 2017, Vol. 37
2017, Vol. 37文章信息
- 陈吉龙, 何蕾, 温兆飞, 寥华军, 王炳亮, 崔林林, 李国胜
- CHEN Jilong, HE Lei, WEN Zhaofei, LIAO Huajun, WANG Bingliang, CUI Linlin, LI Guosheng.
- 辽河三角洲河口芦苇沼泽湿地植被固碳潜力
- Carbon sequestration potential of a Phragmites salt marsh in the Liaohe River estuarine wetland
- 生态学报. 2017, 37(16): 5402-5410
- Acta Ecologica Sinica. 2017, 37(16): 5402-5410
- http://dx.doi.org/10.5846/stxb201605241004
-
文章历史
- 收稿日期: 2016-05-24
- 网络出版日期: 2017-03-27





2. 中国科学院地理科学与资源研究所, 北京 100101;
3. 鄱阳湖湿地与流域研究教育部重点实验室, 南昌 330000;
4. 中国地质调查局滨海湿地生物地质重点实验室, 青岛 266071
2. Institute of Geographic Sciences and Natural Resources Research, Beijing 100101, China;
3. Key Laboratory of Poyang Lake Wetland and Watershed Research, Ministry of Education, Nanchang 330000, China;
4. Key Laboratory of Coastal Wetland Biogeosciences, China Geologic Survey, Qingdao 266071, China
由温室气体浓度升高而导致的全球变暖受到广泛影响, 固碳和减排已成为国际社会的共识。《京都议定书》明确提出增加陆地生态系统的碳汇是一种非常有效的固碳和减排机制[1]。为了能在气候变化谈判中取得优势, 各国对陆地生态系统固碳潜力进行了深入的研究。最初《京都议定书》只认可通过造林、再造林、森林管理、农地管理、草地管理以及植被恢复等人为活动增加的碳汇来抵消温室气体排放, 湿地暂不在列, 因而对固碳潜力研究集中于森林[2-3]、草地[4-5]和农田生态系统[6-7], 而湿地固碳潜力并未引起重视。
湿地是水陆相互作用形成的独特陆地生态系统, 虽然只占全球陆地面积的6%[8], 但是高的净初级生产力(NPP), 以及水淹形成的还原环境抑制了有机物的分解[9], 碳在土壤中大量累积, 形成了陆地生物圈碳库的最大组成部分[10]。近年来湿地的碳汇功能引起了人们的关注, 政府间气候变化专门委员会也决定对清单指南进行补充, 并发布了《2006年IPCC国家温室气体清单指南2013年增补:湿地》。可见, 湿地的碳汇功能已引起了重视, 研究湿地固碳潜力也必将成为碳循环研究的热点方向。
尽管如此, 目前仍缺乏一个广泛通用的湿地固碳潜力的定义及计算方法。湿地生态系统的固碳能力受到湿地面积、NPP、异氧呼吸以及湿地土壤碳库等因素的影响, 则湿地生态系统的固碳潜力就是在最适环境因素共同作用下, 湿地固碳能力达到的理论最大值。由于湿地具有很高的NPP, 增加湿地面积能够固定更多碳[11], 通过面积增加的潜力和固碳速率便可获得湿地固碳潜力, 这也是目前计算固碳潜力最常用的方法。段晓男利用此方法研究了我国不同类型湿地的固碳潜力, 结果表明我国不同类型的湿地都具有可观的固碳潜力[12]。另一方面, 由于气候变化和不合理的人为干扰引起湿地水文等结构和功能变化, 造成了现存大部分湿地NPP的下降。黄河三角洲的芦苇湿地由于受水的制约, 灌溉季节得不到及时的灌溉, 严重制约了芦苇的产量[13]。对这些因缺水而面临退化的湿地, 人工灌溉补水是恢复湿地功能的重要措施[14]。监测数据表明人为灌溉补水能明显提高芦苇沼泽湿地的生产力[15-16]。对扎龙湿地灌溉前后的芦苇形态结构对比研究表明, 人工灌溉明显促进了芦苇的生长[17]。James等发现恢复的泥炭湿地的群落生产力可以恢复到自然湿地的水平, 固碳能力达到了自然泥炭湿地的2倍[18]。在北美森林湿地, 通过水文重建恢复退化的森林湿地, 固碳能力在短时间内明显提高[19]。在我国的黄河三角洲河口湿地, 通过多种措施恢复湿地的水文条件, 使退化的芦苇沼泽湿地得到很好的恢复, 生产力也明显增加[20]。由此可见, 通过合理的人工湿地管理措施来恢复湿地水文等过程, 提高现存湿地的NPP, 也能够增强生态系统的碳汇, 而这种管理措施在湿地生态系统固碳潜力中往往被忽略。
