 2017, Vol. 37
2017, Vol. 37文章信息
- 王佺珍, 刘倩, 高娅妮, 柳旭
- WANG Quanzhen, LIU Qian, GAO Yani, LIU Xu.
- 植物对盐碱胁迫的响应机制研究进展
- Review on the mechanisms of the response to salinity-alkalinity stress in plants
- 生态学报. 2017, 37(16): 5565-5577
- Acta Ecologica Sinica. 2017, 37(16): 5565-5577
- http://dx.doi.org/10.5846/stxb201605160941
-
文章历史
- 收稿日期: 2016-05-16
- 网络出版日期: 2017-03-25

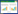




世界上大约有20%的灌溉土壤受到盐度的影响, 且呈不断恶化的趋势[1-2]。预计到2050年, 50%以上的耕地会发生盐碱化[3], 严重威胁着土地利用率和作物产量[4]。中国盐碱地尤其是内陆盐碱地多是盐化和碱化混合, 成分复杂且程度各异, 使人们很容易将盐碱混为一谈, 统称其为盐碱地[5]。实际上, 土壤盐化与碱化分别以盐度、pH值升高为主要特点, 并非两种相同的非生物胁迫[6]。盐化和碱化常常同时发生, 这种现象在很多地区普遍存在。最近数据统计显示, 中国东北盐碱侵害的草地面积已达70%[7], 且仍在扩大。盐、碱对植物的危害程度从大到小依次是盐碱胁迫、碱胁迫、盐胁迫[8-9]。盐碱胁迫会降低土壤渗透势、使离子失衡、打乱生理过程、抑制植物生长、降低作物的质量和产量。严重地区甚至会导致植物死亡。随着科学技术的进步, 盐碱地在技术改良方面已经取得了很多成果, 也是应该继续努力的方向。但是, 还存在着很多地域、资源、成本等限制, 因而, 培育耐盐碱品种的植物, 提高植物的耐盐碱能力是缓解盐碱地对植物影响的一个有效生物措施, 同时还可以产生较好的生态和经济效益, 促进农业的可持续发展。因此, 关于植物适应盐碱逆境的研究已成为国内外专家学者们当前研究中的一个热点。之前关于植物对盐胁迫或碱胁迫的响应及缓解机制的研究很多, 但是对混合盐碱胁迫下所涉及的植物的生理生化反应、基因组学、分子生物学等多角度的响应及缓解机制的综述研究却鲜有报道。因此, 本文综合分析了植物对盐碱响应及缓解机制的不同观点, 提出了提高植物抗盐碱性的有效措施, 最后针对植物适应盐碱逆境方面的研究进行了展望, 以期为提高植物耐盐碱能力、增加作物产量提供科学参考和理论依据。
1 盐碱对植物的伤害类似于其他非生物胁迫, 植物响应盐碱逆境过程中涉及了复杂的生理生化反应[10]。盐碱胁迫会对植物产生很多不利的影响, 包括盐胁迫造成的渗透胁迫、离子毒害、氧化应激等, 除此之外, 还会由于碱胁迫下pH的升高对植物造成进一步的伤害。根系周围土壤pH值升高时, 一些金属离子如Fe2+、Mg2+、Ca2+等沉积, 伴随着无机阴离子减少, 植物对矿质营养的吸收受阻, 造成严重营养胁迫[11-12](图 1), 进而干扰植物的各种代谢活动。这些结论大部分是在短期胁迫下研究得出的, 而Munns等[10-13]证明了在长期盐碱胁迫下, 随着胁迫时间的延长, 植物呈两个生长阶段, 第一阶段是其对短期胁迫的响应(由水或渗透胁迫造成的), 第二阶段是对长期胁迫的调整[14](由植物体内离子毒害作用造成的)。植物在盐碱化土壤上生存, 既要通过渗透调节和离子均衡来躲避渗透胁迫和离子毒害, 又要维持体内的pH平衡。那么盐碱胁迫具体是通过怎样的生理生化过程来影响植物生长的?又可以通过什么样的机制来缓解这种伤害的呢?越来越多的学者开始深入地研究这些问题。

|
| 图 1 盐碱胁迫对植物的伤害 Fig. 1 The harmness of salinity-alkalinity stress to plants |
外部形态和生长状况是反映植物受盐碱伤害程度大小的最直观表现。早期种子萌发时就会受到环境胁迫的影响。Vu等[15]研究发现草木樨(Melilotus officinalis)种子在未发芽阶段可以通过某种机制来抵抗胁迫, 在萌发阶段, 盐度是影响其萌发的主导因素, pH是次要因素。低浓度的盐可以促进种子萌发, 高浓度则会明显抑制其萌发, 且随着盐浓度的升高, 抑制作用增强。类似结果在栓皮栎(Quercus variabilis)[16]、苜蓿(Medicago sativa L.)[17]、红坚木(Dysoxylum spp.)[18]、非洲菊(Gerbera jamesonii Bolus)[19]种子等研究中均有报道, 同时, 渠晓霞等[20]解释了休眠是种子抵抗盐碱胁迫的主要策略。二型性种子在各种盐度下的萌发策略不同。盐胁迫下, 盐地碱蓬(Suaeda sals L.)[21-23]、藜(Chenopodium album L.)[24]的棕色种子比黑色种子离子含量和萌发率更高, 表现出更强的耐盐性。外源ACC与KNO3的协同作用可以显著提高藜黑色种子的萌发率, 低浓度的CaCl2则对棕色种子有更好的缓解作用[24-25]。GA、硫脲和硝酸盐能缓解盐度对荞麦(Halopyrum mucronatum L.)夏季种子的影响, GA、CTK和甜菜碱能够减弱盐度对冬季种子发芽的抑制[26]。此外, 一些物理方法如低温层积、松土、去除种皮通过打破休眠也可以降低盐度对种子的伤害程度[27]。
植物对盐碱胁迫的形态响应主要是通过地上、地下生物量的分配体现的, 且地上部形态和生物量表现更显著。混合盐碱胁迫下, 植物株高、叶片数、茎长及地上部分干物质重等均有所下降, 地下部含水量和幼苗根长下降幅度相对较小, 根茎茎节长度和比根长增加[28], 对于一些耐盐碱植物, 低浓度盐碱胁迫还可以促进其生长, 各耐盐指数呈正态分布, 表现出低促高抑的效应[29], 植株地上部分/地下部分干鲜重显著下降[30]。混合盐碱胁迫主要是通过影响植物对水分的吸收利用[31]、光合作用、酶代谢、气孔因素[32]等来影响其重量的。
2.2 光合作用减弱光合作用是植物物质能量来源的关键代谢过程, 盐碱胁迫下, 叶片细胞结构发生显著变化[33], 光合受阻, 光合速率下降。许多研究从各个角度对此做出了全面解释。Yang等[6]实验证明了随着盐度和pH的增大, 苜蓿叶片中净光合速率、气孔导度和胞间CO2浓度下降。盐碱胁迫中产生的离子毒害和高pH值对植物的光系统II反应中心会造成损伤, 使光合电子传递和PSII的光合作用活力被抑制[34]。同时, 叶绿体结构损伤[35], 参与光反应和卡尔文循环的酶和蛋白下调[36]。而叶片色素的含量还与植物抗盐碱性和盐碱的种类、浓度有关。也有研究者认为, 光合作用产生的干物质主要在叶片中积累, 盐碱胁迫下叶原基的发生受到抑制, 单株植物的光合面积减少, 因而间接造成植物碳同化量减少使得植物生长受到影响[37]。此外, Mg2+是叶绿素合成的必要元素, 而碱胁迫还会导致Mg2+沉淀, 使得叶绿素合成受阻[38], 含量减少, 光合作用减弱。实际上, 凡是涉及到能量供应的碳水化合物代谢、光合作用、TCA循环都会受到盐碱胁迫的抑制, 且碱胁迫的抑制程度大于盐胁迫, 高丰度的单糖和TCA循环中间体的积累提供了更多的能量来源[39]。
2.3 扰乱离子动态平衡离子在植物正常生长过程中起着重要的作用, 土壤盐碱化会打破植物体内的离子的动态平衡[40]。特别是碱胁迫下pH的升高会破坏根系离子的吸收运输。Shaheen[41]等研究发现羊草的根茎和叶片中, Na+含量随着土壤盐碱程度的增大显著增加, 而K+含量则明显减少。在禾本科盐生互米花草(Spartina alterniflora)[42]、甜高粱(Sorghum bicolor)[43]、玉米(Zea maize L.)[44]中也得到了相似的结论。同时, H+-ATP酶活性增强, Na+/ K+交换活动显著上调, 而H+-PPase活性则有所下降。这也说明了H+-ATP酶和Na+/K+的平衡在植物抗盐碱胁迫中发挥着重要作用。根部K+外流引起的K+-Na+不平衡还会使气孔导度变小, 气孔的长度、宽度和气孔孔径宽度均随盐碱浓度的增加而降低[45]。Wang等[9]发现限制向日葵离子运输的主要部位是子叶节区。不同浓度的盐碱胁迫对抗逆性不同的植物影响有所差异, 但高浓度的盐碱胁迫会普遍对离子平衡产生不利影响。
2.4 膜透性增大细胞膜是使细胞维持稳定胞内代谢环境的必要屏障, 其具有的选择透过性可以调节和选择物质进出。在盐碱胁迫中, 植物首先受到伤害的是细胞膜的结构和功能, 引起质膜透性增大。一方面植物体内渗透势升高, 使植物的各种膜系统产生渗透胁迫。另一方面, 由于带电单价离子浓度的升高对细胞产生专性毒害作用, 使一些生物大分子如酶、蛋白质以及膜结构的稳定性被破坏[46]。植物叶片质膜受伤害程度随盐碱浓度的增大而增加, 高盐浓度下, 膜的选择透过性被破坏, 大量电解质外渗。
生物膜的破坏主要是由膜质过氧化造成的, 质膜的破坏程度可以用细胞质膜透性的变化和丙二醛(malondialdehyde, MDA)的含量来反映[47]。具有细胞毒性的膜脂过氧化产物丙二醛, 会引起生物大分子如蛋白质、核酸等的交联聚合。它与质膜透性的变化规律相同[48], 两者显著正相关, Li等[49]发现在轻度和中度盐碱胁迫下, 欧李(Cerasus humilis (Bge.) Sok.)叶片中MDA含量的变化趋势与CK类似, 而在重度胁迫下MDA在叶片中的含量从一开始就急剧增加。这在白刺[50]、水稻(Oryza sativa)[51]中都得到了验证。因此, 细胞膜透性的变化反应了外部不良环境对植物细胞的伤害程度, 同时细胞膜在逆境下的稳定性也反映了植物抗逆性的高低。
3 植物对盐碱胁迫的缓解机制盐碱逆境下, 植物主要通过合成渗透调节物质、提高酶的抗氧化能力、对离子选择性吸收、营养平衡、改变代谢类型、调整生物量的分配等方法来减轻不良环境对其生长发育造成的伤害。其中, 植物体根的代谢调控发挥着重要作用[52-53], 是植物应对盐碱胁迫的第一道屏障, 积累溶质来进行渗透调节是缓解盐碱胁迫的主要途径。
3.1 合成渗透调节物质参与植物抗盐碱过程的渗透调节物质主要包括两大类:(1) 从外界环境进入植物细胞的无机离子, 如K+、NO3-、Cl-和无机酸盐等。(2) 细胞内合成的有机溶质, 如脯氨酸、甜菜碱、胆碱、有机酸等, 还包括一些代谢中间产物如糖类及其衍生物等。这两类渗透调节物质在植物抵抗盐碱胁迫中都发挥着重要作用。
Ranganayakulu等[54]发现高浓度盐、碱胁迫下, 花生(Arachis hypogaea L.)茎叶中甜菜碱大量积累。外源甜菜碱可以明显提高莴苣(Lactuca sativa L.)[55]的生长。甜菜碱不仅参与细胞的渗透调节还在植物气孔运动、稳定生物大分子、呼吸作用等过程中发挥着重要作用。喷施外源甜菜碱, 可以提高作物的渗透调节能力, 减轻逆境对其的伤害[56]。
