 2017, Vol. 37
2017, Vol. 37文章信息
- 张凯龙, 刘宇航, 童浩杰, 余晓霞, 金园庭
- ZHANG Kailong, LIU Yuhang, TONG Haojie, YU Xiaoxia, JIN Yuanting.
- 贵南沙蜥洞穴空间分布与昆虫资源分布特征的关系
- Correlation between the spatial distribution of active cave and insect resources in Phrynocephalus guinanensis (Lacertilia:Agamidae)
- 生态学报. 2017, 37(16): 5550-5555
- Acta Ecologica Sinica. 2017, 37(16): 5550-5555
- http://dx.doi.org/10.5846/stxb201605180958
-
文章历史
- 收稿日期: 2016-05-18
- 网络出版日期: 2017-03-27

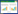




食物是动物生存所需的能量与物质的来源, 是动物与环境交流的枢纽, 也是决定爬行动物群落结构的主要生态维度之一[1], 食物的多样性是动物生存与繁殖不可或缺的重要条件与基本保障[2-3]。研究比较不同栖息地食性资源的多样性有助于了解该物种所占有的营养生态位水平以及种群分布特征的影响因素。对不同生境动物种群数量、分布与食物资源关系的研究有助于人们综合了解动物个体生态需求、种群动态及其受到的相关生态选择压力。
近年来, 已有一些与蜥蜴类食性相关的研究, 如对中国石龙子[4]、变色树蜥[5]、胎生蜥蜴[3]、草原沙蜥[6]、密点麻蜥[7]的研究表明种内两性之间的营养生态位重叠度较高, 而不同发育阶段个体的营养生态位重叠较低, 这被认为有利于幼体蜥蜴生存。对其它一些蜥蜴物种(如Liolaemus pseudoanomalus)的研究表明当食物资源丰富的时候成体与幼体食物组成又趋相近[8]。Zhao和Liu研究表明荒漠沙蜥雌、雄两性食性差异小而不能用来解释其成体两性异形[9], 此结果与对健肢蜥属一物种(Cnemidophorus littoralis)的研究相似[10]。也有对邻近地区或相似环境下的不同种类蜥蜴间的食性进行对比, 研究其种间营养生态位的重叠与分化程度[11-14], 还有对不同海拔以及不同环境分布下的不同种蜥蜴的食性差异进行对比研究其食性群体变异[15-16]。
贵南沙蜥(Phrynocephalus guinanensis)隶属有鳞目(Squamata)、鬣蜥科(Agamidae)、沙蜥属(Phrynocephalus), 是新近命名的一种蜥蜴[17], 在系统分类上被认为是贵德沙蜥(P. putjatia)[18], 但是贵南沙蜥具有明显的两性异形的体色表型, 成年的两性个体在背部颜色、尾侧面颜色、前肢侧面以及外腹部颜色都存在差异, 而贵德沙蜥没有这些颜色表型特征, 并且两种沙蜥成体背部体色深浅差异显著[17], 所以贵南沙蜥可以认为是贵德沙蜥的生态种。贵南沙蜥生活在青海省贵南县的一片植被非常稀疏的孤立沙丘。本文以贵南沙蜥为研究对象, 旨在研究其不同生境的活动洞穴的空间分布及其与可利用昆虫食物资源之间的关系。
1 材料与方法 1.1 调查方法2015年8月中旬, 贵南沙蜥活动季节, 在青海省贵南县有贵南沙蜥分布的区域, 寻找一个草地与沙地接触带(101.04E, 35.79N), 生境如图 1所示, 在该地带设计3条与草地沙地分界线垂直的样线(图 2), 分别标记为A、B、C, 每条样线相隔20 m, 样线的总长度为60 m, 覆盖了局部环境下贵南沙蜥的所有洞穴, 将草地和沙地样线各分成5等份, 即每隔6 m做一个正方形样方, 以其中心且加上1 m2内的4个角做5个陷阱, 陷阱采用装有4%的甲醛水溶液的直径为5.5 cm的170 mL塑料杯, 并且在杯子里滴加5滴甘油防止挥发, 每隔24 h收一次虫子, 每条样线共收3次, 收完后回收塑料杯防止污染环境, 虫子保存于无水乙醇中并且带回实验室依据相关昆虫以及蛛形纲鉴定文献[19-21]鉴定虫子种类, 并统计数量。在样线划定周缘与内部区域(40 m × 60 m)范围内, 寻找有贵南沙蜥活动痕迹的洞穴并记录其与分界线的距离。由于研究区域范围较小(2400 m2), 所以采用传统目估法测量草地与沙地的植被盖度, 草地植被盖度目估达到了60%, 而沙地则只有3%, 根据曹永翔等对植被盖度的分级标准[22], 本研究区域的草地达到了高覆盖植被等级, 对应优良草地, 而沙地则为极低覆盖植被等级, 对应流动沙地。

|
| 图 1 贵南沙蜥生境景观照 Fig. 1 Habitat of P. guinanensis |

|
| 图 2 样线分布图 Fig. 2 Distribution of line transect |