湿地生态系统的固碳潜力主要包括植被NPP和土壤碳库两部分, 而植被的NPP又是土壤碳库的最重要来源。不同的环境因素对两部分的作用机制也不同, 有利于增强植被NPP的环境因素有可能增加土壤碳的排放, 比如温度升高能够提高植被NPP, 但同时异氧呼吸碳排放也会急剧增加。因而模拟湿地生态系统的固碳潜力较为复杂, 而分别计算湿地植被和土壤固碳潜力则较易实现。鉴于此, 本文对辽河三角洲芦苇沼泽湿地植被的固碳潜力进行剖析和定量计算, 这对全面研究湿地生态系统的固碳潜力具有重要意义, 也为研究类似河口湿地的固碳潜力提供参考和借鉴。
辽河三角洲河口湿地是我国重要的河口湿地之一, 是亚洲最大的暖温带滨海湿地[21], 已建成由东郭、羊圈子、赵圈河和南圈河构成的世界第二大苇场[22], 修建了完善的水利排灌体系, 人工灌溉成为管理芦苇湿地主要措施。灌溉不仅为芦苇补充了水分, 而且带走了多余的土壤盐分[23], 从而影响着芦苇沼泽群落的固碳能力。但由于种种原因, 对湿地的研究和监测开展较晚, 湿地生态系统缺少长期、系统的观测, 湿地的研究还主要停留在景观层面上[24], 对其固碳潜力还缺乏定量的认识。研究其植被固碳潜力, 对全面研究河口湿地碳循环以及挖掘我国湿地的固碳潜力都具有重要的现实意义。
1 研究方法 1.1 研究区域辽河三角洲河口湿地位于辽宁省西南部辽河平原南端, 渤海辽东湾的顶部, 西、北邻锦州, 东界鞍山, 南临渤海辽东湾(图 1), 地理位置介于121°30′—122°24′ E, 40°21′—41°21′ N, 是由辽河、双台子河、大凌河、小凌河、大清河等一系列河流作用形成的冲海积平原。气候属于暖温带大陆性半湿润季风气候。四季分明, 雨热同季, 年平均气温为8.6℃, 年平均降水量为631 mm, 主要集中在7—9月, 全年日照时数为2768 h, 其季节分布是春季大于秋季[25]。受海洋和陆地交互、咸淡水交汇作用, 复杂动力机制造就了复杂多样的湿地类型, 发育了以芦苇(Phragmites australis)、翅碱蓬(Suaeda pterantha)、獐毛(Aeluropus sinensis)、香蒲(Typha orientalis Presl)等作为建群种的湿地植物群落;其中芦苇沼泽面积达到756 km2, 是亚洲第一大芦苇湿地[22]。

|
| 图 1 辽河三角洲河口湿地的位置及样点的分布(背景为TM321合成真彩色影像) Fig. 1 Location of the sampling sites in Liaohe river estuarine wetland |
根据辽河三角洲河口湿地实际情况, 将植被固碳潜力分为湿地演替、人工灌溉和气候变化影响的潜力。
1.2.1 湿地演替固碳潜力的计算方法湿地演替固碳潜力包括两方面, 一方面河口湿地位于河流入海口, 河流所携带的泥沙淤积使湿地面积呈现增长趋势。湿地面积增长的固碳潜力可表示为[12]:
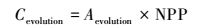
|
(1) |
式中,Cevolution表示湿地演替的固碳潜力, NPP表示不同演替阶段各植被群落的净初级生产力(利用野外样方调查数据), Aevolution表示面积增长的潜力, 利用潮滩淤进速率和时间计算, 淤进速率利用多年历史观测数据, 同时考虑海平面上升对潮滩的侵蚀作用, 利用布容法则计算侵蚀速率, 计算公式为:R=S×cotθ[26], 式中R为海岸线后退速率, S为海平面上升速率(利用已有的预测结果), θ为近海滩剖面的平均坡度(采用已有调查结果)。
湿地演替固碳潜力还包括植被演替引起的NPP变化。由于陆地、海洋和河流相互作用, 复杂的动力过程引起水分和盐度的分异[27], 使辽河三角洲河口湿地呈现图 2所示的演替过程, 以翅碱蓬群落→芦苇、獐毛草甸→芦苇沼泽正向演替过程最为典型。演替初期翅碱蓬群落NPP要低于芦苇、獐毛草甸群落, 而芦苇、獐毛草甸又要低于芦苇沼泽, 这种植被正向演替也会提高湿地固碳能力;通过两类植被NPP的差异和面积便可以计算出这类固碳潜力。

|
| 图 2 辽河三角洲河口湿地演替示意图 Fig. 2 Sketch map of wetland succession for Liaohe river estuarine wetland |