游离状态的脯氨酸在植物体内广泛分布, 是主要的有机渗透调节物质。在稳定生物大分子结构、解除氨毒及作为能量库调节细胞氧化还原势等方面也起着重要作用。脯氨酸的积累是植物在盐碱胁迫下自身出现的一种防御性行为, 也是其遭受逆境胁迫的一种信号[57]。叶面喷施脯氨酸可以刺激茎叶和根部的生长, 对提高植物干鲜重、光合速率、抗氧化酶活性等都有贡献[58]。
可溶性糖不仅可以为有机物的合成提供物质和能量, 还作为一种渗透调节物质在植物的生长发育起着重要作用, 随着盐碱浓度的增大, 植物可溶性糖含量呈先增多后减少的趋势[59], 耐盐性强的在较高盐碱浓度下可溶性糖含量仍显著增加。在鹰嘴豆生殖生长期输入蔗糖, 可以增加不同组织中的总糖和缓解盐诱导的繁殖障碍, 增强其耐盐性[60]。
有机酸可以清除单态氧和包含OH-的自由基, 在离子平衡和酸碱调节中发挥作用。燕麦幼苗在盐胁迫和碱胁迫下的生理响应机制是不同的, 盐胁迫下以积累Cl-为主, 碱胁迫下以积累有机酸为主[31]。Guo等[61]发现星星草(Puccinellia tenuiflora)在响应碱胁迫中起关键作用的是根际有机酸(主要是柠檬酸)的积累, 同时还发现甜菜碱、胆碱在盐胁迫和碱胁迫下都对渗透调节的影响是微不足道的。这与大多数研究者的结论略有差异, 其中特殊的机理还有待进一步探讨。此外, 在盐碱胁迫下, 羊草中主要积累柠檬酸, 蓼科中主要积累草酸[62-63], 说明不同植物在缓解盐碱对其造成的伤害中有机酸代谢调节不同。随着胁迫时间的延长, 有机酸含量下降[64], 但是尚不明确这是由于其中的盐离子及pH引起的还是合成代谢被抑制导致的。
3.2 提高酶的抗氧化能力植物体内的抗氧化酶包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)等。它们主要通过清除体内的活性氧来保护酶系统, 还参与细胞的光合、呼吸、木质素的形成等, 在叶绿体、线粒体、细胞质中发挥作用。这些酶的活性可以反应植物体内代谢和抗逆性的变化。渗透调节能力越强, 保护酶活性越高的植物, 对盐碱逆境能表现出更好的适应性[65-66]。
受盐碱胁迫的影响, 植物体内ROS积累[67], SOD、POD、CAT活性先增强后降低[30], 而柽柳(Tamarix chinensi)中由于活性氧代谢平衡被打破, APX活性一直呈下降趋势[68], SOD以去除超氧离子为主[69], 将O2-·歧化为H2O2, CAT和POD以清除过氧化物H2O2为主, 将H2O2氧化还原为O2和H2O以减缓其对细胞的毒害[70]。由于植物生长减慢, 这些抗氧化酶合成减少, 活性下降。而乔枫等[71]在沙棘(Hippophae rhamnoides L.)幼苗研究中则发现在高浓度盐碱胁迫下, POD活性基本处于上升趋势, 仍可发挥其清除H2O2的作用, 猜测高浓度盐碱胁迫下, 不同植物的主要抗氧化酶略有差异, POD可能在沙棘抗逆过程中起主要作用。
3.3 对离子的选择性吸收及pH平衡在盐碱胁迫下, Na+的积累会使细胞的膜系统受损, 而K+作为一种重要的无机溶质, 对降低植物细胞渗透势和维持水分平衡至关重要。植物通过限制Na+进入细胞, 并选择性吸收K+来维持组织细胞的高K+/Na+值以保证的正常生理代谢[72]。在植物抗盐碱中, 这种离子选择性吸收和离子平衡比渗透调节物脯氨酸发挥着更大的作用[73]。
植物盐胁迫信号转导途径主要有蛋白激酶途径、ABA途径(依赖ABA途径、不依赖ABA途径)、SOS途径(图 2)。Ca2+作为第二信使与碱胁迫显著正相关, 高浓度的钙会立即触发SOS[74]钠排除系统来减轻钠的伤害[75-76], 还可以诱导合成新的胁迫蛋白。研究发现, 在盐碱胁迫下钙离子含量的增加有利于提高大米[77]、小麦[78]、番茄(Lycopersicon esculentum Mill.)[79]等的耐盐碱能力。因此, 植物可以通过增强对钙的吸收来缓解盐碱胁迫。最新研究还发现, 高盐胁迫下, 胡杨释放的胞内Ca2+和H2O2会介导产生胞外ATP (eATP)信号[69], 该物质不仅可以调节植物的生长和抗氧化防御, 还能间接调节其耐盐性。

Mg2+是叶绿素形成必不可少的元素, 在光合和呼吸作用中的某些酶也需要有Mg2+的激活才能发挥作用, 此外, 它还可以调节气孔关闭, 影响离子平衡。Rubio等[81]报道提高盐碱处理液中Ca2+的含量后, 胡椒(Piper nigrum L.)的蒴果产量增加。盐碱胁迫下植物会吸收这些对自身有利的离子, 排出有害离子。同时, 植物还可以通过提高对微量元素的吸收及分配能力来保持体内营养均衡[82], 更好地适应盐碱胁迫。
在调节植物体pH平衡中, 起主导作用的是有机酸和无机阴离子, 盐胁迫下细胞内有机酸变化幅度较小, 而在碱胁迫下会显著增加来补充体内的阴离子保证细胞pH稳定。
3.4 诱导抗盐碱相关基因表达盐碱胁迫会抑制一些正常基因的表达, 而加强与抗胁迫相关基因的表达。植物的耐盐碱性不是由单个基因决定的, 是多个抗逆基因共同表达调控的。拟南芥中PutAPX超表达提高叶绿素含量、降低膜脂过氧化程度[83];GmST1增加对ABA的敏感性, 减少活性氧的产生[84];AtHD2D[85]增加根冠比、减少MAD含量。不同的基因通过调控不同的代谢途径都可以提高其耐盐碱性, 这些耐盐碱基因主要包括四大类:(1) 合成渗透保护物质的相关基因(2) 离子转运蛋白基因(3) 抗氧化相关基因(4) 信号转导相关基因。PEAMT基因催化产生磷酸胆碱提高辽宁碱蓬抗盐性[86]、McNHX2表达产物NHX逆向转运蛋白作为离子转运体通过调节Na+浓度维持冰花(Mesembryanthemum crystallinum)细胞内离子稳态提高其耐盐性[87]。SlSAMS1在番茄中过表达可促进多胺的积累, 而多胺可以提高H2O2酶的活性, 从而减缓细胞氧化损伤增强其抗盐碱性[8]。烟草中NtABF及RrANR对ROS的清除和ABA信号途径的正调控可使植物免受高盐碱危害[88]。这些不同类型的基因共同协作帮助植物度过盐碱逆境。
随着盐碱胁迫的增加, 总糖含量和可溶性蛋白质也会增加。张建秋等[89]在盐碱胁迫下用双向电泳技术分析白刺中蛋白表达情况时发现, 其叶片中蛋白总量无显著变化, 但对盐碱胁迫有正贡献的特异蛋白表达如66, 28kD蛋白明显增多。近年来, 越来越多的研究发现, 铁蛋白的mRNA和蛋白质也会在逆境胁迫下显著增加, 铁蛋白通过贮藏铁离子来帮助植物抵御氧化胁迫[90]。有报道显示, 转基因烟草(Nicotiana tabacum L.)过表达铁蛋白后, 植株对活性氧的胁迫耐受性显著提高[91]。野大豆(Glycine gracilis )的根系可以依赖特定的转录因子和氧化还原相关基因来响应盐碱胁迫[92], 越来越多的耐盐碱相关蛋白如SGF14c蛋白[93]、GsSKP21蛋白[94]、GmbZIP110[95]等在大豆中被发现。此外, SAM合成酶[96]、MdSOS2L1蛋白激酶[97]等通过多胺代谢也能提高番茄的抗盐碱能力。半胱氨酸蛋白酶抑制剂GsCPI14与钙/钙调素结合的受体样蛋白激酶GsCBRLK相互作用帮助大豆抵御盐碱胁迫[98]。由此可见, 植物通过抗逆基因的选择性表达, 产生一些抗盐碱相关蛋白也是植物抵抗盐胁迫的一种有效方式, 而这些蛋白究竟是通过怎样的机理来帮助植物抵抗盐碱胁迫的还有待进一步研究。
4 提高植物抗盐碱性的途径 4.1 外源物质的加入施加外源物质是缓解盐碱胁迫的一种有效抗盐方式, 现在已经有越来越多对盐碱胁迫下幼苗生长有缓解作用的外源物质被发现。较常用的外源添加物主要包括四大类, 一是渗透调节物质如甜菜碱、糖类[99]、有机酸等;二是与降低膜透性有关的物质如水杨酸、腐殖酸、Ca2+等;三是可以提高植物抗氧化能力的物质如硒、硅[100-101]、NO[102]、γ-氨基丁酸[103]等, 四是植物生长调节剂如茉莉酸[104]、CTK[105]、IAA等, 此外还有一些其他物质如镧[106]、降黑素[107]等也可以提高植物的抗盐碱性。当然, 还有很多外源物质尚未发现, 需要我们深入研究发现更多可以提高植物抗盐碱性的外源物质。张毅等[108-109]研究发现叶面喷施亚精胺(Spd)会明显促进番茄生长, 提高其耐盐碱性, 并证实了这与Spd对光合机构的保护效应和参与氮代谢提高营养平衡密切相关。此外该物质还可以减轻对番茄根系线粒体的伤害[110]。Chunthaburee等[111]补充Spd还可以通过增强花青素, 酚类物质含量和抗氧化能力提高水稻的耐盐碱性。因此, 作为一种低成本的抗盐碱方式, 外源物质缓解逆境胁迫成为研究中的热点。
4.2 与真菌的协同效应内生菌可以通过参与渗透调节[112]、光合作用、提高植物营养[113]、增强抗氧化系统等方面来缓解盐碱胁迫对植物的伤害。Herriel等[114]报道, 在NaCl胁迫下接2种AMF对辣椒(Capsicum annuum)和洋葱(Allium cepa)的生长都有促进作用, 能明显提高这两种作物的抗盐碱性。Zarea等[115]研究发现在小麦上接种内生菌可以通过提高叶绿素含量, 促进光合作用来提高植物的抗盐碱性。用菌根真菌既可以提高植物的抗盐碱性又可以减少化学制剂的使用, 减轻环境压力。在这方面的研究也可以挖掘更多的思路。
4.3 利用生物技术手段转基因技术一直是学术研究中的热点, 其安全性也是一个备受争议的话题。转OsCYP2基因水稻在苗期的抗盐碱能力要高于对照[116]。野生大豆中分离出的基因GsJAZ2[117]、新型混合富含脯氨酸型基因GsEARLI17[118];甜高粱中的SbCIPKs基因[119];苜蓿中的MtWRKY76[120]、GsGSTU13[121]等都对植物耐盐碱胁迫起着正调控作用。用携带小麦TaNHX2基因的根癌农杆菌LBA4404侵染辣椒(Capsicum annuum L.), 发现转基因辣椒的脯氨酸、叶绿素、SOD、APX、相对含水量水平都增强、并减少H2O2、丙二醛含量[122]。此外, 有研究报道相同条件下种植转SOS基因高羊茅会减低土壤pH和盐碱度。但是还没有探明到底是哪种差异引起的。虽然现在在转基因技术方面的研究越来越热, 但是相比于其他方法, 这方面的研究成果相对较少, 还有很大的提升的空间。
4.4 培育耐盐碱品种与生物技术手段相结合, 对植物基因进行修饰改造实现抗盐碱基因的转移;品种间杂交;对现有植物进行耐盐碱筛选等都是培育耐盐碱品种的有效途径。如拟南芥中多胺氧化酶5的功能缺失突变[123]、二倍体野生西方白三叶和普通白三叶回交[124]都可以选育出耐盐碱品种。还要大量培育像沙棘、甜高粱、结缕草等这样本身有较强耐盐碱性的植物。此外, 抗盐碱性差异较大的两种植物还可以通过嫁接[125]提高抗性。
4.5 抗性锻炼抗盐碱锻炼可以提高植物的抗盐碱性。高等植物会有一些“胁迫记忆”或“胁迫印记”, 植物首次受到的胁迫可以通过适应性响应的诱导增强植物的抗逆性[126]。一般植物在幼苗时期对盐碱最为敏感, 播种前用不同梯度的盐碱溶液浸泡种子一段时间, 吸水膨胀后再进行萌发, 可以提高植物的耐盐碱性。拟南芥种子用甲萘醌亚硫酸氢钠(MSB)浸种后发现对种子萌发并没有影响, 但是却会使拟南芥生长更快, 积累更多的脯氨酸, 表现出更高的耐盐碱性[127]。