采用Simpon指数(D)、Shannon-Wiener多样性指数(H)、Pielou均匀性指数(J)进行多样性分析[23-24], 具体计算公式如下:
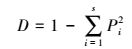
|
(1) |
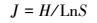
|
(2) |
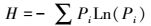
|
(3) |
式中, S代表群落中的总物种数, Pi表示各物种个体数占总个体数的比例(即丰富度), Pi = Ni/N, Ni代表i物种的个体数, N为总个体数, 利用Excel软件对以上指数进行计算。
数据在统计分析前经过正态性与方差同质性检验, 草地与沙地上得到的昆虫个体数与物种数在3条样线之间的统计学差异采用多变量方差分析(MANOVA)进行分析, 洞穴分布与昆虫资源的分布以及两者之间的关系采用SPSS 19.0软件进行Spearman等级相关性分析。
2 结果 2.1 可利用昆虫食物资源组成本次捕捉到的样本鉴定结果如表 1所示, 有42种, 隶属2纲, 10目, 26科, 共1236个, 其中昆虫纲包含半翅目(Hemiptera)、蚤目(Siphonaptera)、膜翅目(Hymenoptera)、鞘翅目(Coleoptera)、双翅目(Diptera)、鳞翅目(Lepidoptera)和毛翅目(Trichoptera)共7目, 21科, 37种, 共978个;蛛型纲包含蜘蛛目(Araneida)、蜱螨目(Acarina)以及盲蛛目(Opiliones)共3目, 5科, 5种, 共258个。半翅目、膜翅目、鞘翅目、双翅目以及蜱螨目是该研究区域的优势类群, 这5个目的种类占全部种类的88.10%, 个体数量占总样本的97.74%。草地样本有37种, 隶属2纲, 9目, 22科, 共570个;沙地样本有33种, 隶属2纲, 9目, 21科, 共666个。
| 分类 Classification |
草地Grassland | 沙地Sand dunes | |||
| 种数 Number of species |
个体数 Number of individuals |
种数 Number of species |
个体数 Number of individuals | ||
| 半翅目Hemiptera | 蚜科Aphididae | 2 | 63 | 2 | 18 |
| 叶蝉科Cicadellidae | 1 | 2 | - | - | |
| 蝽科Pentatomidae | - | - | 1 | 1 | |
| 蚤目Siphonaptera | - | 1 | 2 | - | - |
| 膜翅目Hymenoptera | 茧蜂科Braconidae | 1 | 2 | - | - |
| 细蜂科Serphidae | 1 | 3 | 1 | 3 | |
| 泥蜂科Sphecidae | 1 | 2 | 1 | 1 | |
| 蚁科Formicidae | 4 | 28 | 1 | 1 | |
| 鞘翅目Coleoptera | 未知鞘翅目Unknown Coleoptera | 1 | 1 | 1 | 1 |
| 瓢甲科Coccinellidae | - | - | 1 | 1 | |
| 步甲科Carabidae | 3 | 88 | 2 | 45 | |
| 拟步甲科Tenebrionidae | 1 | 15 | 1 | 332 | |
| 象甲科Curculionidae | 3 | 154 | 4 | 85 | |
| 鳃金龟科Melolonthidae | - | - | 1 | 3 | |
| 双翅目Diptera | 蝇科Muscidae | 5 | 39 | 4 | 26 |
| 麻蝇科Sarcophagidae | 1 | 1 | 1 | 1 | |
| 蚊科Culicidae | 4 | 26 | 4 | 14 | |
| 大蚊总科Tipuloidea | 1 | 12 | 1 | 3 | |
| 食虫虻科Asilidae | 1 | 1 | - | - | |
| 鳞翅目Lepidoptera | - | 1 | 1 | 1 | 1 |
| 毛翅目Trichoptera | 石蛾科Caddisfly | - | - | 1 | 2 |
| 蜱螨目Acarina | 未知蜱螨目1 Unknown acarina1 | 1 | 65 | 1 | 80 |
| 未知蜱螨目2 Unknown acarina2 | 1 | 29 | 1 | 26 | |
| 未知蜱螨目3 Unknown acarina3 | 1 | 22 | 1 | 14 | |
| 盲蛛目Opiliones | 长奇盲蛛科Phalangiidae | 1 | 2 | 1 | 2 |
| 蜘蛛目Araneida | 跳蛛科Salticidae | 1 | 12 | 1 | 6 |