人工灌溉苇田的固碳潜力是通过模拟NPP与灌溉之间的关系, 获取在最佳灌溉措施下的潜在固碳速率, 结合灌溉面积计算。但各苇场灌溉数据非常缺乏, 难以构建NPP与灌溉之间的关系模型。在辽河三角洲河口湿地, 人工灌溉是通过改变水分和盐度影响芦苇沼泽的NPP[27]。因此, 人工灌溉苇田的潜在NPP可通过构建水分、盐度和NPP的关系模型模拟获取, 这与通过NPP与灌溉之间的关系获取潜在NPP在本质上是一致的。人工灌溉苇田的固碳潜力用公式表示为:
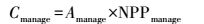
|
(2) |
式中,Cmanage表示人工灌溉芦苇湿地的固碳潜力, Amanage表示灌溉苇田面积, NPPmanage表示芦苇沼泽在最佳灌溉措施下可能达到的最大净初级生产力, 通过不同水盐梯度样点的数据, 构建水分、盐度与NPP的关系模型模拟获取。
1.2.3 气候背景下芦苇沼泽的固碳潜力计算未来气候对芦苇沼泽NPP的影响是利用CASA模型[28-29], 以遥感获取植物光合生理参数, 结合气候观测数据, 模拟芦苇沼泽的NPP。假定其它因素不变的情况下, 利用各文献报道的辽河三角洲气候变化情景, 以气温、降水、辐射为变量模拟出芦苇沼泽湿地的NPP, 并结合对应时间的湿地面积, 计算未来气候变化情景下芦苇沼泽湿地的固碳潜力。CASA构架可表示为[28-29]:

|
(3) |
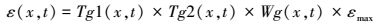
|
(4) |
式中, APAR(Absorbed photosyn-thetically active radiation)为吸收光合有效辐射, 利用太阳总辐射量和归一化植被指数NDVI计算[30]。εmax为最大光能转化率利用已有报道的模拟值[31], Tg1和Tg2为温度胁迫系数, 利用月平均气温和NDVI计算, Wg为水分胁迫系数, 根据Food and Agriculture Organization(FAO)推荐的作物潜在蒸散的Penman-Monteith方程[32]和Boucher提出的互补关系计算[33]。
1.3 野外调查 1.3.1 不同演替阶段样点设置与调查不同演替阶段各植被群落的NPP通过野外样方调查获得。在对湿地全面考查的基础上, 根据植被演替规律, 选择不同演替阶段有代表性的植被群落。每个群落中设置一个大样地(图 1, 样点1—3), 大样地面积设为30 m×30 m, 每一样地的四角和中心设置5个2 m×2 m小样方。在生长季结束时按收获法调查样方植被的地上总鲜重, 并取样品一份(300—500 g左右), 用于测定干物质比率。同时收集样方中所有调落物并测定鲜重, 取样品一份, 测算调落物的干物质比率。将样品带回实验室烘干48 h(80℃)至恒重并称重, 然后根据干物质比率及其总鲜重, 推算样方内地上总干重, 以5个小样方的平均值代表样地单位面积生物量地上部分, 最后乘以碳转换系数(0.45), 换算成单位面积植被地上NPP。
1.3.2 不同水盐梯度样点设置与调查根据辽宁省下辽河平原水文地质普查报告, 湿地盐度由内陆向滨海地区递增, 盐化程度逐渐加重[34]。从滨海向内陆地区, 潮水的影响逐渐减弱, 土壤水分也发生规律性变化。根据此规律, 在芦苇沼泽湿地核心区域, 不同的水盐梯度下, 设置了6个群落样点(图 1, 样点4—9), 采用与湿地不同演替阶段NPP调查相同的方法, 调查各个芦苇沼泽群落样点的地上NPP。同时在每个样点利用内径为10 cm、长为50 cm不锈钢土壤取样器, 钻取40 cm深的土壤样品, 利用三针型土壤参数仪(WET Sensor)测定土壤盐度和水分(图 3)。利用这些样点的数据, 构建芦苇沼泽NPP与土壤水分、盐度的数值模型, 模拟出理想水盐条件下芦苇沼泽NPP的最大值。

|
| 图 3 土壤样品采集与环境因素测定 Fig. 3 Soil sampling and measurements of environmental factors |
部分NPP在生长期分配到根部, 经过累积形成地下生物量。由于芦苇具有发达的深根系, 难以采集地下生物量, 加上难以从根系生物量中分离当年NPP。因此, 本文样地调查仅测定了植被地上部分生物量, 采用根、茎、叶动态配置模式(Pierre′s model), 根据可利用资源光(L)、水分(W)和营养(N)对不同器官进行碳的动态配置[35], 模拟出不同植被地上和地下部分的NPP分配模式, 并利用测定的植被地上NPP, 计算总NPP。
