生物学报, 2013, 37(2): 185–190. DOI:10.7541/2013.3 |
| [14] | 林龙山, 郑元甲, 马春艳. 台湾海峡夏秋季游泳动物资源分布及群落结构. 应用生态学报, 2005, 16(10): 1948–1951. DOI:10.3321/j.issn:1001-9332.2005.10.029 |
| [15] | 陈亚瞿, 徐兆礼. 南黄海、东海鲐鲹鱼索饵场浮游动物生态特征. 应用生态学报, 1990, 1(4): 327–332. |
| [16] | 刘瑞玉, 钟振如. 南海对虾类. 北京: 农业出版社, 1988: 84–88. |
| [17] | 宋海棠, 俞存根, 薛利建, 姚光展. 东海经济虾蟹类. 北京: 海洋出版社, 2006: 1–85. |
| [18] | 卢占晖, 薛利建, 张龙, 徐开达, 张亚洲. 东海大陆架虾类资源量评估. 水生生物学报, 2013, 37(5): 855–862. DOI:10.7541/2013.109 |
| [19] | 叶泉土, 黄培民, 叶孙忠. 闽东北外海假长缝拟对虾时空分布和生物学特性. 福建水产, 2006, 28(2): 7–11. |
| [20] | 王飞跃. 闽东北海域假长缝拟对虾的生物学特性及利用前景. 福建水产, 2014, 36(6): 285–293. |
| [21] | David F N, Moore P G. Notes on contagious distributions in plant populations. Annals of Botany, 1954, 18(1): 47–53. DOI:10.1093/oxfordjournals.aob.a083381 |
| [22] | Lloyd M. 'Mean crowding'. Journal of Animal Ecology, 1967, 36(1): 1–30. DOI:10.2307/3012 |
| [23] | 刘育莎. 福建三沙湾、兴化湾饵料浮游动物主要生态特征及次级产量的初步估算[D]. 厦门: 厦门大学, 2009. http://dspace.xmu.edu.cn/handle/2288/52843?show=full |
| [24] | 郑元甲, 陈雪忠, 程家骅, 王云龙, 沈新强, 陈卫忠, 李长松. 东海大陆架生物资源与环境. 上海:上海科学技术出版社, 2003: 169–175. |
| [25] | 徐兆礼, 晁敏, 陈亚瞿. 东海浮游动物生物量分布特征. 海洋学报, 2004, 26(3): 93–101. |
| [26] | 边疆晖. 根田鼠种群密度制约扩散的生态学过程及遗传效应[D]. 杭州: 浙江大学, 2005. |
| [27] | 张龙. 扩散种群的动力学模型研究[D]. 乌鲁木齐: 新疆大学, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10755-2007165529.htm |
| [28] | 毛凯, 李日华. 种群竞争模型的稳定性分析. 生物数学学报, 1999, 14(3): 288–292. |
| [29] | 陈玲玲, 林振山, 陈成忠, 梁仁君. 似Allee效应对2-物种集合种群竞争动态的影响. 生态学报, 2008, 28(2): 645–655. |