5 展望在盐碱胁迫方面, 关于植物对盐胁迫的响应机制是研究最多的, 在碱胁迫和混合盐碱胁迫方面的研究的相对较少, 而且很多研究中将盐碱胁迫笼统的称为盐胁迫, 缺乏科学性。盐化与碱化常常同时发生, 存在相互影响的交互作用。因此, 研究混合盐碱胁迫对提高作物产量和环境建设的影响有重要的现实意义。
(1) 找到平衡点, 确定主导影响因子盐碱胁迫虽会降低植物产量, 但也会提高一些植物果实的质量, 如小麦[1]、番茄[128]、燕麦[129]等, 因此, 确定能使植物产量和质量达到最优的盐浓度平衡阈值, 是今后研究的一个新的突破口。目前, 大多数研究多集中在种子萌发过程和植物幼苗时期, 对植物整个生命过程的系统综合研究较少, 从植物生长的不同时期不同部位进行探讨比较各指标的动态变化和具体的信号转导过程或许会有新的发现。此外, 各生理过程受到不同离子、蛋白、次级代谢产物等交互作用的影响, 发现新指标、确定不同时期影响植物不同部位的主导因素以及植物耐盐碱主导因子也是必要的。
(2) 筛选抗性基因, 全面解释机制目前, 植物对盐碱胁迫响应的分子机制研究还处于初级阶段, 筛选耐盐碱相关基因、对基因家族进行转录组测序、系统发育分析、亚细胞定位及利用蛋白质互作和表达谱信息对基因功能深层预测是当前研究的重中之重。同时, 也不能忽视对植物抗性有负调控作用的基因改造研究, 如拟南芥中的AtRNP1[130]。植物抗盐碱性是由多个基因共同调控的一个复杂过程, 从耐盐植物和敏盐植物之间在抗盐碱方面差异的角度开展高通量差异数据分析, 进而找出造成差异的根本原因, 发挥蛋白质和基因组学的优势, 利用cDNA芯片、双向电泳技术、农杆菌介导法等对植物耐盐碱基因进行生物学鉴定, 实现抗盐碱基因的转移, 从植物生理学、基因组学、分子生物学、蛋白组学等不同学科综合系统地挖掘和解释植物的抗盐碱机制对于改良作物具有重要的意义, 也将是今后研究的热门课题。此外, 很多植物中检测到差异表达蛋白中超过一半的蛋白质信息是未知信息, 这些未知信息里必定蕴藏着与抗盐碱相关的有用信息, 这些信息的揭示必将在植物抗盐碱研究中有重大突破。改造并筛选得到越来越多耐盐碱的修复植物必将带来更大的生态和经济效益, 这种环境友好型的生物手段无疑具有很大的潜在优势和发展前景。
| [1] | Zhang X X, Shi Z Q, Tian Y J, Zhou Q, Cai J, Dai T B, Cao W X, Pu H C, Jiang D. Salt stress increases content and size of glutenin macropolymers in wheat grain. Food Chemistry, 2016, 197: 516–521. DOI:10.1016/j.foodchem.2015.11.008 |
| [2] | Zhao X Y, Bian X Y, Li Z X, Wang X W, Yang C J, Liu G F, Jiang J, Kentbayev Y, Kentbayeva B, Yang C P. Genetic stability analysis of introduced Betula pendula, Betula kirghisorum, and Betula pubescens families in saline-alkali soil of northeastern China. Scandinavian Journal of Forest Research, 2014, 29(7): 639–649. DOI:10.1080/02827581.2014.960892 |
| [3] | Wang W X, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures:Towards genetic engineering for stress tolerance. Planta, 2003, 218(1): 1–14. DOI:10.1007/s00425-003-1105-5 |
| [4] | Wang J C, Yao L R, Li B C, Meng Y X, Ma X L, Lai Y, Si E J, Ren P R, Yang K, Shang X W, Wang H J. Comparative proteomic analysis of cultured suspension cells of the halophyte Halogeton glomeratus by iTRAQ provides insights into response mechanisms to salt stress. Frontiers in Plant Science, 2016, 7(30): 1–12. |
| [5] | Shi D C, Yin S J, Yang G H, Zhao K F. Citric acid accumulation in an alkali-tolerant plant Puccinellia tenuiflora under alkaline stress. Acta Botanica Sinica, 2002, 44(5): 537–540. |
| [6] | Yang J Y, Zheng W, Tian Y, Wu Y, Zhou D W. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings. Photosynthetica, 2011, 49(2): 275–284. DOI:10.1007/s11099-011-0037-8 |
| [7] | Shi D C, Wang D L. ffects of various salt-alkali mixed stresses on Aneurolepidium chinense (Trin.) Kitag. Plant Soil, 2005, 271(1/2): 15–26. |
| [8] | Gong B, Wang X F, Wei M, Yang F J, Li Y, Shi Q H. Overexpression of S-adenosylmethionine synthetase 1 enhances tomato callus tolerance to alkali stress through polyamine and hydrogen peroxide cross-linked networks. Plant Cell, Tissue and Organ Culture, 2016, 124(2): 377–391. DOI:10.1007/s11240-015-0901-5 |
| [9] | Wang X P, Jiang P, Ma Y, Geng S J, Wang S C, Shi D C. Physiological strategies of sunflower exposed to salt or alkali stresses:restriction of ion transport in the cotyledon node zone and solute accumulation. Agronomy Journal, 2015, 107(6): 2181–2192. DOI:10.2134/agronj15.0012 |
| [10] | Chen L, Wang R Z. Anatomical and physiological divergences and compensatory effects in two Leymus chinensis (Poaceae) ecotypes in Northeast China. Agriculture, Ecosystems & Environment, 2009, 134(1/2): 46–52. |
| [11] | 郭瑞, 李峰, 周际, 李昊儒, 夏旭, 刘琪. 亚麻响应盐、碱胁迫的生理特征. 植物生态学报, 2016, 40(1): 69–79. DOI:10.17521/cjpe.2015.0240 |
| [12] | Capula-Rodríguez R, Valdez-Aguilara L A, Cartmill D L, Cartmill A D, Alia-Tejacalc I. Supplementary calcium and potassium improve the response of tomato (Solanum lycopersicum L.) to simultaneous alkalinity, salinity, and boron stress. Communications in Soil Science and Plant Analysis, 2016, 47(4): 505–511. |
| [13] | Munns R. Comparative physiology of salt and water stress. Plant, Cell & Environment, 2002, 25(2): 239–250. |
| [14] | Wang L X, Fang C, Wang K. Physiological responses of Leymus chinensis to long-term salt, alkali and mixed salt-alkali stresses. Journal of Plant Nutrition, 2015, 38(4): 526–540. DOI:10.1080/01904167.2014.937874 |
| [15] | Vu T S, Zhang D W, Xiao W H, Chi C Y, Xing Y, Fu D D, Yuan Z N. Mechanisms of combined effects of salt and alkaline stresses on seed germination and seedlings of Melilotus Officinalis (Fabaceae) in Northeast of China. Pakistan Journal of Botany, 2015, 47(5): 1603–1611. |
| [16] | 李志萍, 张文辉, 崔豫川. NaCl和Na2CO3胁迫对栓皮栎种子萌发及幼苗生长的影响. 生态学报, 2015, 35(3): 742–751. |
| [17] | Gao Z W, Zhu H, Gao J C, Yang C W, Mu C S, Wang D L. Germination responses of Alfalfa (Medicago sativa L.) seeds to various salt-alkaline mixed stress. African Journal of Agricultural Research, 2011, 6(16): 3793–3803. |