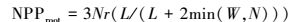
|
(5) |
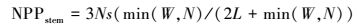
|
(6) |

|
(7) |
式中, Nr和Ns代表没有资源限制时, NPP分配到根和茎中的比率, 默认取值0.3。资源可利用性因子L、W、N取值介于0.1(资源限制)与1(资源充分并容易获取)之间。L利用叶面积指数来估算, L=e-K×LAI, K为消光系数, 默认取值为0.5。水分因子W取决于土壤湿度和土壤质地。假营养因子N利用湿度和气温来计算, N=Fp×Ft, 其中Fp=PPT/PET, Ft=2[(T-30)/10], PPT为降水量, T为平均气温, PET潜在蒸散, 根据Boucher提出的互补关系计算[33]。
2 结果 2.1 湿地演替的固碳潜力已有研究及多年监测结果表明, 辽河三角洲滩涂淤涨速率为100—150 m/a[36-37], 利用布容法则、坡降调查[38]和海平面变化研究结果[39-40]计算海岸线后退速率为11—31 m/a, 则潮滩淤涨速率约为90—120 m/a, 结合潮滩海岸线长度, 滩涂面积递增潜力为3.78—5.04 km2/a, 按照目前滩涂上翅碱蓬生长比例计算, 翅碱蓬面积递增潜力为0.49—0.65 km2/a, 结合野外调查的翅碱蓬群落平均NPP 109.03 g C/m2计算, 翅碱蓬面积扩张可递增固碳潜力为0.054—0.071 GgC/a。
野外调查发现湿地从初期翅碱蓬演替到中期芦苇、獐毛草甸的面积非常有限, 大部分被人工改造成芦苇沼泽。遥感监测结果显示1988—2006年间, 平均每年有0.076 km2的滩涂转变为芦苇沼泽[27, 32], 其中翅碱蓬面积为0.048 km2[24]。按此速率结合调查的芦苇沼泽平均NPP 994.6 gC/m2计算, 总NPP可由0.0053 GgC/a提高到0.076 GgC/a, 可递增固碳潜力0.07 GgC/a。在苇场灌溉条件较差的地方, 芦苇沼泽明显退化, 獐毛、碱篷大量生长。通过人工灌溉将这部分草甸改造成为芦苇沼泽, 野外调查的芦苇、獐毛草甸平均NPP为369.73 gC/m2, 总NPP可由10.08 GgC/a提高到27.28 GgC/a, 可新增固碳潜力17.2 GgC/a。
2.2 人工灌溉苇田的固碳潜力辽河三角洲芦苇沼泽湿地具有很高的固碳能力, 平均NPP达到940—1037 gC/m2。在水分和盐度条件较好的地方, NPP超过1200 gC/m2。按照火电厂发1度电需要消耗400标准煤, 同时排放997克CO2计算, 芦苇沼泽湿地每年固定的CO2相当于年发电量为27.2—30.2亿度的火电厂一年排放量, 相当于节约了108.9—120.7万t标准煤, 由此可见辽河三角洲芦苇沼泽湿地的巨大减排能力。
然而实际样点水分和盐度并不一定是芦苇沼泽生长的最理想环境, 因此, 通过适宜的灌溉管理措施调节芦苇沼泽的水分和盐度, NPP还有提升的空间。通过模拟样点的NPP与土壤水分、盐度之间的关系计算得出, 在理想水盐条件下, 辽河三角洲芦苇沼泽湿地的NPP可达到1700 gC/m2[34]。由于东郭、羊圈子、赵圈河和南圈河四大苇场都位于双台子河口自然保护区范围内, 假定它们的景观类型不发生转变, 则通过人工灌溉管理措施, 四大苇场的总NPP可提高到1234.63 GgC/a, 增长62.5%—78.9%, 可新增固碳潜力474.6—544.6 GgC/a, 。
除了4个大型的苇场外, 在大辽河、辽河以及绕阳河的沿岸也分布有部分苇田。另外, 潮滩面积扩大每年也会有部分滩涂转变为苇田。对这部分苇田也实行人工灌溉管理措施, 到2030年, NPP可提高68.4—74.9 GgC/a;到2050年, NPP可提高69.5—75.9 GgC/a;到2100年, NPP可提高72.1—78.6 GgC/a。
2.3 气候变化情景下芦苇沼泽的固碳潜力根据已有气候变化研究及预测结果, 辽河三角洲2011—2030年平均增温约0.6℃[41], 太阳辐射和降水量的变化不明显;2011—2050平均增温约1.8℃[42];太阳总辐射增幅约为20 MJ m-2 10a-1);降水量下降幅度约为15—20 mm/10a[43];2100年平均气温将增加约3℃, 年降水量将增加约100 mm[44]。以此气候变化情景为基础, 利用CASA模型模拟结果表明芦苇沼泽NPP在2011—2100年呈增加趋势, 到2030年, 芦苇沼泽湿地的总NPP可达784.9—862.8GgC/a, 新增固碳潜力14.3—15.7GgC/a;到2050年, 芦苇沼泽的总NPP可达809.3—889.4 GgC/a, 新增固碳潜力37.2—40.9 GgC/a;到2100年, 芦苇沼泽的总NPP可达881.9T—968.8 GgC/a, 新增固碳潜力106T—116.4 GgC/a。不同潜力大小见表 1。
