 2017, Vol. 37
2017, Vol. 37文章信息
- 张力, 闫文德, 郑威, 刘益君, 梁小翠, 高超, 方晰
- ZHANG Li, YAN Wende, ZHENG Wei, LIU Yijun, LIANG Xiaocui, GAO Chao, FANG Xi.
- 基于GA-BP人工神经网络的樟树林土壤呼吸对施氮响应的研究
- Responses of Camphor forest soil respiration to nitrogen addition determined based on the GA-BP network
- 生态学报. 2017, 37(16): 5391-5401
- Acta Ecologica Sinica. 2017, 37(16): 5391-5401
- http://dx.doi.org/10.5846/stxb201605230990
-
文章历史
- 收稿日期: 2016-05-23
- 网络出版日期: 2017-03-27





2. 南方林业生态应用技术国家工程实验室, 长沙 410004;
3. 广西壮族自治区林业科学研究院, 南宁 530002;
4. 城市森林生态湖南省重点实验室, 长沙 410004
2. National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China, Changsha 410004, China;
3. Guangxi ForestryRresearch Institute, Nanning 530002, China;
4. Key Laboratory of Urban Forest Ecology of Hunan Province, Changsha 410004, China
土壤呼吸作为全球碳循环主要的组成部分之一, 是土壤碳素向大气输出的主要途径[1], 其总体估值为68—77×1015 g C/a[2-4]。所以土壤呼吸微小变化会明显的影响大气CO2含量乃至全球碳循环[5]。土壤呼吸过程与很多因素相关, 其中土壤呼吸对不同氮素水平下的响应自20世纪80年代以来被越来越多的研究者所重视, 中国作为世界范围内氮沉降最高的地区之一, 对土壤呼吸特征及其控制因子的研究极为重要[5]。
目前已开展很多施氮对土壤呼吸影响研究, 但研究成果大多基于实验观测结果, 受实验地自然条件的限制, 不能研究在一定条件范围内土壤呼吸对施氮响应的完整的连续变化。且由于地下生态过程自身的复杂性, 土壤温湿度、施氮量等因素对土壤呼吸影响, 以及各因素间的协同关系仍存在极大的不确定性, 这就使得土壤呼吸需要一个连续、完整且能反映土壤呼吸各因素间交互作用的模型[6]。
人工神经网络是利用计算机模拟人类神经元工作的程序, 近年来成为解决某些传统方法所无法分析问题的有力工具。人工神经网络可以通过训练带正确结果的样本自动获得“合理的”求解规则, 使其适合解决复杂多因素、非线性问题。在训练完成后, 输入待解决的样本又可达到推广预测的目的[7]。其中GA-BP人工神经网络是一种使用遗传算法优化的BP人工神经网络, 其输出结果较传统的BP人工神经网络稳定性好, 已在多个科学研究领域得到应用。目前, 已有研究者验证了人工神经网络方法模拟CO2通量的可行性[8-9]。同时, 也有在森林、草地和农田生态系统CO2研究中采用能量通量和温度来构建的人工神经网络模型[10]。在我国, 有研究者利用气象环境因子和碳通量数据, 建立了人工神经网络模型, 探讨对碳通量影响很大但并不冗余的环境要素, 模拟和预测森林生态系统与大气间的碳通量[11]。基于人工神经网络能解决复杂问题的特性, 以及模拟土壤呼吸连续、完整变化的需求, 本研究将使用GA-BP人工神经网络研究樟树林土壤呼吸对施氮响应及其影响因素。
1 材料与方法 1.1 研究地概况试验地设于湖南省植物园(113°01′—113°02′ E, 28°06′—28°07′ N), 海拔46—114 m, 坡度为5°—25°。年平均气温17.2℃;1月平均温度最低, 为4.7℃;7月平均温度最高, 为29.4℃;无霜期为270—300 d, 日照时数年均1677.1 h, 年平均降雨量1422 mm。属典型的亚热带湿润季风气候。
研究区属典型红壤丘陵区, 其地层主要是第四纪更新世的冲积性网纹红土和砂砾, 土壤为典型红壤, pH为4.01±0.17。当地无机氮湿沉降量为3.14 g m-2 a-1, NH4-N/NO3--N琋约为6:4。