| 总计Total | 37 | 570 | 33 | 666 | |
在草地上的3条样线之间的昆虫物种数(MANOVA, P=0.104) 和个体数量(MANOVA, P=0.260) 差异都不显著, 在沙地上的3条样线之间的昆虫物种数(MANOVA, P=0.367) 和个体数量(MANOVA, P=0.288) 差异不显著, 所以3条样线得到的数据可以合并分析。样线调查区域优势种群为拟步甲科(Tenebrionidae, 28.07%)、象甲科(Curculionidae, 19.34%)以及蜱螨目(19.09%), 其中草地优势种群为象甲科(27.02%)、蜱螨目(20.35%)以及步甲科(Carabidae, 15.44%), 沙地优势种群为拟步甲科(49.85%)和蜱螨目(18.02%)。
研究区域内昆虫Shannon-Wiener多样性指数(H)为2.509, Pielou均匀度指数(J)为0.671, Simpson指数(D)为0.861, 其中草地范围Shannon-Wiener多样性指数(H)为2.719, Pielou均匀度指数(J)为0.753, Simpson指数(D)为0.891;沙地范围的Shannon-Wiener多样性指数(H)为1.947, Pielou均匀度指数(J)为0.557, Simpson指数(D)为0.718, 均比草地低。
2.3 洞穴分布特征与昆虫资源分布特征研究区域共发现贵南沙蜥活动洞穴47个, 其中草地18个, 沙地29个, 空间分布如图 3所示。分析发现草地洞穴分布与离分界线的距离不显著负相关(Spearman相关性r=-0.866, P=0.058), 而沙地洞穴数量与离分界线距离显著负相关(Spearman相关性r=-0.975, P=0.005)。

|
| 图 3 调查样线草地与沙地的贵南沙蜥活动洞穴的空间数量分布 Fig. 3 Spatial distribution of active caves in grassland and sand dunes |
昆虫资源具体分布如表 2所示, 草地上昆虫种类与离分界线的距离无显著相关性(Spearman相关性r=-0.667, P=0.219), 昆虫数量与离分界线的距离也无关(Spearman相关性r=0.100, P=0.873);沙地上昆虫种类与离分界线的距离呈高度负相关(Spearman相关性r=-0.975, P=0.005), 而昆虫数量则与离分界线的距离非显著负相关(Spearman相关性r=-0.500, P=0.391)。
| 离分界线的距离/m Distance from the boundary |
A线line A | B线line B | C线line C | 总计Total | |||||
| 种数 | 个体数 | 种数 | 个体数 | 种数 | 个体数 | 种数 | 个体数 | ||
| 草地Grassland | 0—6 | 9 | 25 | 16 | 37 | 14 | 37 | 25 | 99 |
| 6—12 | 12 | 20 | 15 | 43 | 17 | 35 | 23 | 98 | |
| 12—18 | 12 | 25 | 12 | 41 | 16 | 76 | 24 | 142 | |
| 18—24 | 12 | 30 | 10 | 23 | 12 | 38 | 20 | 91 | |
| 24—30 | 14 | 29 | 11 | 45 | 18 | 66 | 23 | 140 | |
| 沙地Sand dunes | 0—6 | 22 | 37 | 6 | 24 | 10 | 66 | 23 | 127 |
| 6—12 | 12 | 57 | 10 | 30 | 13 | 84 | 21 | 171 | |
| 12—18 | 11 | 86 | 10 | 37 | 10 | 58 | 17 | 181 | |
| 18—24 | 9 | 40 | 5 | 19 | 9 | 22 | 14 | 81 | |
| 24—30 | 8 | 60 | 12 | 46 | 0 | 0 | 14 | 106 | |
以与离分界线的距离每相隔6 m为一个梯度, 计算每个相邻梯度内昆虫洞穴数与昆虫种类与数量, 比较不同梯度洞穴分布与食物资源分布之间的关系, 发现草地上贵南沙蜥洞穴分布与昆虫种类分布不显著相关(Spearman相关性r=0.740, P=0.152), 与昆虫数量也不显著相关(Spearman相关性r=-0.289, P=0.638), 而在沙地上贵南沙蜥洞穴分布与昆虫种类分布呈显著正相关(Spearman相关性r=1.000, P<0.001), 但与昆虫数量分布非显著正相关(Spearman相关性r=0.564, P=0.322)。