| 潜力来源Classification | 2030年 | 2050年 | 2100年 | |
| 湿地演替 | 翅碱蓬沼泽扩张 | 1.01—1.35 | 2.07—2.77 | 4.73—6.31 |
| Wetland plant succession | 芦苇、獐毛草甸转变为芦苇沼泽 | 17.2 | 17.2 | 17.2 |
| 滩涂转变为芦苇 | 1.34 | 2.74 | 6.26 | |
| 人工灌溉Irrigation to Phragmites fields | 543.0—619.1 | 544.1—620.6 | 546.7—623.2 | |
| 气候变化Influence of the climate change | 14.3—15.73 | 37.2—40.9 | 106—116.4 | |
| 潜力小计Sum of carbon sequestration potential | 576.9—655.1 | 603.3—684.1 | 680.9—769.4 | |
| 净生产力总量Net primary productivity | 1330.11—1484.78 | 1356.51—1513.79 | 1434.14—1599.09 |
湿地具有很高的NPP, 因而增加湿地面积能固定更多碳。在淤涨型河口湿地, 由于河流所携带的泥沙淤积, 使潮滩面积逐年扩大, 湿地固碳能力逐年增长。但由于刚成陆的近海端环境恶劣, 先锋植物需要更多能量用于抵御环境的影响, 需要更多代价才能获得足够营养以维持正常生命活动。因此, NPP较低[45], 对固碳潜力贡献有限。在辽河三角洲河口湿地, 潮滩整体以淤进为主, 面积不断扩大[36], 固碳能力随着时间推移不断提高, 但对新增固碳潜力贡献较低。另一方面, 尽管未来气候变化会使芦苇沼泽群落NPP提高, 但气候变化也能引起风暴潮以及极端气候事件频率的增加, 对刚成陆的滩涂沼泽湿地形成极大威胁[46]。因此, 河口地区的淤积扩张对提高湿地固碳潜力具有一定的不确定性。
由于全球气候变化和不合理的人为干扰, 辽河三角洲河口湿地面临着退化的威胁, 影响了芦苇沼泽湿地的固碳能力。本文研究结果表明采取有效保护和人工灌溉管理措施, 芦苇沼泽的NPP可提高62.5%—78.9%, 表明合理的人工湿地管理措施具有十分可观的固碳潜力。辽河三角洲是我国著名的商品粮基地, 盘锦大米被冠以“中国名牌”和“中国驰名商标”等殊荣。因此, 水田需水历来得到政府的优先保证, 水田面积扩张对水的需求量大, 与苇田用水之间矛盾十分尖锐。近年来由于气候变暖, 径流大幅减少, 整个辽宁省处于普遍缺水状况, 分配给盘锦市的用水指标甚至还无法保证水田的需求。过去有1/2的苇田能够适时灌水, 近年来能适时灌水的苇田只有1/3左右[47]。因此, 上述所计算的人工灌溉芦苇的固碳潜力难以达到。假设按照目前有1/3—1/2的苇田能够有水源保障, 通过灌溉管理提高水资源的利用效率, 四大苇场的总NPP仍可提高26%—33%, 增加固碳潜力197.6—231.4GgC/a。
4 结论采用野外调查与模型模拟相结合的方法, 研究了辽河三角洲河口湿地演替、人工灌溉苇田的和气候变化的固碳潜力。河口湿地固碳能力随着湿地的正向演替而不断增强, 其中, 翅碱蓬面积扩张可递增固碳潜力0.053—0.071 GgC/a, 由滩涂转变为芦苇沼泽可递增固碳潜力为0.07 GgC/a, 而通过芦苇、獐毛草甸演替到芦苇沼泽可增加固碳潜力17.2 GgC/a。随着未来气温的升高有利于植被的光合作用, 气候变化也具有一定的固碳潜力, 对增强植被的碳汇有着积极的作用。通过人工灌溉管理措施, 四大苇场固碳潜力为474.6—544.6 GgC/a, 说明了人工灌溉管理措施对提高芦苇沼泽的固碳能力具有十分显著的效果, 为将湿地管理活动纳入减排和固碳机制提供了科学参考。