本研究选择亚热带樟树人工林作为研究对象, 造林时间为1979—1982年, 成林后自然更新, 林龄为10—30 a, 林下植被有白栎(Quercus fabri)、毛泡桐(Paulowwnia tomaentosa)、苦槠(CastanoPsis scleroPhylla)、山矾(SymPlocos caudata), 糙叶树(APhananthe asPera)和柘树(Cudrania tricusPidata)等灌木和小乔木, 草本植物以淡竹叶(LoPhatherum gracile)、酢浆草(Oxalis. Comiculata)、鸡矢藤(Paederia scandens)和商陆(Phytolacca acinosa)为主, 林分特征见表 1。
| 树龄 Age/a |
密度 Stem density/ (株/hm2) |
胸径 DBH/cm |
树高 Height/m |
郁闭度 Crown density |
有机碳 Organic carbon/ (mg/g) |
全氮 Total nitrogen/ (mg/g) |
容重 Bulk density/ (g/cm3) |
pH |
| 20 | 1600 | 14.92 (1.21) |
12.55 (1.03) |
0.9 | 13.18 (0.51) |
1.27 (0.0.03) |
1.50 (0.11) |
4.00 (0.03) |
| 括号内为标准误差 | ||||||||
2010年4月, 在样地内设立12块25 m×25 m的正方形固定样地。2010年5月份, 在样地进行首次施氮处理。根据当地的森林的大气氮沉降背景和其他同类研究, 采用NH4NO3进行施氮, 共设置对照(C, no N added), 低氮(L, 5 g N m-2 a-1), 中氮(M, 15 g N m-2 a-1), 高氮(H, 30 g N m-2 a-1)共4种处理水平, 每一水平设置3个重复, 每个重复样方大小为25 m×25 m。将每个样方需要的采用NH4NO3混合20 L自来水, 采用喷雾器均匀喷洒至样地, 对照样地仅施加20 L自来水。在每年干湿交替季的5月、10月份两次等量施加[12], 本研究期内共施氮5次。在每个重复样方随机选择3个测定点安放PVC连接环(Φ21 cm×H8 cm), 环插入土壤4 cm左右, 测量期间环不再移动。
1.2.2 凋落物收集在每个施氮水平的重复样方内随机设置2个凋落物收集器, 共2×12=24个, 收集器尺寸为3 m×4 m, 收集器采用尼龙网, 离地高度约40 cm。
1.3 测定方法 1.3.1 土壤呼吸测定为了提高模型模拟的准确度, 本研究将2010年作为样地的缓冲期, 以减小施氮对土壤呼吸产生的临时效应。2011年1月至2012年6月上旬, 半月1次测定土壤呼吸。土壤呼吸采用LI-8100(Li-COR Inc., Lincoln, NE, USA)测定;土壤5 cm深度处温度、湿度测定与土壤呼吸速率测定同步进行, 采用LI-8100通量系统配套的温、湿度探针LI-8100—201 Omega、LI-8100—202测定。
1.3.2 凋落物量测定进行土壤呼吸测量时同步测量凋落物鲜重。凋落物量数据用于修正凋落物量对土壤呼吸产生的影响。
1.3.2 GA-BP人工神经网络建模(1) 样本选择
样本取2011年1月至2012年6月上旬间测得土壤呼吸结果作为样本的输出数据, 施氮水平以及同步测量的凋落物量, 土壤温度, 土壤湿度作为输入数据。为了保证数据的稳定性和代表性, 防止模型接受偏离度较大的数据影响, 在输入模型之前将同一块样方重复测量的数据取平均值作为模型输入值, 除去因仪器原因产生错误的5组数据, 最终得到415组数据。为避免人工神经网络发生过拟合, 需要将训练样本和验证样本分别独立。训练样本用于模型的建立、训练, 验证样本通过输入非训练样本集数据检验模型泛用性。随机抽取370组数据作为训练样本, 剩余45组数据作为模型训练样本。同时, 为了防止数据数量级对模型训练带来的影响, 模拟前对输入及输出数据进行归一化处理。归一化公式为:
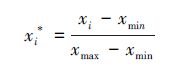
|
式中, xi*为归一化后的数据, xi为归一化前的数据, xmin为样本最小值, xmax为样本最大值
(2) 网络结构和参数的确定
本研究采用最为普及的单隐藏层结构的BP神经网络, 以求得网络输入及输出的一种函数关系。隐含层的节点数(Hidden sizes), 即隐含层的神经元数, 该参数直接影响人工神经网络的训练和模拟效果。当隐含层节点过少时, 训练数据会由于迭代次数过少而影响模拟效果, 隐含层节点过多时, 数据会由于过拟合影响模型的泛用性。
(3) 网络训练与评价
