 2017, Vol. 37
2017, Vol. 37文章信息
- 宋贤冲, 郭丽梅, 田红灯, 邓小军, 赵连生, 曹继钊
- SONG Xianchong, GUO Limei, TIAN Hongdeng, DENG Xiaojun, ZHAO Liansheng, CAO Jizhao.
- 猫儿山不同海拔植被带土壤微生物群落功能多样性
- Variation of soil microbial community diversity along an elevational gradient in the Mao'er Mountain forest
- 生态学报. 2017, 37(16): 5428-5435
- Acta Ecologica Sinica. 2017, 37(16): 5428-5435
- http://dx.doi.org/10.5846/stxb201605311042
-
文章历史
- 收稿日期: 2016-05-31
- 网络出版日期: 2017-03-25





2. 中国林业科学研究院资源昆虫研究所, 昆明 650224;
3. 广西壮族自治区亚热带作物研究所, 南宁 530001;
4. 猫儿山国家级自然保护区管理局, 桂林 541001
2. The Research Institute of Resources Insect, Chinese Academy of Forestry, Kunming 650224, China;
3. Guangxi Zhuang Autonomous Region Subtropical Crops Research Institute, Nanning 530001, China;
4. Mao'er Mountain National Nature Reserve Administration, Guilin 541001, China
土壤微生物是评价土壤质量的重要指标之一, 其参与生态系统物质循环和能量流动, 在调控生物地球化学循环过程和维持生态系统功能方面起着关键作用[1-3]。近年来, 土壤微生物群落多样性已成为生态学领域的研究热点, 目前的研究主要集中在环境变化和人为干扰对土壤微生物多样性的影响, 如环境胁迫[4]、施肥方式[5]、植被类型改变[6]、作物连栽[7-8]。对微生物群落多样性进行描述的方法有很多, 但以群落水平碳源利用类型为基础的Biolog氧化还原技术, 因其较为简单、快速、灵敏度高、分辨率强, 而广泛用于土壤微生物群落的功能多样性研究[9]。
猫儿山国家级自然保护区位于我国广西壮族自治区桂林市北部, 在行政区划上地跨桂林市兴安、资源、龙胜3县。猫儿山国家级自然保护区位于中亚热带区域, 在中国植被区划上, 属于亚热带常绿阔叶林区、东部湿润阔叶林亚区、中亚热带常绿阔叶林南部亚地带的三江流域山地栲类木荷林, 石灰岩植被区和南岭山地栲类蕈树林区的过渡带, 拥有丰富的物种资源和典型的森林植被类型——亚热带常绿阔叶林[10]。该区域植被垂直带谱分布完整, 依次为常绿阔叶林、落叶阔叶混交林、针阔混交林、高山矮林和山顶灌草丛等[11]。近年来, 国内一些学者对该区域动植物[12-13]和大型蕈菌[14]多样性方面做了大量研究, 而对土壤微生物多样性的研究较少, 不同海拔梯度土壤的微生物群落结构和分布特性尚不明确。
海拔是一种重要的山地地形因子。随着海拔梯度的上升, 植被类型、气候等环境因子也发生改变, 进而引起土壤微生物群落多样性的变化[15]。不同区域土壤微生物群落多样性沿海拔梯度的变化规律有所不同。刘秉儒等[16]对处于干旱风沙区的贺兰山不同海拔的6个典型植被带土壤微生物多样性进行研究, 发现随着海拔的升高, 土壤微生物群落多样性指数呈增大的趋势。吴则焰等[17]研究了中亚热带武夷山不同海拔植被带的土壤微生物群落多样性差异, 结果表明随着海拔上升土壤微生物群落多样性逐渐下降。Shen等[18]研究了长白山细菌群落多样性的海拔梯度变化, 发现细菌群落多样性随着海拔升高并没有明显的变化规律。本研究采用Biolog微平板技术, 对猫儿山国家级自然保护区不同海拔植被带的土壤微生物群落结构和代谢功能多样性的变化规律进行研究, 旨在探讨不同海拔植被带土壤微生物群落功能的差异和变化规律。研究结果不仅可以为揭示中亚热带森林生态系统土壤微生物群落变化规律提供理论依据, 也为进一步探讨微生物群落功能与环境因子的相关性奠定基础。