| [1] | IPCC (Intergovernmental Panel on Climate Change). Climate Change 2007:The Physical Science Basis. Contribution of Working Group to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Solomon S D, Qin M, Manning Z, Chen M, Marquis K B, Averyt M, Miller H L, eds. Cambridge, UK:Cambridge University Press, 2007. |
| [2] | 吴庆标, 王效科, 段晓男, 邓立斌, 逯非, 欧阳志云, 冯宗炜. 中国森林生态系统植被固碳现状和潜力. 生态学报, 2008, 28(2): 517–524. |
| [3] | Mehraj A S, Munesh K. Carbon sequestration potential of trees on two aspects in sub-tropical forest. International Journal of Conservation Science, 2010, 1(3): 143–148. |
| [4] | 曹广民, 龙瑞军, 张法伟, 李以康, 林丽, 郭小伟, 韩道瑞, 李婧. 青藏高原高寒矮嵩草草甸碳增汇潜力估测方法. 生态学报, 2010, 30(23): 6591–6597. |
| [5] | Wolf S, Eugster W, Potvin C, Turner B L, Buchmann N. Carbon sequestration potential of tropical pasture compared with afforestation in Panama. Global Change Biology, 2011, 17(9): 2763–2780. DOI:10.1111/gcb.v17.9 |
| [6] | Sun W J, Huang Y, Zhang W, Yu Y Q. Carbon sequestration and its potential in agricultural soils of China. Global Biogeochemical Cycles, 2010, 24(3): GB3001. |
| [7] | 覃章才, 黄耀. 基于模型的农田土壤固碳潜力估算. 中国科学:生命科学, 2010, 40(7): 658–676. |
| [8] | 崔保山, 杨志峰. 湿地学. 北京: 北京师范大学出版社, 2006. |
| [9] | 宋长春. 湿地生态系统碳循环研究进展. 地理科学, 2003, 23(5): 622–628. |
| [10] | Lal R. Carbon sequestration. Philosophical Transactions of the Royal Society B:Biological Sciences, 2007, 363: 815–830. |
| [11] | IPCC. Climate Change 2007:Mitigation. Contribution of Working Group Ⅲ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Metz B, Davidson O R, Bosch P R, Dave R, Meyer L A, eds. New York:Cambridge University Press, 2007:555-576. |
| [12] | 段晓男, 王效科, 逯非, 欧阳志云. 中国湿地生态系统固碳现状和潜力. 生态学报, 2008, 28(2): 463–469. |
| [13] | 刘慧超, 孟维芳, 杨学贤. 黄河三角洲芦苇定量补充灌溉效益分析. 农业技术经济, 1998(5): 61–64. |
| [14] | Henry CP, Amoios C. Restoration ecology of riverine wetlands:Ⅰ. A scientific base. Environmental Management, 1995, 19(6): 891–902. DOI:10.1007/BF02471940 |