研究添加了一种可逐一计算各隐含层节点情况下数训练样本的MSE, 并挑选出MSE最低时的隐含层节点数的功能。隐含层输入和输出权重即各个输入数据变化对输出数据的影响程度, 通常在GA-BP人工神经网络开始训练时该值为随机取值开始训练。GA-BP即使用遗传算法优化该值已达到更好的模拟效果, 并将优化好的权重值重新赋予GA-BP人工神经网络模型训练。在人工神经网络中, MSE是网络的性能函数, 网络的均方误差, 即"Mean Square Error"。Observed为观测值, predicted为模拟值, 则全体的均方误差为:
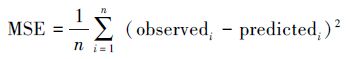
|
训练结果显示, 网络最佳隐含层节点数为17层, 通过拟合训练样本的观测值和模拟值, 两者MSE为0.47, 相关系数R=0.86, 决定系数R2=0.74(图 1)。

|
| 图 1 训练样本模拟值和观测值的回归分析 Fig. 1 Regression analysis of simulated and observed values of training samples |
利用Matlab R2015b建立人工神经网络模型和绘制响应曲面图, 利用Excel 2016和SPSS 24.0对模拟数值进行分析。差异性检验中, 极值分析章节里温度分析部分方差齐性检验数据具有齐性, 采用LSD方法检验数据差异性, 其余部分方差齐性检验结果数据都不具有齐性, 采用Dunnett′s T3方法检验差异性。
2 结果与分析 2.1 土壤呼吸动态樟树林土壤呼吸受太阳辐射、降水、植被类型、土壤性质等因素控制表现出明显的季节变化。土壤呼吸最大值出现6—8月, 最小值出现在1—3月, 施氮对土壤呼吸变化幅度产生了影响, 但未改变土壤呼吸的季节动态(图 2)。

|
| 图 2 土壤呼吸的季节动态 Fig. 2 Seasonal pattern of soil respiration |
模型验证结果表明, 虽有个别值偏差较大, 但观测值和模拟值总体趋势较为吻合(图 3), 总值较为接近, 两者的值分别为111.28和111.94, 差值为-0.34。通过对观测值和模拟值回归分析显示, 验证样本观测值和模拟值MSE为0.65, 相关系数R=0.83, 决定系数R2=0.69(图 4)。模型验证的结果表明, 模型在个别值上会产生较大误差, 但整体上能较好的模拟土壤呼吸的趋势和程度, 该模型能解释土壤呼吸的0.69—0.74。验证样本MSE值虽然较小, 但仍大于训练样本, 说明该模型在一定程度上存在着过拟合的情况。

|
| 图 3 验证样本模拟值和观测值误差比较 Fig. 3 Comparison of simulated and observed values x轴为验证样本的样本序列 |

|
| 图 4 验证样本模拟值和观测值的回归分析 Fig. 4 Regression analysis of simulated and observed values |
本研究在模型输入时设置凋落物水平为50 g/m2, 该水平约为样地凋落物量的年平均值。模拟结果显示各处理下(C、L、M、H), 土壤呼吸速率平均值分别为2.64 μmol m-2 s-1, 2.07 μmol m-2 s-1, 2.01 μmol m-2 s-1, 2.27 μmol m-2 s-1。土壤呼吸平均速率从大至小依次C>H>L>M。不同处理下C与L、M、H, L与H, M与H之间差异显著(P < 0.05), L与M差异不显著(P > 0.05)。施氮在总体上抑制了土壤呼吸速率, 但在H处理下土壤呼吸速率较L与M处理出现了增加, 但未达到C处理的水平。
2.3.2 土壤呼吸极值分析本研究取模拟结果最高和最低的20组数据(约前5%)分析各处理下极大值与极小值。结合图 5与图 6分析, 较低的土壤呼吸速率主要出现在土壤温度降较低区域, 其中在土壤湿度都较低区域的最低。而较高的土壤呼吸速率主要出现在土壤温湿度较高的区域。

|
| 图 5 不同处理下土壤呼吸极大值与极小值 Fig. 5 The value of soil respiration in different treatments |

|
| 图 6 不同处理下土壤呼吸极大值与极小值的土壤温度和土壤湿度 Fig. 6 Max and min values of soil temperature and humidity in soil respiration under different treatments |