1 研究地区与研究方法 1.1 研究区概况研究地点位于广西壮族自治区桂林市兴安县的猫儿山国家级自然保护区(25°48′—25°58′N, 110°20′—110°35′E), 主峰海拔2142 m。该地区属于中亚热带季风气候;年平均气温12.8℃, 平均月最低和最高气温分别为2.0℃和21.8℃;年均相对湿度92%, 多年年均降水量为2509.1 mm, 降水主要发生在5—6月[10]。猫儿山海拔相对高差大, 植被垂直植被带变化明显, 沿海拔植被带依次为常绿阔叶林、常绿落叶阔叶混交林、针叶阔叶混交林、高山矮林和山顶灌草丛等[11]。主要地貌类型为中山, 地带性土壤以山地红壤为主, 由花岗岩风化形成。
1.2 研究方法 1.2.1 土壤样品采集与处理2014年8月, 参照国家林业行业标准LY/T 1952—2011[19]分别选取常绿落叶林(evergreen broad-leaved forest, EBF)、落叶阔叶混交林(deciduous broad-leaved forest, DBF)和针阔混交林(coniferous and broad-leaved forest, CBF) 3种有代表性的典型植被带, 在每个植被带设置3个20 m×20 m样地, 样地基本情况见表 1。在每个样地内采用梅花5点法采集枯枝落叶层下0—10 cm土壤, 混合为1个土样, 装入封口袋并放入装有冰块的保温箱。新鲜土样带回实验室后分为2份, 1份过2 mm筛后放在4℃冰箱保存, 用于土壤微生物群落特征测定;另1份自然风干后过2 mm筛, 用于测定土壤理化性质。
| 植被带 Vegetation zones |
海拔/m Elevation |
地理坐标 Geographic coordinate |
坡度/(°) Slope |
土壤类型 Soil type |
林分起源 Stands origin |
代表植被 Conditions of vegetation |
| 常绿阔叶林 EBF |
1138 | 110°29′11″E 25°52′54″N |
18 | 山地黄壤 | 天然林 | 桂南木莲Manglietia chingii、银木荷Schima argentea、罗浮栲Castanopsis faberi、羊角杜鹃Rhododendron cavaleriei等 |
| 落叶阔叶 混交林DBF |
1562 | 110°27′57″E 25°54′37″N |
29 | 山地黄壤 | 天然林 | 多脉青冈Cyclobalanopsis multinervis、 红岩杜鹃Rhododendron haofui、 枫香Liquidambar formosana等 |
| 针阔混交林 CBF |
2042 | 110°26′01″E 25°53′48″N |
3 | 山地沼泽泥炭土 | 次生林 | 南方铁杉Tsuga chinensis、 曼椆Cyclobalanopsis oxyodon、 丁香杜鹃Rhododendron farrerae、 箭竹Fargesia spathacea等 |
| EBF:常绿阔叶林evergreen broad-leaved forest; DBF:落叶阔叶混交林deciduous broad-leaved forest; CBF:针阔混交林coniferous and broad-leaved forest | ||||||
土壤理化性质测定参照鲁如坤主编的《土壤农业化学分析方法》[20]和《中华人民共和国林业行业标准LY/T 1210-1275—1999》[21]。土壤pH值用水浸提电位法(水土比为2.5:1);土壤质量含水量测定采用烘干法;土壤总有机碳(TOC)测定采用重铬酸钾外加热法;全氮(TN)采用半微量凯式法;速效氮(AN)采用碱解扩散法;全磷(TP)和有效磷(AP)采用钼锑抗比色法;全钾(TK)和速效钾(AK)采用火焰光度法。