| [18] | Meloni D A, Gulotta M R, Martinez C A. Salinity tolerance in Schinopsis quebracho colorado:Seed germination, growth, ion relations and metabolic responses. Journal of Arid Environments, 2008, 72(10): 1785–1792. DOI:10.1016/j.jaridenv.2008.05.003 |
| [19] | Tavakkoli M M, Roosta H R, Hamidpour M. Effects of alkali stress and growing media on growth and physiological characteristics of gerbera plants. Journal of Agricultural Science and Technology, 2016, 18(2): 453–466. |
| [20] | 渠晓霞, 黄振英. 盐生植物种子萌发对环境的适应对策. 生态学报, 2005, 25(9): 2389–2398. |
| [21] | 刘艳, 周家超, 张晓东, 李欣, 范海, 王宝山, 宋杰. 盐地碱蓬二型性种子及其幼苗对盐渍环境的适应性. 生态学报, 2013, 33(17): 5162–5168. |
| [22] | 史功伟, 宋杰, 高奔, 杨青, 范海, 王宝山, 赵可夫. 不同生境盐地碱蓬出苗及幼苗抗盐性比较. 生态学报, 2009, 29(1): 138–143. |
| [23] | Li W Q, Liu X J, Khan M A, Yamaguchi S. The effect of plant growth regulators, nitric oxide, nitrate, nitrite and light on the germination of dimorphic seeds of Suaeda salsa under saline conditions. Journal of Plant Research, 2005, 118(3): 207–214. DOI:10.1007/s10265-005-0212-8 |
| [24] | Yao S X, Chen S S, Zhao J, Xu D S, Lan H Y, Zhang F C. Effect of three salts on germination and seedling survival of dimorphic seeds of Chenopodium album. Botanique, 2010, 88(9): 821–828. DOI:10.1139/B10-052 |
| [25] | Faiza S, Bilquess G, Li W Q, Liu X J, Ajmal K M. Effect of calcium and light on the germination of Urochondra setulosa under different salts. Journal of Zhejiang University Science B, 2007, 8(1): 20–26. DOI:10.1631/jzus.2007.B0020 |
| [26] | Khan M A, Ungar I A. Alleviation of salinity stress and the response to temperature in two seed morphs of Halopyrum mucronatum (Poaceae). Australian Journal of Botany, 2001, 49(6): 777–783. DOI:10.1071/BT01014 |
| [27] | Wang L, Huang Z Y, Baskin C C, Baskin J M, Dong M. Germination of dimorphic seeds of the desert annual halophyte Suaeda aralocaspica (Chenopodiaceae), a C4 plant without kranz anatomy. Annals of Botany, 2008, 102(5): 757–769. DOI:10.1093/aob/mcn158 |
| [28] | Liu B S, Kang C L, Wang X, Bao G Z. Physiological and morphological responses of Leymus chinensis to saline-alkali stress. Grassland Science, 2015, 61(4): 217–226. DOI:10.1111/grs.12099 |
| [29] | 慈敦伟, 张智猛, 丁红, 宋文武, 符方平, 康涛, 戴良香. 花生苗期耐盐性评价及耐盐指标筛选. 生态学报, 2015, 35(3): 805–814. |
| [30] | 李晓荣, 姚世响, 陈莎莎, 兰海燕. 藜异型性种子后代植株盐响应生理机制. 生态学报, 2015, 35(24): 8139–8147. |
| [31] | Gao Z W, Han J Y, Mu C S, Lin J X, Li X Y, Lin L D, Sun S N. Effects of saline and alkaline stresses on growth and physiological changes in oat (Avena sativa L.) seedlings. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2014, 42(2): 357–362. |
| [32] | 赵春桥, 李继伟, 范希峰, 侯新村, 武菊英, 胡跃高, 刘吉利. 不同盐胁迫对柳枝稷生物量、品质和光合生理的影响. 生态学报, 2015, 35(19): 6489–6495. |
| [33] | 李学孚, 倪智敏, 吴月燕, 李美芹, 刘蓉, 饶慧云. 盐胁迫对'鄞红'葡萄光合特性及叶片细胞结构的影响. 生态学报, 2015, 35(13): 4436–4444. |
| [34] | Kalaji H M, Jajoo A, Oukarroum A, Brestic M, Zivcak M, Samborska I A, Cetner M D, Lukasik I, Goltsev V, Ladle R J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiologiae Plantarum, 2016, 38: 102. DOI:10.1007/s11738-016-2113-y |
| [35] | 刘卫国, 丁俊祥, 邹杰, 林喆, 唐立松. NaCl对齿肋赤藓叶肉细胞超微结构的影响. 生态学报, 2016, 36(12). DOI:10.5846/stxb201410122011 |
| [36] | Cheng T L, Chen J H, Zhang J B, Shi S Q, Zhou Y W, Lu L, Wang P K, Jiang Z P, Yang J C, Zhang S G, Shi J S. Physiological and proteomic analyses of leaves from the halophyte Tangut Nitraria reveals diverse response pathways critical for high salinity tolerance. Frontiers in Plant Science, 2015, 6: 30. |
| [37] | Grieve C M, Lesch S M, Mass E V, Francois L E. Leaf and spikelet primordia initiation in salt-stressed wheat. Crop Science, 1992, 33(6): 1286–1294. |
| [38] | Lu N W, Duan B L, Li C Y. Physiological responses to drought and enhanced UV-B radiation in two contrasting. Picea asperata populations, 2007, 37(7): 1253–1262. |
| [39] | Pang Q Y, Zhang A Q, Zang W, Yan X F. Integrated proteomics and metabolomics for dissecting the mechanism of global responses to salt and alkali stress in Suaeda corniculata. Plant and Soil, 2016, 402(1/2): 379–394. |
| [40] | Ruiz K B, Biondi S, Martinez E A, Orsini F, Antognoni F, Jacobsen S E. Quinoa-a model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosystems, 2016, 150(2): 357–371. DOI:10.1080/11263504.2015.1027317 |
| [41] | Shaheen H L, Iqbal M, Azeem M, Shahbaz M, Shehzadi M. K-priming positively modulates growth and nutrient status of salt-stressed cotton (Gossypium hirsutum) seedlings. Archives of Agronomy and Soil Science, 2016, 62(6): 759–768. DOI:10.1080/03650340.2015.1095292 |
| [42] | Li R, Shi F, Fukuda K. Interactive effects of various salt and alkali stresses on growth, organic solutes, and cation accumulation in a halophyte Spartina alterniflora (Poaceae). Environmental and Experimental Botany, 2010, 68(1): 66–74. DOI:10.1016/j.envexpbot.2009.10.004 |