施氮下, 土壤呼吸极大值显著减小且随着施氮量增加显著降低(P < 0.05)。除L处理和M处理间不明显外(P > 0.05), 土壤呼吸极小值则受施氮影响显著增加(P < 0.05)。若以土壤温度和土壤湿度为坐标描述土壤呼吸极值, 则各处理下土壤呼吸极大值坐标可表示为C(23.98, 0.336), L(23.2, 0.338), M(21.84, 0.339), H(17.04, 0.338), 极小值可表示为C(8.38, 0.124), L(8.36, 0.154), M(8.34, 0.324), H(8.18, 0.233)。从图 5可以看出, 土壤呼吸在极大值时的土壤温度受施氮的影响发生了显著变化(P < 0.05), 而土壤湿度变化不显著(P > 0.05)。相反的, 土壤呼吸极小值时的土壤温度受施氮的影响不显著(P > 0.05), 而土壤湿度变化较显著(P < 0.05)。
2.4 不同温湿度条件下土壤呼吸对施氮的响应本研究将土壤温度作为x轴, 范围为8.04—25.58℃(为了减小模型边际效应, 模拟时土壤温、湿度输入范围按样本分布缩小10%), 分度为0.4℃, 将土壤湿度作为y轴, 范围为0.11—0.34, 分度为0.004, 以此得到一个含有44×47个数据的矩阵, 将此矩阵每一个数据输入模型模拟, 并将得到的模拟值作为z轴返回到图形, 最终得到一个土壤呼吸关于土壤湿度和土壤温度的响应曲面图及对应的等高线图(图 7)。并将C处理与其他处理下的土壤呼吸相减, 并以同样方法绘制施氮对土壤呼吸变动率的响应曲面图及其对应的等值线图(图 8)。

|
| 图 7 不同处理下土壤呼吸与土壤温湿度的响应曲面图及其对应的等值线图 Fig. 7 Response surface maps of soil respiration and soil temperature and humidity under different treatments 红色“+”为观测值 |

|
| 图 8 对照处理与各施氮量处理下土壤呼吸变动率响应曲面图及其对应的等值线图 Fig. 8 Response surface maps of soil respiration rate under different nitrogen treatments and the corresponding contour maps |
从图 7可知, 不同土壤温湿度条件下土壤呼吸对施氮响应是不同的, 结合图 8可更明显的看出这该响应在不同土壤温度下区分度不大, 但在不同土壤湿度下截然不同。土壤湿度较高范围内, 土壤呼吸受抑制的现象较为明显, 这主要表现在随着施氮量的增加, 土壤呼吸极大值的受抑制程度增加以及总体变动率呈现负增长状态。在土壤湿度较低的范围内, 土壤呼吸速率无论从均值还是极小值看, 均有明显提高, 其变动率也随着施氮量增加而增加。
根据这个现象, 本研究将土壤湿度大于0.25的高湿度区域和土壤湿度小于0.2的低湿度区域分别进行了模拟。高湿度区域模拟结果显示, 各处理下(C、L、M、H), 土壤呼吸速率平均值分别为3.40, 2.44, 2.18, 2.20 μmol m-2 s-1。土壤呼吸平均速率从大至小依次C>L>H>M。除M处理和H处理差异不显著(P > 0.05) 外, 其余处理之间差异均显著(P < 0.05)。这说明在土壤湿度高的区域施氮对土壤呼吸主要呈现抑制作用, 这一点在土壤温度和湿度都很高的区域尤为明显, 但在土壤湿度高、温度低的区域表现相反。
低湿度区域模拟结果显示, 各处理下(C、L、M、H), 土壤呼吸速率平均值分别为1.87, 1.74, 1.92, 2.36 μmol m-2 s-1, 如图 9所示。土壤呼吸平均速率从大至小依次H>M>C>L。除C处理和L处理差异不显著(P > 0.05) 外, 其余处理之间差异均显著(P < 0.05)。这说明在土壤湿度较低的区域, 施氮主要提高了土壤呼吸的速率, 此现象主要分布在土壤湿度和温度都较低的区域, 且随着施氮量的增加, 此现象也随之变得明显。

|
| 图 9 土壤呼吸总平均速率和不同土壤湿度条件下土壤呼吸平均速率图 Fig. 9 Average soil respiration rate and average soil respiration rate under different soil moisture conditions |
总体上看来, 随着施氮量的增加不同湿度范围内土壤呼吸的差异性随之减小, 土壤呼吸总速率、低土壤湿度范围的土壤呼吸和高土壤湿度范围的土壤呼吸, 三者比例从C处理的1:0.83:1.29, 到L处理的1:0.84:1.18, 再至M处理的1:0.95:1.08, 最后到H处理的1:1.04:0.96。结合图 9可以看出随着施氮量的增加, 不同湿度范围内土壤呼吸有向均值趋近的现象。