| [15] | MauchampA, Blanch S, Grillas P. Effects of submergence on the growth of Phragmites australis seedlings. Aquatic Botany, 2001, 69(2/4): 147–164. |
| [16] | Engloner A I. Annual growth dynamics and morphological differences of reed (Phragmites australis[Cav.] Trin. ex Steudel) in relation to water supply. Flora-Morphology, Distribution, Functional Ecology of Plants, 2004, 199(3): 256–262. DOI:10.1078/0367-2530-00153 |
| [17] | 黄璞玮, 于洪贤, 柴龙会. 人为补水对扎龙湿地芦苇生长的影响. 东北林业大学学报, 2011, 39(1): 38–40. |
| [18] | Waddington J M, Warner K. Atmospheric CO2 sequestration in restored mined peatlands. Ecoscience, 2001, 8(3): 359–368. DOI:10.1080/11956860.2001.11682664 |
| [19] | Trettin C C, Jurgensen M F, Grigal D F, Gale M R, Jeglum J R. Northern Forested Wetlands, Ecology and Management. London:CRC Press, 1996: 311–331. |
| [20] | 唐娜, 崔保山, 赵欣胜. 黄河三角洲芦苇湿地的恢复. 生态学报, 2006, 26(8): 2616–2624. |
| [21] | 肖笃宁, 胡远满, 李秀珍. 环渤海三角洲湿地的景观生态学研究. 北京: 科学出版社, 2001. |
| [22] | 王西琴, 李力. 辽河三角洲湿地退化及其保护对策. 生态环境, 2006, 15(3): 650–653. |
| [23] | 汲玉河, 周广胜. 1988-2006年辽河三角洲植被结构的变化. 植物生态学报, 2010, 34(4): 359–367. |
| [24] | 黄桂林, 何平. 辽河三角洲湿地景观演变研究——以盘锦市为例. 林业资源管理, 2011(3): 82–89. |
| [25] | 王炳亮, 李国胜. 1961-2010年辽河三角洲参考蒸散发变化特征及主导因子分析. 地理科学, 2014, 34(10): 1233–1238. |
| [26] | Bruun P. Sea level rise as a cause of shore erosion. Journal of the Waterways and Harbors Division, 1962, 88: 117–130. |
| [27] | 汲玉河, 周广胜. 辽河三角洲植被生产功能的年际变化及其驱动因子分析. 植物生态学报, 2010, 34(7): 792–799. |
| [28] | Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production:a process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811–841. DOI:10.1029/93GB02725 |
| [29] | Field B C, Randerson J T, Malmström C M. Global net primary production:combining ecology and remote sensing. Remote Sensing of Environment, 1995, 51(1): 74–88. DOI:10.1016/0034-4257(94)00066-V |
| [30] | Roujean J L, Breon F M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote sensing of Environment, 1995, 51(3): 375–384. DOI:10.1016/0034-4257(94)00114-3 |