| [43] | Dai Y L, Zhang L J, Jang S J, Yin K D. Saline and alkaline stress genotypic tolerance in sweet sorghum is linked to sodium distribution. Acta Agriculturae Scandinavica, Section B-Soil & Plant Science, 2014, 64(6): 471–481. |
| [44] | Abdel A A L, Tran L S P. Impacts of Priming with Silicon on the Growth and Tolerance of Maize Plants to Alkaline Stress. Frontiers in Plant Science, 2016, 7: 243. |
| [45] | Guo P, Wei H X, Zhang W J, Bao Y J. Physiological responses of alfalfa to high-level salt stress:root ion flux and stomatal characteristics. International Journal of Agriculture and Biology, 2016, 18(1): 125–133. |
| [46] | Dinneny J R. Traversing organizational scales in plant salt-stress responses. Current Opinion in Plant Biology, 2015, 23: 70–75. DOI:10.1016/j.pbi.2014.10.009 |
| [47] | Liu J, Cai H, Liu Y, Zhu Y M, Ji W, Bai X. A study on physiological characteristics and cmparison of salt resistance of two Medicago sativa at the seedling stage. Acta Prataculturae Sinica, 2015, 22(3): 250–256. |
| [48] | 曹齐卫, 李利斌, 孔素萍, 邱岸, 陈伟, 张允楠, 孙小镭. 不同黄瓜品种幼苗对等渗Mg(NO3)2和NaCl胁迫的生理响应. 应用生态学报, 2015, 26(4): 1171–1178. |
| [49] | Tavakkoli E, Fatehi F, Coventry S, Rengasamy P, Mcdonald G K. Additive effects of Na+ and Cl- ions on barley growth under salinity stress. Journal of Experimental Botany, 2011, 62(6): 2189–2203. DOI:10.1093/jxb/erq422 |
| [50] | 闫永庆, 刘兴亮, 王崑, 樊金萍, 石溪婵. 白刺对不同浓度混合盐碱胁迫的生理响应. 植物生态学报, 2010, 34(10): 1213–1219. DOI:10.3773/j.issn.1005-264x.2010.10.010 |
| [51] | Hazman M, Hause B, Eiche E, Nick P, Riemann M. Increased tolerance to salt stress in OPDA-deficient rice ALLENE OXIDE CYCLASE mutants is linked to an increased ROS-scavenging activity. Journal of Experimental Botany, 2015, 66(11): 3339–3352. DOI:10.1093/jxb/erv142 |
| [52] | Wang H, Lin X, Cao S, Wu Z. Alkali tolerance in rice (Oryza sativa L.):growth, photosynthesis, nitrogen metabolism, and ion homeostasis. Photosynthetica, 2015, 53(1): 55–65. DOI:10.1007/s11099-015-0079-4 |
| [53] | Gong B, Wen D, Bloszies S, Li X, Wei M, Yang F J, Shi Q H, Wang X F. Comparative effects of NaCl and NaHCO3 stresses on respiratory metabolism, antioxidant system, nutritional status, and organic acid metabolism in tomato roots. Acta Physiologiae Plantarum, 2014, 36(8): 2167–2181. DOI:10.1007/s11738-014-1593-x |
| [54] | Ranganayakulu G, Veeranagamallaiah G, Sudhakar C. Effect of salt stress on osmolyte accumulation in two groundnut cultivars (Arachis hypogaea L.) with contrasting salt tolerance. African Journal of Plant science, 2013, 7(12): 586–592. DOI:10.5897/AJPS |
| [55] | Yildirim E, Ekinici M, Turan M, Dursun A, Kul R, Parlakova F. Roles of glycine betaine in mitigating deleterious effect of salt stress on lettuce (Lactuca sativa L.). Archives of Agronomy and Soil Science, 2015, 61(12): 1673–1689. DOI:10.1080/03650340.2015.1030611 |
| [56] | Ashraf M, Foolad M R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and Experimental Botany, 2007, 59(2): 206–216. DOI:10.1016/j.envexpbot.2005.12.006 |
| [57] | 刘铎, 丛日春, 党宏忠, 李庆梅, 刘德玺, 杨庆山. 柳树幼苗渗透调节物质对中、碱性钠盐响应的差异性. 生态环境学报, 2014, 23(9): 1531–1535. |
| [58] | Butt M, Ayyub C M. Amjad M, Ahmad R. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress. Pakistan Journal of Agricultural Sciences, 2016, 53(1): 43–49. DOI:10.21162/PAKJAS |
| [59] | Adams P, Thomas J C, Vernon D M, Bohnert H J, Jensen R G. Distinct cellular and organismic responses to salt stress. Plant & Cell Physiology, 1992, 33(8): 1215–1223. |
| [60] | Khan H A, Siddique K H M, Colmer T D. Vegetative and reproductive growth of salt-stressed chickpea are carbon-limited:sucrose infusion at the reproductive stage improves salt tolerance. Journal of Experimental Botany, 2016. DOI:10.1093/jxb/erw177 |
| [61] | Guo L Q, Shi D C, Wang D L. The Key Physiological response to alkali stress by the alkali-resistant halophyte Puccinellia tenuiflora is the accumulation of large quantities of organic acids and into the rhyzosphere. Journal of Agronomy and Crop Science, 2010, 196(2): 123–135. DOI:10.1111/jac.2010.196.issue-2 |
| [62] | 刘滨硕, 康春莉, 王鑫, 包国章. 羊草对盐碱胁迫的生理生化响应特征. 农业工程学报, 2014, 30(23): 166–173. DOI:10.3969/j.issn.1002-6819.2014.23.021 |
| [63] | Chen W C, Cui P J, Sun H Y, Guo W Q, Yang C W, Jin H, Fang B, Shi D C. Comparative effects of salt and alkali stresses on organic acid accumulation and ionic balance of seabuckthorn (Hippophae rhamnoides L.). Industrial Crops and Products, 2009, 30(3): 351–358. DOI:10.1016/j.indcrop.2009.06.007 |
| [64] | 戴凌燕, 唐呈瑞, 殷奎德, 阮燕晔, 张立军. 苏打盐碱胁迫对甜高粱植株有机酸含量的影响. 生态学杂志, 2015, 34(3): 681–687. |
| [65] | 罗青红, 寇云玲, 史彦江, 宋锋惠, 韩强. 6种杂交榛对新疆盐碱土的生理适应性研究. 西北植物学报, 2013, 33(9): 1867–1873. |