3 讨论和结论 3.1 土壤呼吸对施氮的不同响应施氮总体上抑制了土壤呼吸, 但这是施氮对土壤呼吸抑制和促进两种作用交互的结果, 其程度主要是由施氮量、土壤温度和土壤湿度决定的, 使得在不同土壤温湿度条件下, 土壤呼吸总体上对施氮呈现显著上升、显著下降和无显著变化3种响应状态。
与本研究对土壤呼吸总平均值的模拟结果相似, 施氮抑制土壤呼吸的结果在全球范围的施氮试验中, 特别是对森林生态系统的实验占据多数[13]。北美温带红松林施氮1年后, 土壤呼吸降低21%—25%, 巴拿马的热带雨林施氮1a后土壤呼吸也出现14%的下降[5], Cusack[14]、Burton[15]等也有相同的发现。
从我国热带、亚热带的相关研究来看, 吴迪等[16]、Mo等[17]、李仁洪等[18]的研究结论也为施氮抑制土壤呼吸, 也本研究结论相似。但以上研究均认为, 随着施氮量的增加, 土壤呼吸速率受抑制程度随之加强, 这与本研究土壤呼吸总速率的变化趋势稍有不同, 但与高土壤湿度下模拟结果一致。其原因为吴迪等[16]、Li R H等[18]的研究属热带或亚热带季风性湿润气候, 温度和降水的分布为夏季高温多雨, 冬季温和湿润, 因此这些研究的结果与本研究在高土壤湿度下的模拟结果表现一致。
另一方面, 本研究在低土壤湿度的模拟结果与张芳等[19]、姜继韶等[20]研究结果相似, 都表现为施氮促进了土壤呼吸。这些研究成果多处于干旱或半湿润气候, 与本研究在该前提下的模拟结果一致, 但对该区域的研究对象多为农作物, 对森林生态系统的研究成果较少, 所以此结论有待进一步证实。
此外, 一些研究成果也显示施氮对土壤呼吸的作用不显著, 如李娇等[21]研究施氮对青藏高原东缘窄叶鲜卑花灌丛土壤呼吸的影响时发现, 施氮对生长季土壤总呼吸和异养呼吸无显著影响, 而对自养呼吸有显著的抑制作用。从本研究对土壤呼吸变动率图可以观察出, 施氮对土壤呼吸不同作用之间有一条较狭长的土壤呼吸变动率较低的区域, 其表现出的土壤温、湿度变化趋势与李娇等研究样地情况有相似之处。
本研究认为, 造成土壤呼吸对施氮的不同响应的主要原因可能是土壤温湿度条件不同。在土壤呼吸速率主要受土壤温湿度限制的条件下, 其整体处于一个较低水平, 在这个水平下微生物活性和根系活性都不高, 间接减弱了施氮对土壤呼吸的抑制作用, 从而使得施氮对土壤呼吸的促进作用变得明显。相反, 在土壤温度和适度相对较适宜的范围, 如温湿度都较高的区域, 施氮对土壤呼吸的抑制作用就显得十分突出。
施氮对植被细根和土壤微生物产生不同的影响是造成土壤呼吸变化的主要原因。一方面施氮减少微生物的量, 从而直接导致了土壤呼吸的降低, 氮添加还会改变分解者食物结构, 造成氮依赖性强、分解效率高的群落占据竞争优势, 造成土壤呼吸的降低[23]。此外植物的细根生物量也会受到施氮的抑制[18], 原因可能为高水平氮添加会降低植物体投入碳至营养吸收系统(地下根系)的需求[23], 直接或间接引起土壤自养、异养呼吸降低[24]。另一方面, 在植物凋落物量固定的情况下, 施氮促进了根系分泌物输入, 提高了土壤有机碳含量, 进而通过影响土壤微生物来影响土壤呼吸速率。
3.2 GA-BP人工神经网络对土壤呼吸的模拟目前对土壤呼吸的模拟主要有两类, 一类是建立土壤呼吸及其影响因素的多元回归模型。这类模型有大量的研究基础, 在某些条件下能取得极好的模拟效果[25], 但这类模型通常考虑因素较少, 且受观测条件限制极大。第二类则是通过一些模型的局部应用模拟土壤呼吸过程, 这类模型由于不是土壤呼吸专用模型, 通常需要大量相关数据才能满足模型运行要求, 但一些和土壤呼吸相关程度较高的因素不能直接输入模型, 或者需要对模型进行二次开发才能输入[26], 这影响了模型模拟的准确率并增加了应用要求和成本。
本研究通过GA-BP人工神经网络建立土壤呼吸对施氮响应的模型, 研究施氮下土壤呼吸在不同土壤温湿度条件下的连续变化, 并将模拟结果采用响应曲面法展现, 得出直观的模拟结果。同时模型的输入数据也可根据实验条件随时调整, 为研究影响土壤呼吸的多因素间关系提供极大的灵活性。但本研究在模拟土壤呼吸对施氮但模型输入数据仍不够全面, 缺少辐射、大气成分、微生物等数据输入, 使得模型对土壤呼吸变化的解释程度不够高。同时, 一些影响土壤呼吸对施氮响应的临时效应也未考虑入模型, 如施氮时对土壤温湿度的改变, 以及脉冲性施氮对土壤呼吸短期的的“激发效应”[27]。这些因素变化相对长期观测虽然持续时间短、影响程度小, 但仍给模型带来了一定的误差。
| [1] | Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7–20. DOI:10.1023/A:1006247623877 |
