 2017, Vol. 37
2017, Vol. 37文章信息
- 刘家霖, 满秀玲
- LIU Jialin, MAN Xiuling.
- 降雨和非降雨日兴安落叶松天然林蒸腾及蒸散发特征
- Characteristics of transpiration and evapo-transpiration from natural Larix gmelinii forests on rainy and non-rainy days
- 生态学报. 2017, 37(15): 5059-5069
- Acta Ecologica Sinica. 2017, 37(15): 5059-5069
- http://dx.doi.org/10.5846/stxb201605050858
-
文章历史
- 收稿日期: 2016-05-05
- 网络出版日期: 2017-03-22

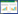




蒸散发(ET)主要包括自由水蒸发和植物蒸腾耗水(T)[1-2], 是地表能量平衡和全球水循环的重要组成部分[3-5]。Jasechko等[2]的研究指出T是陆地表面最大的水汽通量来源, 占全球陆地生态系统ET的80%—90%;而Schlesinger等[5]通过对全球81个不同生态区域的T和ET进行整合分析, 发现热带雨林的T/ET最高(平均为70%), 北方针叶林其次(平均为65%), 地中海灌木林最低(平均为47%)。可见, 植物的蒸腾耗水是陆地生态系统水分平衡的主要贡献者和全球水分循环的主导力量[5-6]。当前, 树干液流法(SF)和涡动相关技术(EC)分别是T和ET的主要测定手段[1, 4-5, 7-8]。SF法主要利用热扩散原理, 通过测定插入树干边材的热探针与参比探针的温度差或电压差的方法来估算液流速率(Fs)[9]。应用热技术测定树干液流的方法已十分成熟[7], 但该技术仅能对单木尺度T进行测定, 需要利用胸径[10]或边材面积[11]等指标进行尺度扩展, 进而估算林分尺度的蒸腾耗水量。EC是指某种物质的浓度与其垂直速度的协方差[12], 自1990年Wofsy等[13]利用EC技术对温带森林的CO2通量进行测定以来, 该技术已经成为直接测定植物-大气界面气体交换通量的标准方法[8]。利用EC技术可以对ET进行连续测定, 但难以区分蒸发或T等动态过程对ET的贡献程度, 而结合SF法可以实现对ET及其组分的同步研究[1, 4-5, 7, 14]。然而, 国内以ChinaFLUX为代表的EC站点大多位于亚热带和温带区域, 缺乏对大兴安岭北部寒温带森林生态系统碳-水通量的系统性了解[1]。
降雨是生态系统水分输入的主要途径[5], 水分状况通过影响净辐射(Rn)在显热通量(H)和潜热通量(LE)等能量组分间的分配比例[15], 进而改变空气水热状况及水汽压亏缺(VPD)等林内微生境[16-17], 从而实现对T和ET的综合调控[4, 16-19]。同时, 以往对树干液流的研究多集中于林分内的优势木[5, 20], 往往忽略对中等木和劣势木的监测, 在尺度扩展过程中易造成误差[21]。此外, 对于郁闭度较高的森林, 林下植被的液流密度(Fd)远低于乔木[22-23], 因而乔木T在生态系统蒸腾耗水量中明显占优, 且该现象在夜间时更为明显[24], 通常忽略对林下植被蒸腾耗水量的估算[15]。
兴安落叶松(Larix gmelinii)林是大兴安岭地区的地带性植被和中国唯一的寒温带针叶林优势建群种[25], 认识该区域典型森林生态系统的水汽通量特征, 有助于了解区域能量平衡和水分循环, 对于全面理解和掌握我国不同类型森林生态系统的水汽循环也具有重要意义。该研究于2015年8月12日—9月12日, 在兴安落叶松天然林中选取优势木、中等木和劣势木各3株, 综合运用SF法和EC技术对林木T和生态系统ET进行连续监测, 分析降雨和非降雨日的水汽通量变化特征及其对主要环境因子的响应, 以了解寒温带典型落叶松林的T和ET规律, 为大兴安岭北部水资源的有效利用和科学管理提供科学依据和数据支持。
1 试验地概况研究地设在黑龙江漠河森林生态系统研究站(53.27°—53.28° N, 122.33°—122.34° E), 属寒温带大陆性气候, 多年冻土区, 平均海拔290 m, 年均温-4.9 ℃, 多年平均降水量350—500 mm, 全年无霜期85—110 d。地带性土壤为棕色针叶林土, 地带性植被是以兴安落叶松为优势建群种的明亮针叶林。其他常见树种为樟子松(Pinus sylvestris)、白桦(Betula platyphylla)和山杨(Populus davidiana);常见林下植被为杜香(Ledum palustre)、笃斯越桔(Vaccinium uliginosum)、兴安杜鹃(Rhododendron dauricum)和越桔(V. vitis-idaea)。该研究于2015年7月上旬(6—9月为该地区兴安落叶松林的典型生长季)在EC系统东北侧设置15 m × 20 m的样地, 样地内共有兴安落叶松84株、白桦14株、樟子松1株;该研究区下垫面平坦均匀, 符合EC技术观测要求(图 1)。