| [66] | Rangani J, Parida A K, Panda A, Kumari A. Coordinated changes in antioxidative enzymes protect the photosynthetic machinery from salinity induced oxidative damage and confer salt tolerance in an extreme Halophyte salvadora persica L. Frontiers in Plant Science, 2016, 7(537): 1–18. |
| [67] | Singh N, Bhatla S C. Nitric oxide and iron modulate heme oxygenase activity as a long distance signaling response to salt stress in sunflower seedling cotyledons. Nitric Oxide, 2016, 53: 54–64. DOI:10.1016/j.niox.2016.01.003 |
| [68] | 郭楠楠, 陈学林, 张继, 陈金元, 朱愿军, 丁映童. 柽柳组培苗抗氧化酶及渗透调节物质对NaCl胁迫的响应. 西北植物学报, 2015, 35(8): 1620–1625. |
| [69] | 王鑫, 朱悦, 刘滨硕, 包国章. 盐碱胁迫下羊草抗氧化酶活性的变化. 江苏农业科学, 2015, 43(5): 209–211. |
| [70] | Scott M D, Meshnick S R, Eaton J W. Superoxide dismutase-rich bacteria. Paradoxical increase in oxidant toxicity. The Journal of Biological Chemistry, 1987, 262(8): 3640–3645. |
| [71] | 乔枫, 耿贵工. 盐碱胁迫对沙棘种子萌发及幼苗抗氧化酶活性的影响. 东北林业大学学报, 2012, 40(2): 17–19. |
| [72] | Kopittke P M. Interactions between Ca, Mg, Na and K:Alleviation of toxicity in saline solutions. Plant and Soil, 2012, 352(1/2): 353–362. |
| [73] | Abbas Z K, Mobin M. Comparative growth and physiological responses of two wheat (Triticum aestivum L.) cultivars differing in salt tolerance to salinity and cyclic drought stress. Archives of Agronomy and Soil Science, 2016, 62(6): 745–758. DOI:10.1080/03650340.2015.1083553 |
| [74] | Huang G T, Ma S L, Bai L P, Zhang L, Ma H, Jia P, Liu J, Zhong M, Guo Z F. Signal transduction during cold, salt, and drought stresses in plants. Molecular Biology Reports, 2012, 39(2): 969–987. DOI:10.1007/s11033-011-0823-1 |
| [75] | Guo R, Yang Z Z, Li F, Yan C R, Zhong X L, Liu Q, Xia X, Li H R, Zhao L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biology, 2015, 15(70): 1–13. |
| [76] | Sun J, Zhang X, Deng S R, Zhang C L, Wang M J, Ding M Q, Zhao R, Shen X, Zhou X Y, Lu C F, Chen S L. Extracellular ATP signaling is mediated by H2O2 and Cytosolic Ca2+ in the Salt Response of Populus euphratica Cells. PLoS One, 2012, 7(12): 1–15. |
| [77] | Abdula S E, Lee H J, Ryu H, Kang K K, Nou I, Sorrells M E, Cho Y G. Overexpression of BrCIPK1 gene enhances abiotic stress tolerance by increasing proline biosynthesis in rice. Plant Molecular Biology Reporter, 2016, 34(2): 501–511. DOI:10.1007/s11105-015-0939-x |
| [78] | Tian X Y, He M R, Wang Z L, Zhang J W, Song Y L, He Z L, Dong Y J. Application of nitric oxide and calcium nitrate enhances tolerance of wheat seedlings to salt stress. Plant Growth Regulation, 2015, 77(3): 343–356. DOI:10.1007/s10725-015-0069-3 |
| [79] | Wang X P, Geng S J, Ma Y Q, Shi D C, Yang C W, Wang H. Growth, photosynthesis, solute accumulation, and ion balance of tomato plant under sodium-or potassium-salt stress and alkali stress. Agronomy Journal, 2015, 107(2): 651–661. DOI:10.2134/agronj14.0344 |
| [80] | 郭文芳, 农万廷, 李刚强, 刘德虎. 植物耐盐碱基因工程研究进展. 生物技术通报, 2015, 31(7): 11–17. |
| [81] | Rubio J S, García-Sánchez F, Rubio F, Martinez V. Yield, blossom-end rot incidence, and fruit quality in pepper plants under moderate salinity are affected by K+ and Ca2+ fertilization. Scientia Horticulturae, 2009, 119(2): 79–87. DOI:10.1016/j.scienta.2008.07.009 |
| [82] | Han Y, Yin S Y, Huang L. Towards plant salinity tolerance-implications from ion transporters and biochemical regulation. Plant Growth Regulation, 2015, 76(1): 13–23. DOI:10.1007/s10725-014-9997-6 |
| [83] | Guan Q J, Wang Z J, Wang X H, Takano T, Liu S K. A peroxisomal APX from Puccinellia tenuiflora improves the abiotic stress tolerance of transgenic Arabidopsis thaliana through decreasing of H2O2accumulation. Journal of Plant Physiology, 2015, 175: 183–191. DOI:10.1016/j.jplph.2014.10.020 |
| [84] | Ren S X, Lyle C, Jiang G L, Penumala A. Soybean salt tolerance 1(GmST1) reduces ros production, enhances aba sensitivity, and abiotic stress tolerance in Arabidopsis thaliana. Frontiers in Plant Science, 2016, 7: 445. |
| [85] | Han Z F, Yu H M, Zhao Z, Hunter D, Luo X J, Duan J, Tian L N. AtHD2D gene plays a role in plant growth, development, and response to abiotic stresses in Arabidopsis thaliana. Frontiers in Plant Science, 2016, 7: 310. |
| [86] | Li L Q, Xie J H, Ma X Q, Li D. Molecular cloning of Phosphoethanolamine N-methyltransferase (PEAMT) gene and its promoter from the halophyte Suaeda liaotungensis and their response to salt stress. Acta Physiologiae Plantarum, 2016, 38(2): 39. DOI:10.1007/s11738-016-2063-4 |
| [87] | Villicaña C, Warner N, Arce-Montoya M, Rojas M, Angulo C, Orduño A, Gómez-Anduro G. Antiporter NHX2 differentially induced in Mesembryanthemum crystallinum natural genetic variant under salt stress. Plant Cell, Tissue and Organ Culture, 2016, 124(2): 361–375. DOI:10.1007/s11240-015-0900-6 |