1.2.3 土壤微生物群落功能多样性的测定称取10 g新鲜土样加入含100 mL 0.85%无菌NaCl溶液的三角瓶中, 封口, 180 r/min, 28℃振荡30 min。静置2 min, 取上清液2 mL加入18 mL 0.85%无菌NaCl溶液中, 如此重复稀释1次, 制得1:1000的提取液。取150 μL该提取液接种到生态板的每一个孔中, 最后将接种好的板置于28℃的恒温培养箱中培养, 每隔24 h在Biolog读数仪读取590 nm的光密度值, 培养时间共为168 h。
1.3 数据处理土壤微生物群落利用碳源的能力, 用平均颜色变化率(Average well color development, AWCD)来表示, 其计算方法如下[22]:
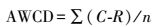
|
式中, C为每个有培养基孔的光密度值;R为对照孔的光密度值;n为碳源底物种类, n=31;C-R≤0的孔在计算中记为0[23]。土壤微生物群落功能多样性指数分析采用丰富度指数、Shannon指数和均匀度指数[24]。丰富度指数指被利用的碳源的总数目, 微孔的光密度值≥0.2, 则认为是阳性值并计入微生物群落的丰富度S[25]。
为了消除接种密度带来的差异, 本研究采用培养96 h的光密度值来进行主成分分析、相关分析和多样性指数分析。统计分析采用Excel和SPSS 17.0进行。
2 结果与分析 2.1 不同海拔植被带土壤特征随着海拔梯度的上升, 3种植被带土壤理化性质表现出不同的变化趋势(表 2)。土壤pH随海拔梯度的上升表现出递减的趋势, 而土壤含水量表现出逐渐增加的趋势。不同海拔植被带土壤化学性质差异显著, 其中TOC、TN、AN、TP、AP和AK随海拔梯度上升均呈现先下降后上升的趋势;TK表现出递减的趋势。TOC、TN、AN、AP、AK的变化规律相同, 即CBF>EBF>DBF。
| 植被带 Vegetation belts |
pH | 含水量 Water content/% |
总有机碳 TOC / (g/kg) |
全N TN / (g/kg) |
全P TP / (g/kg) |
全K TK / (g/kg) |
速效N AN / (mg/kg) |
速效P AP / (mg/kg) |
速效K AK / (mg/kg) |
| 常绿阔叶林EBF | 4.37±0.50a | 39.19±3.43b | 108.12±9.63b | 7.82±0.69b | 2.49±0.15a | 28.72±0.19a | 633.47±81.16b | 4.13±0.81b | 129.90±17.98b |
| 落叶阔叶混交林DBF | 3.84±0.12ab | 39.89±3.29b | 51.91±13.48c | 3.87±1.14c | 1.05±0.20c | 24.15±2.02b | 496.43±165.05b | 3.67±1.55b | 78.17±6.88c |
| 针阔混交林CBF | 3.57±0.06b | 77.22±2.24a | 392.74±11.43a | 20.34±0.42a | 2.03±0.08b | 5.26±0.10c | 2095.50±93.45a | 38.67±2.59a | 209.73±12.02a |
| 同一列数据中不同字母表示差异显著(P<0.05) | |||||||||
平均颜色变化率(AWCD)反映土壤微生物群落对单一碳源的利用能力, 是土壤微生物活性及群落功能多样性的重要指标[17]。由图 1可知, 不同植被带AWCD随培养时间的延长而增大, 微生物利用碳源底物的能力即活性随培养时间的延长而提高。前48 h内土壤微生物活性增加比较缓慢, 48 h后开始急剧增加, 在96—120 h内增长速率最高, 144 h后逐渐趋于缓慢变化。不同海拔植被带的土壤微生物利用单一碳源能力的顺序为:EBF>DBF>CBF。

|
| 图 1 不同植被带土壤微生物群落的平均颜色变化率 Fig. 1 Average well color development of soil microbe community in different vegetation belts 不同小写字母表示差异显著(P<0.05);EBF:常绿阔叶林evergreen broad-leaved forest;DBF:落叶阔叶混交林deciduous broad-leaved forest;CBF:针阔混交林coniferous and broad-leaved forest |