3 讨论本文对贵南沙蜥活动季节(8月中旬)栖息地潜在可利用的昆虫食物资源进行研究, 统计发现其共有2纲10目26科42种, 其中以鞘翅目、蜱螨目以及双翅目昆虫个体数量为多。多变量方差分析结果表明草地或沙地上的虫子个体数与物种数在3条样线间无显著差异, 说明研究区域同种生境不同样线数据可以合并分析。分析显示草地昆虫物种多样性指数以及均匀度指数均大于沙地, 表明草地昆虫资源较丰富, 且个体分布较均匀, 草地生态系统可利用的昆虫资源种类较多。
比较草地与沙地的贵南沙蜥洞穴空间分布, 发现沙地上的洞穴远多于草地, 说明沙地的环境更适合贵南沙蜥生存。在草地上洞穴分布均匀, 且其与离分界线的距离无关, 昆虫资源的种类和数量也与离分界线的距离无关, 而在沙地上的贵南沙蜥洞穴明显趋向于分布在离分界线较近的位置, 并且离分界线越近沙地昆虫种类越多。对洞穴分布与昆虫分布之间关系进行相关性分析, 发现草地上洞穴分布与潜在可利用性的昆虫食物资源分布都较均一, 沙地上的昆虫种类与洞穴空间分布不均匀且呈高度的正相关。研究结果表明昆虫资源空间分布特征是促使贵南沙蜥活动洞穴倾向草地分布的重要因素, 而活动洞穴在沙地分布多, 草地分布少, 可能与贵南沙蜥所受到的其它生态因素有关系, 如草原沙蜥(P. frontalis)在不同基底环境下具有不一样的跑速[25], 本文观察到贵南沙蜥在草地跑动较慢, 这会影响其捕食效率与反捕食能力。
综上所述, 草地上洞穴分布较均匀、无规律, 而对应昆虫资源也分布均匀、无规律, 沙地上洞穴与昆虫种类都倾向接近草地分布, 说明贵南沙蜥洞穴分布与食物资源的分布有密切的联系。食物资源分布均匀, 洞穴分布也均匀;食物资源分布不均匀, 洞穴则趋向分布于食物资源较多的地区。贵南沙蜥更多栖息在沙丘, 但是由于草地昆虫资源相比沙地多, 离草地越近越容易捕食, 所以贵南沙蜥的洞穴更趋向于分布在离草地较近的沙地区域, 这也有利于贵南沙蜥的生存。
致谢: 曾炜炜同学协助野外工作,特此致谢。| [1] | Toft C A. Resource partitioning in amphibians and reptiles. Copeia, 1985, 1985(1): 1–21. DOI:10.2307/1444785 |
| [2] | Congdon J D. Proximate and evolutionary constraints on energy relations of reptiles. Physiological Zoology, 1989, 62(2): 356–373. DOI:10.1086/physzool.62.2.30156175 |
| [3] | 赵文阁, 董丙君, 刘鹏, 刘志涛. 胎生蜥蜴的捕食行为及其食性分析. 生态学杂志, 2006, 25(12): 1520–1523. DOI:10.3321/j.issn:1000-4890.2006.12.014 |
| [4] | 林植华, 计翔. 浙江丽水中国石龙子的食性、两性异形和雌性繁殖. 生态学报, 2000, 20(2): 304–310. |
| [5] | 邱清波, 马小梅, 计翔. 海南变色树蜥个体发育中形态和食性的变化. 动物学研究, 2001, 22(5): 367–374. |
| [6] | 赵雪, 毕俊怀, 刘睿, 何志超, 陈绍勇. 草原沙蜥秋季食性分析. 动物学杂志, 2013, 48(3): 321–330. |
| [7] | 刘睿, 毕俊怀, 付明霞, 陈绍勇. 鄂尔多斯地区密点麻蜥的食性分析. 四川动物, 2015, 34(2): 193–199. |
| [8] | Kozykariski M L, Belver L C, Avila L J. Diet of the desert lizard Liolaemus pseudoanomalus (Iguania:Liolaemini) in northern La Rioja Province, Argentina. Journal of Arid Environments, 2011, 75(11): 1237–1239. DOI:10.1016/j.jaridenv.2011.06.004 |
| [9] | Zhao W, Liu N F. Sexual dimorphism of head size in Phrynocephalus przewalskii:testing the food niche divergence hypothesis. Asian Herpetological Research, 2013, 4(4): 242–247. |
| [10] | Menezes V A, Amaral V C, Sluys M V, Rocha C F D. Diet and foraging of the endemic lizard Cnemidophorus littoralis (Squamata, Teiidae) in the restinga de Jurubatiba, Macaé, RJ. Brazilian Journal of Biology, 2006, 66(3): 803–807. DOI:10.1590/S1519-69842006000500005 |