| [88] | Luo P, Shen Y X, Jin S X, Huang S S, Cheng X, Wang Z, Li P L, Zhao J, Bao M Z, Ning G G. Overexpression of Rosa rugosa anthocyanidin reductase enhances tobacco tolerance to abiotic stress through increased ROS scavenging and modulation of ABA signaling. Plant Science, 2016, 245: 35–49. DOI:10.1016/j.plantsci.2016.01.007 |
| [89] | 张建秋, 陆海, 王智, 杜希华, 张玉玲. 双向电泳技术分析白刺盐胁迫蛋白的表达. 吉林农业大学学报, 2004, 26(5): 511–514. |
| [90] | Carrondo M A. Ferritins, Iron uptake and storage from the bacterioferritin viewpoint. New EMBO Member's Review, 2003, 22(9): 1959–1968. |
| [91] | Hegedüs A, Janda T, Horváth G V, Dudits D. Accumulation of overproduced ferritin in the chloroplast provides protection against photoinhibition induced by low temperature in tobacco plants. Journal of Plant Physiology, 2008, 165(15): 1647–1651. DOI:10.1016/j.jplph.2008.05.005 |
| [92] | DuanMu H Z, Wang Y, Bai X, Cheng S F, Deyholos M K, Wong G K S, Li D, Zhu D, Li R, Yu Y, Cao L, Chen C, Zhu Y M. Wild soybean roots depend on specific transcription factors and oxidation reduction related genesin response to alkaline stress. Functional & Integrative Genomics, 2015, 15(6): 651–660. |
| [93] | Qiu Y W, Wen H T, Ao J X, Zhang M H, Li F L. Soy 14-3-3 protein SGF14c, a new regulator of tolerance to salt-alkali stress. Plant Biotechnology Reports, 2015, 9(6): 369–377. DOI:10.1007/s11816-015-0374-3 |
| [94] | Liu A L, Yu Y, Duan X B, Sun X L, Duanmu H Z, Zhu Y M. GsSKP21, a Glycine soja S-phase kinase-associated protein, mediates the regulation of plant alkaline tolerance and ABA sensitivity. Plant Molecular Biology, 2015, 87(1/2): 111–124. |
| [95] | Xu Z L, Ali Z, Xu L, He X L, Huang Y H, Yi J X, Shao H B, Ma H X, Zhang D Y. The nuclear protein GmbZIP110 has transcription activation activity and plays important roles in the response to salinity stress in soybean. Scientific Reports, 2016, 6: 1–12. DOI:10.1038/s41598-016-0001-8 |
| [96] | Gong B, Li X, VandenLangenberg K M, Wen D, Sun S S, Wei M, Li Y, Yang F J, Shi Q H, Wang X F. Overexpression of S-adenosyl-L-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant Biotechnology Journal, 2014, 12(6): 694–708. DOI:10.1111/pbi.2014.12.issue-6 |
| [97] | Hu D G, Ma Q J, Sun C H, Sun M H, You C X, Hao Y J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiologia Plantarum, 2016, 156(2): 201–214. DOI:10.1111/ppl.2016.156.issue-2 |
| [98] | Sun X L, Yang S S, Sun M Z, Wang S T, Ding X D, Zhu D, Ji W, Cai H, Zhao C Y, Wang X D, Zhu Y M. A novel Glycine soja cysteine proteinase inhibitor GsCPI14, interacting with the calcium/calmodulin-binding receptor-like kinase GsCBRLK, regulated plant tolerance to alkali stress. Plant Molecular Biology, 2014, 85(1/2): 33–48. |
| [99] | 赵莹, 杨克军, 李佐同, 赵长江, 徐晶宇, 胡雪微, 石新新, 马丽峰. 外源糖浸种缓解盐胁迫下玉米种子萌发. 应用生态学报, 2015, 26(9): 2735–2742. |
| [100] | Liang X L, Fang S M, Ji W B, Zheng D F. The positive effects of silicon on rice seedlings under saline-alkali mixed stress. Communications in Soil Science and Plant Analysis, 2015, 46(17): 2127–2138. DOI:10.1080/00103624.2015.1059848 |
| [101] | Li Y T, Zhang W J, Cui J J, Lang D Y, Li M, Zhao Q P, Zhang X H. Silicon nutrition alleviates the lipid peroxidation and ion imbalance of Glycyrrhiza uralensis seedlings under salt stress. Acta Physiologiae Plantarum, 2016, 38: 96. DOI:10.1007/s11738-016-2108-8 |
| [102] | 周万海, 冯瑞章, 师尚礼, 寇江涛. NO对盐胁迫下苜蓿根系生长抑制及氧化损伤的缓解效应. 生态学报, 2015, 35(11): 3606–3614. |
| [103] | 向丽霞, 胡立盼, 胡晓辉, 潘雄波, 任文奇. 外源γ-氨基丁酸调控甜瓜叶绿体活性氧代谢应对短期盐碱胁迫. 应用生态学报, 2015, 26(12): 3746–3752. |
| [104] | Ding H, Lai J B, Wu Q, Zhang S C, Chen L, Dai Y S, Wang C F, Du J J, Xiao S, Yang C W. Jasmonate complements the function of Arabidopsis lipoxygenase 3 in salinity stress response. Plant Science, 2016, 244: 1–7. DOI:10.1016/j.plantsci.2015.11.009 |
| [105] | Me X Q, Zhang J, Huang B R. Cytokinin-mitigation of salt-induced leaf senescence in perennial ryegrass involving the activation of antioxidant systems and ionic balance. Environmental and Experimental Botany, 2016, 125: 1–11. DOI:10.1016/j.envexpbot.2016.01.002 |
| [106] | Liu R Q, Xu X J, Wang S, Shan C J. Lanthanum improves salt tolerance of maize seedlings. Photosynthetica, 2016, 54(1): 148–151. DOI:10.1007/s11099-015-0157-7 |
| [107] | Jiang C Q, Cui Q R, Feng K, Xu D F, Li C F, Zheng Q S. Melatonin improves antioxidant capacity and ion homeostasis and enhances salt tolerance in maize seedlings. Acta Physiologiae Plantarum, 2016, 38(4): 82. DOI:10.1007/s11738-016-2101-2 |
| [108] | 张毅, 石玉, 胡晓辉. 外源亚精胺对盐碱胁迫下番茄幼苗光合特性的影响. 西北农林科技大学学报:自然科学版, 2016, 44(2): 144–150. |
| [109] | 张毅, 石玉, 胡晓辉, 邹志荣, 曹凯, 张浩. 外源Spd对盐碱胁迫下番茄幼苗氮代谢及主要矿质元素含量的影响. 应用生态学报, 2013, 24(5): 1401–1408. |
| [110] | 潘雄波, 向丽霞, 胡晓辉, 任文奇, 张丽, 倪新欣. 外源亚精胺对盐碱胁迫下番茄幼苗根系线粒体功能的影响. 应用生态学报, 2016, 27(2): 491–498. |