| [2] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B, 1992, 44(2): 81–99. DOI:10.3402/tellusb.v44i2.15428 |
| [3] | Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles, 1995, 9(1): 23–36. DOI:10.1029/94GB02723 |
| [4] | Schlesinger W H. Carbon balance in terrestrial detritus. Annual Review of Ecology and Systematics, 1977, 8(1): 51–81. DOI:10.1146/annurev.es.08.110177.000411 |
| [5] | Bowden R D, Davidson E, Savage K, Arabia C, Steudler P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecology and Management, 2004, 196(1): 43–56. DOI:10.1016/j.foreco.2004.03.011 |
| [6] | Xu M, Qi Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology, 2001, 7(6): 667–677. DOI:10.1046/j.1354-1013.2001.00435.x |
| [7] | 杨晓帆, 陈廷槐. 人工神经网络固有的优点和缺点. 计算机科学, 1994, 21(2): 23–26. |
| [8] | 何洪林, 于贵瑞, 张雷明, 孙晓敏, 苏文. 基于人工神经网络的ChinaFLUX观测站CO2模拟研究通量. 中国科学D辑:地球科学, 2006, 36(S1): 234–243. |
| [9] | Melesse A M, Hanley R S. Artificial neural network application for multi-ecosystem carbon flux simulation. Ecological Modelling, 2005, 189(3/4): 305–314. |
| [10] | Cleveland C C, Townsend A R. Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(27): 10316–10321. DOI:10.1073/pnas.0600989103 |
| [11] | 温旭丁. ANN模型在亚热带杉木林CO2通量研究中的应用[D]. 长沙: 中南林业科技大学, 2014. http://www.cnki.com.cn/Article/CJFDTOTAL-RMHH200803016.htm |
| [12] | Janssens I A, Dieleman W, Luyssaert S, Subke J A, Reichstein M, Ceulemans R, Ciais P, Dolman A J, Grace J, Matteucci G, Papale D, Piao S L, Schulze E D, Tang J, Law B E. Reduction of forest soil respiration in response to nitrogen deposition. Nature Geoscience, 2010, 3(5): 315–322. DOI:10.1038/ngeo844 |
| [13] | Koehler B, Corre M D, Veldkamp E, Sueta J P. Chronic nitrogen addition causes a reduction in soil carbon dioxide efflux during the high stem-growth period in a tropical montane forest but no response from a tropical lowland forest on a decadal time scale. Biogeosciences, 2009, 6(12): 2973–2983. DOI:10.5194/bg-6-2973-2009 |