| [31] | 陈吉龙, 李国胜, 寥华军, 王炳亮, 崔林林. 辽河三角洲河口湿地典型芦苇群落最大光能转化率模拟. 生态学报, 2017, 37(7). DOI:10.5846/stxb201512162511 |
| [32] | Allen R G, Pereira L S, Raes D, Smith M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements. Rome:FAO Irrigation and Drainage paper 56, 1998. |
| [33] | 张新时. 植被的PE(可能蒸散)指标与植被-气候分类(二)——几种主要方法与PEP程序介绍. 植物生态学与地植物学学报, 1989, 13(3): 197–207. |
| [34] | 陈吉龙. 辽河三角洲河口湿地生态系统净初级生产力特征及植被固碳潜力[D]. 北京: 中国科学院大学, 2013. http://www.irgrid.ac.cn/handle/1471x/144663/handle/1471x/54665/browse?type=title&sort_by=1&order=ASC&year=&rpp=20&offset=1163 |
| [35] | Friedlingstein P, Joel G, Field C B, Fung I Y. Toward an allocation scheme for global terrestrial carbon models. Global Change Biology, 1999, 5(7): 755–770. DOI:10.1046/j.1365-2486.1999.00269.x |
| [36] | 朱龙海, 吴建政, 胡日军, 徐永臣, 王楠. 近20年辽河三角洲地貌演化. 地理学报, 2009, 64(3): 357–367. DOI:10.11821/xb200903011 |
| [37] | 王宪礼, 胡远满, 布仁仓. 辽河三角洲湿地的景观变化分析. 地理科学, 1996, 16(3): 260–265. |
| [38] | 鲍永恩, 黄水光. 辽河口海口沉积特征及潮滩动态预测. 沉积学报, 1993, 11(2): 105–112. |
| [39] | 栾维新, 崔红艳. 基于GIS的辽河三角洲潜在海平面上升淹没损失评估. 地理研究, 2004, 3(6): 805–814. |
| [40] | 肖笃宁, 韩慕康, 李晓文, 刘岳峰. 环渤海海平面上升与三角洲湿地保护. 第四纪研究, 2003, 23(3): 237–246. |
| [41] | 涂钢, 李巧萍, 苏丽欣, 袁福香. IPCC SRE A2情景下吉林省气候未来趋势预估及对粮食产量的可能影响. 吉林大学学报:地球科学版, 2010, 40(Z1): 146–153. |
| [42] | 张建平, 赵艳霞, 王春乙, 王靖. 不同气候情景下中国东北与华北作物产量变化趋势模拟. 干旱地区农业研究, 2012, 30(3): 262-266, 278-278. |
| [43] | 刘景利, 纪仰慧, 米娜, 胡丹, 史奎桥, 周丽娜, 李石, 蔡福. B2气候情景下2011-2050年辽宁农业气候资源演变特征. 气象与环境学报, 2012, 28(6): 81–87. |
| [44] | 季劲钧, 黄玫, 李克让. 21世纪中国陆地生态系统与大气碳交换的预测研究. 中国科学D辑:地球科学, 2008, 38(2): 211–223. |
| [45] | 贾建伟, 王磊, 唐玉姝, 李艳丽, 张文佺, 王红丽, 付小花, 乐毅全. 九段沙不同演替阶段湿地土壤微生物呼吸的差异性及其影响因素. 生态学报, 2010, 30(17): 4529–4538. |
| [46] | Twilley R R. Coastal Wetlands & Global Climate Change:Gulf Coast Wetland Sustainability in a Changing Climate. Pew Center on Global Climate, Arlington, 2007. |
| [47] | 郑建平, 王芳, 华祖林. 辽东湾北部河口区生态环境问题及对策. 东北水利水电, 2005, 23(10): 47–50. DOI:10.3969/j.issn.1002-0624.2005.10.022 |