| [111] | Chunthaburee S, Sanitchon J, Pattanagul W, Theerakulpisut P. Application of exogenous spermidine (Spd) improved salt tolerance of rice at the seedling and reproductive stages. Procedia Environmental Sciences, 2015, 29: 134. DOI:10.1016/j.proenv.2015.07.224 |
| [112] | 张义飞, 王平, 毕琪, 张忠辉, 杨允菲. 不同强度盐胁迫下AM真菌对羊草生长的影响. 生态学报, 2016, 36(17): 1–10. |
| [113] | Porcel R, Aroca R, Ruiz-Lozano J M. Salinity stress alleviation using arbuscular mycorrhizal fungi:A review. Agronomy for Sustainable Development, 2012, 32(1): 181–200. DOI:10.1007/s13593-011-0029-x |
| [114] | Al-Karaki G N, Hammad R, Rusan M. Response of two tomato cultivars differing in salt tolerance to inoculation with mycorrhizal fungi under salt stress. Mycorrhiza, 2001, 11(1): 43–47. DOI:10.1007/s005720100098 |
| [115] | Zarea M J, Karimi N, Goltapeh E M, Ghalavand A. Effect of cropping systems and arbuscular mycorrhizal fungi on soil microbial activity and root nodule nitrogenase. Journal of the Saudi Society of Agricultural Sciences, 2011, 10(2): 109–120. DOI:10.1016/j.jssas.2011.04.003 |
| [116] | 郭伶娜, 刘建文, 麻冬梅, 许兴. 高羊茅转基因植株的耐盐性鉴定. 中国草地学报, 2015, 37(1): 116–120. |
| [117] | Zhu D, Cai H, Luo X, Bai X, Deyholos M K, Chen Q, Chen C, Ji W, Zhu Y M. Over-expression of a novel JAZ family gene from Glycine soja, increases salt and alkali stress tolerance. Biochemical and Biophysical Research Communications, 2012, 426(2): 273–279. DOI:10.1016/j.bbrc.2012.08.086 |
| [118] | Liu A L, Yu Y, Li R T, Duan X B, Zhu D, Sun X L, Duanmu H Z, Zhu Y M. A novel hybrid proline-rich type gene GsEARLI17 from Glycine soja participated in leaf cuticle synthesis and plant tolerance to salt and alkali stresses. Plant Cell, Tissue and Organ Culture, 2015, 121(3): 633–646. DOI:10.1007/s11240-015-0734-2 |
| [119] | Guo M, Liu Q, Yu H, Zhou T T, Zou J, Zhang H, Bian M D, Liu X M. Characterization of alkali stress-responsive genes of the CIPK family in sweet sorghum[Sorghum bicolor (L.) Moench]. Crop Science, 2015, 55(3): 1254–1263. DOI:10.2135/cropsci2013.08.0520 |
| [120] | Liu L P, Zhang Z Q, Dong J L, Wang T. Overexpression of MtWRKY76 increases both salt and drought tolerance in Medicago truncatula. Environmental and Experimental Botany, 2016, 123: 50–58. DOI:10.1016/j.envexpbot.2015.10.007 |
| [121] | Jia B W, Sun M Z, Sun X L, Li R T, Wang Z Y, Wu J, Wei Z W, DuanMu H Z, Xiao J L, Zhu Y M. Overexpression of GsGSTU 13 and SCMRP in Medicago sativa confers increased salt-alkaline tolerance and methionine content. Physiologia Plantarum, 2016, 156(2): 176–189. DOI:10.1111/ppl.2016.156.issue-2 |
| [122] | Bulle M, Yarra R, Abbagani S. Enhanced salinity stress tolerance in transgenic chilli pepper (Capsicum annuum L.) plants overexpressing the wheat antiporter (TaNHX2) gene. Molecular Breeding, 2016, 36: 36. DOI:10.1007/s11032-016-0451-5 |
| [123] | Zarza X, Atanasov K E, Marco F, Arbona V, Carrasco P, Kopka J, Fotopoulos V, Munnik T, Gómez-Cadenas A, Tiburcio A F, Alcázar R. Polyamine oxidase 5 loss-of-function mutations in Arabidopsis thaliana trigger metabolic and transcriptional reprogramming and promote salt stress tolerance. Plant, Cell & Environment, 2016, doi:10.1111/pce.12714. |
| [124] | Hussain S W, Williams W M. Chromosome pairing and fertility of interspecific hybrids between Trifolium repens L. and T. occidentale Coombe. Plant Breeding, 2016, 135(2): 239–245. DOI:10.1111/pbr.2016.135.issue-2 |
| [125] | Mohsenian Y, Roosta H R. Effects of grafting on alkali stress in tomato plants:Datura rootstock improve alkalinity tolerance of tomato plants. Journal of Plant Nutrition, 2015, 38(1): 51–72. DOI:10.1080/01904167.2014.920370 |
| [126] | Pastor V, Luna E, Mauch-Mani B, Ton J, Flors V. Primed plants do not forget. Environmental and Experimental Botany, 2013, 94: 46–56. DOI:10.1016/j.envexpbot.2012.02.013 |
| [127] | Jiménez-Arias D, Pérez J A, Luis J C, Martín-Rodríguez V, Valdés-González F, Borges A A. Treating seeds in menadione sodium bisulphite primes salt tolerance in Arabidopsis by inducing an earlier plant adaptation. Environmental and Experimental Botany, 2015, 109: 23–30. DOI:10.1016/j.envexpbot.2014.07.017 |
| [128] | Jan N E, Din J U, Kawabata S. Impact of saline-alkalistress on the accumulation of solids in tomato fruits. Pakistan Journal of Botany, 2014, 46(1): 161–166. |
| [129] | Bai J H, Liu J H, Zhang N, Yang J H, Sa R L, Wu L. Effect of Alkali Stress on Soluble Sugar, Antioxidant Enzymes and Yield of Oat. Journal of Integrative Agriculture, 2013, 12(8): 1441–1449. DOI:10.1016/S2095-3119(13)60556-0 |
| [130] | Wang Z Y, Zhao X Y, Wang B, Liu E L, Chen N, Zhang W, Liu H. Overexpression of an Arabidopsis heterogeneous nuclear ribonucleoprotein gene, AtRNP1, affects plant growth and reduces plant tolerance to drought and salt stresses. Biochemical and Biophysical Research Communications, 2016, 472(2): 353–359. DOI:10.1016/j.bbrc.2016.02.081 |