| [14] | Cusack D F, Torn M S, McDowell W H, Silver W L. The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Global Change Biology, 2010, 16(9): 2555–2572. |
| [15] | Burton A J, Pregitzer K S, Crawford J N, Zogg G P, Zak D R. Simulated chronic NO3- deposition reduces soil respiration in northern hardwood forests. Global Change Biology, 2004, 10(7): 1080–1091. DOI:10.1111/gcb.2004.10.issue-7 |
| [16] | 吴迪, 张蕊, 高升华, 付晓, 邓红兵, 邵国凡, 张旭东. 模拟氮沉降对长江中下游滩地杨树林土壤呼吸各组分的影响. 生态学报, 2015, 35(3): 717–724. |
| [17] | Mo J M, Zhang W, Zhu W X, Gundersen P, Fang Y T, Li D J, Wang H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Global Change Biology, 2008, 14(2): 403–412. |
| [18] | Li R H, Tu L H, Hu T X, Zhang J, Lu Y, Liu W T, Luo S H, Xiang Y B. Effects of simulated nitrogen deposition on soil respiration in a Neosinocalamus affinis plantation in Rainy Area of West China. Chinese Journal of Applied Ecology, 2010, 21(7): 1649–1655. |
| [19] | 张芳, 郭胜利, 邹俊亮, 李泽, 张彦军. 长期施氮和水热条件对夏闲期土壤呼吸的影响. 环境科学, 2011, 32(11): 3174–3180. |
| [20] | 姜继韶. 施氮和轮作对黄土高原旱塬区土壤温室气体排放的影响[D]. 北京: 中国科学院研究生院(教育部水土保持与生态环境研究中心), 2015. http://cdmd.cnki.com.cn/Article/CDMD-80129-1015996177.htm |
| [21] | 李娇, 尹春英, 周晓波, 魏宇航, 高巧, 刘庆. 施氮对青藏高原东缘窄叶鲜卑花灌丛土壤呼吸的影响. 生态学报, 2014, 34(19): 5558–5569. |
| [22] | Ågren G I, Bosatta E, Magill A H. Combining theory and experiment to understand effects of inorganic nitrogen on litter decomposition. Oecologia, 2001, 128(1): 94–98. DOI:10.1007/s004420100646 |
| [23] | Litton C M, Raich J W, Ryan M G. Carbon allocation in forest ecosystems. Global Change Biology, 2007, 13(10): 2089–2109. DOI:10.1111/gcb.2007.13.issue-10 |
| [24] | Phillips R P, Fahey T J. Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils. New Phytologist, 2007, 176(3): 655–664. DOI:10.1111/nph.2007.176.issue-3 |
| [25] | 周正朝, 上官周平. 黄土高原人工刺槐林土壤呼吸及其与土壤因子的关系. 生态环境学报, 2009, 18(1): 280–285. |
| [26] | 国庆喜, 张海燕, 王兴昌, 王传宽. 东北典型森林土壤呼吸的模拟——IBIS模型的局域化应用. 生态学报, 2010, 30(9): 2295–2303. |
| [27] | 李化山, 汪金松, 刘星, 蒋思思, 张春雨, 赵秀海. 模拟氮沉降对太岳山油松林土壤呼吸的影响及其持续效应. 环境科学学报, 2014, 34(1): 238–249. |