| [11] | Schoener T W. The Anolis lizards of Bimini:resource partitioning in a complex fauna. Ecology, 1968, 49(4): 704–726. DOI:10.2307/1935534 |
| [12] | Schoener T W, Gorman G C. Some niche differences in three lesser Antillean lizards of the genus Anolis. Ecology, 1968, 49(5): 819–830. DOI:10.2307/1936533 |
| [13] | Pianka E R. Niche overlap and diffuse competition. Proceedings of the National Academy of Sciences of the United States of America, 1974, 71(5): 2141–2145. DOI:10.1073/pnas.71.5.2141 |
| [14] | Smith D C. Competitive interactions of the striped plateau lizard (Sceloporus Virgatus) and the tree lizard (Urosaurus Ornatus). Ecology, 1981, 62(3): 679–687. DOI:10.2307/1937736 |
| [15] | 陈小勇, 王建平, 龚大洁. 兰州北山三种蜥蜴春季食性分析. 动物学杂志, 1997, 32(5): 13–18. |
| [16] | 鲍敏, 曾阳, 马建滨, 袁平珍. 青海沙蜥和密点麻蜥的分布状况、生活规律及食性. 青海师范大学学报:自然科学版, 1998(4): 42–45. |
| [17] | Ji X, Wang Y Z, Wang Z. New species of Phrynocephalus (Squamata, Agamidae) from Qinghai, Northwest China. Zootaxa, 2009, 1988: 61–68. |
| [18] | Jin Y T, Yang Z S, Brown R P, Liao P H, Liu N F. Intraspecific lineages of the lizard Phrynocephalus putjatia, from the Qinghai-Tibetan Plateau:Impact of physical events on divergence and discordance between morphology and molecular markers. Molecular Phylogenetics and Evolution, 2014, 71: 288–297. DOI:10.1016/j.ympev.2013.11.004 |
| [19] | 忻介六, 杨庆爽, 胡成业. 昆虫形态分类学. 上海: 复旦大学出版社, 1985. |
| [20] | 李鸿兴, 隋敬之, 周士秀. 昆虫分类检索. 北京: 农业出版社, 1987. |
| [21] | 尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998. |
| [22] | 曹永翔, 刘小丹, 张克斌, 陈明. 青海省都兰县察汗乌苏绿洲植被覆盖度变化研究. 中国沙漠, 2011, 31(5): 1267–1272. |
| [23] | Pielou E C. Ecological Diversity. New York:Wiley Interscience, 1975:16-51. |
| [24] | 赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990. |
| [25] | Li C W, Lian X, Bi J H, Fang H X, Maul T L, Jiang Z G. Effects of sand grain size and morphological traits on running speed of toad-headed lizard Phrynocephalus frontalis. Journal of Arid Environments, 2011, 75(11): 1038–1042. DOI:10.1016/j.jaridenv.2011.06.015 |