按化学基团的性质将Eco板上的31种碳源分成6类:碳水化合物类、氨基酸类、酯类、醇类、胺类、羧酸类。6类碳源均呈现出随着培养时间的延长, 微生物利用碳源的量逐渐增加的趋势。由表 3可知, 随着海拔上升, 除了氨基酸类和醇类外, 不同植被带土壤微生物对其他4类碳源利用率差异显著。不同植被带土壤微生物对不同碳源利用率呈现一致的规律, 即EBF>DBF>CBF。总体来说, 氨基酸类、胺类和酯类为各植被带土壤微生物的主要碳源, 其次为碳水化合物类和羧酸类, 醇类的利用率最小。
| 植被带 Vegetation belts |
碳水化合物类 Carbohydrate |
氨基酸类 Amino acid |
酯类 Ester |
醇类 Alcohol |
胺类 Amine |
羧酸类 Carboxylic acid |
| 常绿阔叶林EBF | 0.780±0.116a | 1.270±0.219a | 1.106±0.094a | 0.654±0.061a | 1.213±0.153a | 0.800±0.044a |
| 落叶阔叶混交林DBF | 0.591±0.067a | 0.926±0.336a | 0.758±0.155bc | 0.483±0.178a | 0.697±0.420b | 0.560±0.014b |
| 针阔混交林CBF | 0.576±0.088a | 0.925±0.067a | 0.677±0.136c | 0.622±0.021a | 0.514±0.066b | 0.549±0.006b |
| 同一列数据中不同字母表示差异显著(P<0.05) | ||||||
碳源利用的丰富度、均匀度和多样性等指数用来描述在颜色变化率一致的情况下, 土壤微生物群落利用碳源的程度, 也反应了微生物群落的功能多样性。根据培养96 h的AWCD值计算得到不同植被带土壤微生物群落的Shannon指数、丰富度指数和均匀度指数(表 4)。结果表明, 随着海拔上升, 各植被带Shannon指数和丰富度指数差异显著, 而均匀度指数差异不显著。
| 植被带 Vegetation belts |
Shannon指数 Shannon index |
丰富度指数 Richness |
均匀度指数 Evenness |
| 常绿阔叶林EBF | 3.07±0.03a | 20.00±3.46a | 1.02±0.04a |
| 落叶阔叶混交林DBF | 2.97±0.04b | 12.00±5.57b | 1.29±0.34a |
| 针阔混交林CBF | 2.61±0.05c | 12.33±0.58b | 1.14±0.01a |
| 同一列数据中不同字母表示差异显著(P<0.05) | |||
根据培养96 h的AWCD值, 对不同海拔植被带土壤微生物对单一碳源的利用特性进行主成分分析。从图 2可以看出, 第1主成分的贡献率为40.42%, 第2主成分的贡献率为15.97%, 前两个主成分累计解释了变量方差的56.39%。在PC1方向, 不同海拔植被带具有明显的分异, EBF位于正端, 得分系数在0.75—1.81之间;而DBF和CBF均位于负端, 得分系数在0.01—-0.92之间。在PC2方向, EBF在正端和负端均有分布, DBF全部位于负端, 而CBF全部位于正端。将主成分得分系数与单一碳源AWCD值做相关分析, 在31种碳源中与PC1显著相关的有17个, 其中16个呈正相关, 主要是酯类、羧酸类、醇类和碳水化合物类碳源。与PC2显著相关的碳源有5个, 均呈正相关, 主要为氨基酸类。可见, 在主成分分离中起主要贡献作用的是酯类、胺类和氨基酸类碳源。

|
| 图 2 不同海拔梯度植被带土壤微生物群落的碳代谢主成分分析 Fig. 2 Principle component analysis for carbon metabolism of soil microbial communities in different vegetation belts along an elevation gradient |
土壤理化性质与土壤微生物生存的环境条件之间有紧密的联系。为了探讨土壤理化性质与微生物群落多样性之间的关系, 进行了土壤理化性质与微生物群落多样性的相关性分析(表 5)。由表 5可知, 土壤微生物群落功能多样性Shannon指数与土壤pH、TP和TK之间呈正相关关系, 与含水量、TOC、TN、AN、AP和AK等呈负相关关系。其中, Shannon指数与TP和AK以外的其他土壤理化性质之间的相关性均达到显著或极显著水平。丰富度指数和均匀度指数与土壤理化性质之间相关性均不显著。由此可见, 土壤理化性质与土壤微生物群落Shannon指数密切相关, 是造成不同海拔植被带土壤微生物群落多样性差异的重要原因。
| 功能多样性指数 Functionaldiversity indexes |
pH | 含水量 Water content |
总有机碳 TOC |
全N TN |
全P TP |
全K TK |
速效N AN |
速效P AP |
速效K AK |
| Shannon指数Shannon index | 0.822** | -0.859** | -0.772* | -0.740* | 0.185 | 0.905** | -0.822** | -0.850** | -0.648 |
| 丰富度指数Richness | 0.519 | -0.360 | -0.254 | -0.196 | 0.587 | 0.501 | -0.289 | -0.362 | -0.021 |
| 均匀度指数Evenness | -0.228 | -0.088 | -0.131 | -0.176 | -0.559 | -0.093 | -0.101 | 0.045 | -0.302 |
| *P<0.05;**P<0.01 | |||||||||
随着海拔梯度的上升, 植被类型、气候等环境因子也发生改变, 进而造成土壤养分特征的差异并呈现一定的规律性。本研究中, 不同海拔土壤理化性质的测定结果表明, 随着海拔的上升, pH和TK表现出递减的趋势, 而土壤含水量表现出逐渐增加的趋势。土壤含水量与降雨量、温度和坡度等因子相关, 随着海拔上升, 降雨量逐渐增大而气温逐渐下降[10], 故土壤含水量表现出逐渐增加的趋势。TOC、TN、AN、TP、AP和AK随海拔梯度上升均呈现先下降后上升的趋势, 其在不同海拔植被带的变化规律相同, 即CBF>EBF>DBF。TOC、TN、AN、AP、AK等土壤养分含量在最高海拔处最高, 可能与地上植被的凋落物输入有关[26], 且受温度的影响[27]。随着温度的降低, 土壤微生物生命活动变弱, 凋落物的分解变慢, 从而增加了土壤中养分的积累。
3.2 海拔变化对土壤微生物群落功能多样性的影响森林生态系统中影响土壤微生物群落多样性的因素有很多, 海拔是其中一个重要因素。海拔梯度发生变化, 植被类型、气候等环境因子也发生改变, 进而引起土壤微生物群落多样性的变化[15]。斯贵才等[28]针对念青唐古拉山不同海拔的高寒沼泽土壤微生物群落和酶活性进行研究, 发现土壤微生物生物量随海拔增加而明显减小, 应用相关性分析与典范对应分析, 结果表明随海拔上升而递减的年平均气温是影响念青唐古拉山区域土壤微生物群落结构的重要因子。丛静等[29]对神农架自然保护区土壤微生物生物量沿海拔梯度的变化进行研究, 发现土壤微生物生物量随海拔的升高而显著增加, 随海拔梯度升高而变化的土壤温湿度、pH、有机碳和全氮是重要的影响因子。本研究结果表明, 随着海拔的上升, 土壤微生物群落的AWCD值逐渐减小, 与吴则焰等[17]的研究结果一致。土壤微生物的Shannon指数、丰富度指数的总体趋势是EBF最高, DBF次之, CBF最低。氨基酸类、胺类和酯类为各植被带土壤微生物的主要碳源, 其次为碳水化合物类和羧酸类, 醇类的利用率最小。与碳源利用相关的两个主成分分别能解释变量方差的40.42%和15.97%, 在主成分分离中起主要贡献的是酯类、氨基酸类和胺类碳源。酯类碳源主要是糖类和氨基酸的代谢中间物, 而糖类是自然界矿化的主要物质, 氨基酸是土壤有机氮的重要组成部分, 是植物根系向土壤介质中分泌的低分子有机化合物之一[30]。胺类在自然界中分布很广, 大多数是由氨基酸脱羧生成的。朱彪等[31]研究了猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局, 结果发现海拔1350 m以上物种丰富度和多样性表现出明显的随海拔梯度的上升而下降的趋势, 其中海拔较坡度和坡向对群落的丰富度和多样性影响更大。可见, 海拔梯度的变化引起植被类型的改变, 影响植物根系的分泌物, 导致土壤理化性质发生改变, 进而对土壤微生物群落功能多样性产生影响。
3.3 土壤理化性质与土壤微生物群落功能多样性的相关性土壤微生物群落多样性与土壤理化性质和植被状况有关[32]。吴则焰等[17]对武夷山不同海拔植被带的土壤微生物群落多样性进行研究, 发现土壤养分含量随海拔梯度上升而递减, 是造成不同海拔土壤微生物群落多样性差异的重要原因。本研究结果表明, 随着海拔上升pH和TK呈下降趋势, 而微生物群落多样性的Shannon指数与TK呈极显著正相关, 与含水量、TOC、TN、AN、AP之间的相关性为显著或极显著, 且为负相关。丰富度指数和均匀度指数与各土壤理化性质之间的相关性均不显著。土壤含水量影响土壤颗粒中的氧气条件, 进而影响了相关土壤中微生物的活性[33]。随着海拔升高, 猫儿山不同海拔植被带土壤的含水量呈上升趋势, 尤其针阔混交林林地土壤为山地沼泽泥炭土, 长时间的潮湿引起了微生物呼吸和生物量的降低, 进而引起土壤微生物功能多样性的降低。土壤TK主要由成土母质决定的。Deng等[34]对我国亚热带地区红壤的微生物群落结构和功能多样性进行研究, 发现成土母质较土地利用方式对亚热带地区土壤微生物群落的影响更大。魏佳宁等[35]研究了三江源区土壤微生物和土壤养分的空间分布规律, 发现土壤微生物群落数量表现为细菌>放线菌>真菌, 而土壤全钾是影响细菌数量的主要因子。由于本研究所采集的土壤样品属于表层土, 其受凋落物组成、环境因子等的影响较大。土壤TK含量和含水量与微生物群落多样性Shannon指数呈极显著相关关系, 可见, 土壤TK含量和含水量可能是造成不同海拔土壤微生物群落功能多样性差异的主要原因。
综上所述, 随着海拔梯度上升, 土壤理化性质和土壤微生物群落功能多样性的变化呈现了一定的规律性。土壤微生物群落AWCD值随海拔梯度逐渐减小, 多样性指数总体趋势是EBF>DBF>CBF, 在主成分分离中起主要贡献的是羧酸类和酯类碳源。土壤TK随海拔上升呈下降趋势, 其与微生物群落多样性的Shannon指数之间呈极显著的正相关。土壤TK随海拔梯度上升而递减, 可能是造成不同海拔微生物群落功能多样性差异的主要原因。土壤微生物是一个庞大且复杂的群体, 是森林生态系统的重要组成部分。研究土壤微生物群落多样性对海拔梯度变化的响应, 对于揭示微生物、环境、植物之间的关系意义重大。本研究仅利用Biolog微平板技术分析了夏季的土壤微生物群落功能多样性随着海拔梯度变化的规律, 后续可以综合分子生物学和土壤酶学的研究方法, 将有助于进一步揭示中亚热带森林生态系统土壤微生物群落变化规律和微生物多样性与环境因子之间的相关性。
致谢: 感谢中山大学孙孝林副教授和南京师范大学邓欢副教授对写作的帮助, 感谢广西大学生命科学与技术学院冯家勋教授课题组在Biolog分析上给予的帮助。| [1] | Bell T, Newman J A, Silverman B W, Turner S L, Lilley A K. The contribution of species richness and composition to bacterial services. Nature, 2005, 436(7054): 1157–1160. DOI:10.1038/nature03891 |
| [2] | Fierer N, Jackson R B. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(3): 626–631. DOI:10.1073/pnas.0507535103 |
| [3] | Zhang X, Zhao X, Zhang M. Functional diversity changes of microbial communities along a soil aquifer for reclaimed water recharge. FEMS Microbiology Ecology, 2012, 80(1): 9–18. DOI:10.1111/j.1574-6941.2011.01263.x |
| [4] | 周玲莉, 姚斌, 向仰州, 尚鹤, 韦秀文, 刘方. 五氯酚胁迫对杨树生长及根际微生物群落的响应特征. 林业科学, 2010, 46(10): 62–68. DOI:10.11707/j.1001-7488.20101010 |
| [5] | 刘晶鑫, 迟凤琴, 许修宏, 匡恩俊, 张久明, 宿庆瑞, 周宝库. 长期施肥对农田黑土微生物群落功能多样性的影响. 应用生态学报, 2015, 26(10): 3066–3072. |
| [6] | 朱平, 陈仁升, 宋耀选, 刘光琇, 陈拓, 张威. 祁连山不同植被类型土壤微生物群落多样性差异. 草业学报, 2015, 24(6): 75–84. DOI:10.11686/cyxb2014272 |
| [7] | 宋贤冲, 杨中宁, 曹继钊, 张照远, 项东云. 萌芽更新对桉树根际土壤微生物群落功能多样性的影响. 桉树科技, 2014, 31(3): 36–40. |
| [8] | 吴林坤, 黄伟民, 王娟英, 吴红淼, 陈军, 秦贤金, 张重义, 林文雄. 不同连作年限野生地黄根际土壤微生物群落多样性分析. 作物学报, 2015, 41(2): 308–317. |
| [9] | 郑华, 欧阳志云, 方治国, 赵同谦. BIOLOG在土壤微生物群落功能多样性研究中的应用. 土壤学报, 2004, 41(3): 456–461. DOI:10.11766/trxb200309010320 |
| [10] | 黄金玲, 蒋得斌. 广西猫儿山自然保护区综合科学考察. 长沙: 湖南科技出版社, 2010: 1–9. |
| [11] | 李光照. 苗儿山植物区系的初步研究. 广西植物, 1985, 5(3): 211–226. |
| [12] | 粟通萍, 王绍能, 蒋爱伍. 广西猫儿山地区鸟类组成及垂直分布格局. 动物学杂志, 2012, 47(6): 54–65. |
| [13] | 段文军, 王金叶. 猫儿山自然保护区森林群落垂直格局与主导因素分析. 生态环境学报, 2013, 22(4): 563–566. |
| [14] | 蒋得斌, 吴兴亮, 李光平, 王绍能, 李泰辉, 宋斌, 邓春英, 邹方伦, 黄浩, 邓大军. 广西猫儿山国家级自然保护区大型真菌资源研究. 贵州科学, 2010, 28(1): 1–11. |
| [15] | 何容, 汪家社, 施政, 方燕鸿, 徐自坤, 权伟, 张增信, 阮宏华. 武夷山植被带土壤微生物量沿海拔梯度的变化. 生态学报, 2009, 29(9): 5138–5144. |
| [16] | 刘秉儒, 张秀珍, 胡天华, 李文金. 贺兰山不同海拔典型植被带土壤微生物多样性. 生态学报, 2013, 33(22): 7211–7220. |
| [17] | 吴则焰, 林文雄, 陈志芳, 方长旬, 张志兴, 吴林坤, 周明明, 陈婷. 中亚热带森林土壤微生物群落多样性随海拔梯度的变化. 植物生态学报, 2013, 37(5): 397–406. |
| [18] | Shen C C, Xiong J B, Zhang H Y, Feng Y Z, Lin X G, Li X Y, Liang W J, Chu H Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biology & Biochemistry, 2013, 57(3): 204–211. |
| [19] | 国家林业局. LY/T1952-2011森林生态系统长期定位观测方法[S]. 北京: 中国标准出版社, 2011. |
| [20] | 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000. |
| [21] | 国家林业局. LY/T1210-1275-1999森林土壤分析方法. 北京: 中国标准出版社, 1999. |
| [22] | Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology, 1991, 57(8): 2351–2359. |
| [23] | Garland J L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biology & Biochemistry, 1996, 28(2): 213–221. |
| [24] | 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. DOI:10.17520/biods.1994027 |
| [25] | Ratcliff A W, Busse M D, Shestak C J. Changes in microbial community structure following herbicide (glyphosate) additions to forest soils. Applied Soil Ecology, 2006, 34(2/3): 114–124. |
| [26] | 陈法霖, 郑华, 欧阳志云, 张凯, 屠乃美. 土壤微生物群落结构对凋落物组成变化的响应. 土壤学报, 2011, 48(3): 603–611. DOI:10.11766/trxb201005130189 |
| [27] | Salinas N, Malhi Y, Meir P, Silman M R, Román-Cuesta R M, Huaman J, Salinas D, Huaman V, Gibaja A, Mamani M, Farfan F. The sensitivity of tropical leaf litter decomposition to temperature:results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New Phytologist, 2011, 189(4): 967–977. DOI:10.1111/nph.2011.189.issue-4 |
| [28] | 斯贵才, 王建, 夏燕青, 袁艳丽, 张更新, 雷天柱. 念青唐古拉山沼泽土壤微生物群落和酶活性随海拔变化特征. 湿地科学, 2014, 12(3): 340–348. |
| [29] | 丛静, 刘晓, 卢慧, 薛亚东, 刘学端, 尹华群, 李迪强, 张于光. 神农架自然保护区土壤微生物生物量碳、氮沿海拔梯度的变化及其影响因素. 生态学杂志, 2014, 33(12): 3381–3387. |
| [30] | 孔滨, 杨秀娟. Biolog生态板的应用原理及碳源构成. 绿色科技, 2011(7): 231–234. |
| [31] | 朱彪, 陈安平, 刘增力, 李光照, 方精云. 广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 44–52. |
| [32] | 王光华, 金剑, 徐美娜, 刘晓冰. 植物、土壤及土壤管理对土壤微生物群落结构的影响. 生态学杂志, 2006, 25(5): 550–556. |
| [33] | 胡亚林, 汪思龙, 颜绍馗. 影响土壤微生物活性与群落结构因素研究进展. 土壤通报, 2006, 37(1): 170–176. |
| [34] | Deng H, Yu Y J, Sun J E, Zhang J B, Cai Z C, Guo G X, Zhong W H. Parent materials have stronger effects than land use types on microbial biomass, activity and diversity in red soil in subtropical China. Pedobiologia, 2015, 58(2/3): 73–79. |
| [35] | 魏佳宁, 马红梅, 邵新庆, 张建全, 余群, 柴琦, 钟梦莹. 三江源区土壤微生物和土壤养分空间分布特性研究. 中国土壤与肥料, 2016(2): 27–31. DOI:10.11838/sfsc.20160205 |