|
| 图 1 研究区域示意图 Fig. 1 Schematic plot of research area |
在胸高处南向用生长锥(CO200, Haglof, Sweden)对选择的60株样树钻取深至髓心的树芯;样芯经自然干燥、固定和打磨后进行扫描;使用WinDENDRO年轮分析软件(Regent, Canada)对扫描图像进行处理, 估测树龄和边材宽度。结果显示, 胸径处于5—10 cm范围的树龄为(37.3±11.8) a, 10—20 cm为(43.7±9.5) a, 20—25 cm为(53.4±8.7) a。兴安落叶松天然林的林分结构较为复杂, 林龄差在一个龄级内(20 a), 可视为相对同龄林[26]。同龄林中, 林木分化主要受林木个体之间的竞争优势及空间生态位决定[27]。因此, 采用相对直径法[26-27]划分林木分化等级, 优势木、中等木和劣势木数量分别为28株、30株和25株。不同分化等级林木的胸径(F2, 82= 145.8)、树高(F2, 82= 162.3) 和边材面积(F2, 82= 110.0) 均差异显著(P<0.01)。选择长势良好、未经处理的不同分化等级林木各3株作为SF法的监测样树(表 1)。
| 林木分化等级 Tree differentiation classes |
样木数 Samples |
胸径 DBH/cm |
树高 Tree height/m |
边材面积 Sapwood area/cm2 |
| 优势木 Dominant tree | 3 | 23.53±2.10 | 20.75±1.20 | 131.00±18.94 |
| 中等木 Intermediate tree | 3 | 11.17±2.73 | 12.82±2.03 | 39.15±15.33 |
| 劣势木 Suppressed tree | 3 | 6.01±0.82 | 8.66±0.76 | 13.79±3.03 |
该研究使用的热扩散式液流探针是依据Granier[28]的探针原理, 采用James等[29]方法制作, 测定结果可靠[30]。结合以往研究[31]和该研究对生长芯的测定结果, 计算边材宽度与胸径的关系;结果显示, 优势木各方位边材宽度均大于2 cm, 而中等木、劣势木北向边材宽度分别为(1.3±0.1) cm和(1.1±0.1) cm。因此, 为尽量减小树干液流方位变异和太阳辐射增温现象的影响, 优势木在东、西、北向分别安装液流探针, 而中等木和劣势木则仅安装在北向[9];中等木和劣势木的探针安装时需斜向插入边材(即与茎轴夹角<90°), 保证探针只与边材接触。兴安落叶松天然林木的胸径与边材面积间具有较好的相关性(式1), 采用边材面积对单木T进行尺度扩展[11, 15]。该研究中的T指广义的植物蒸腾作用, Td指优势木蒸腾量, Ti指中等木蒸腾量, Ts指劣势木蒸腾量, Ttot指林木总蒸腾量(Ttot=Td+ Ti+ Ts)。Fd(cm3 cm-2 h-1)、液流速率(Fs, g/h)和T(mm)的计算公式分别如下[9-10, 32]:
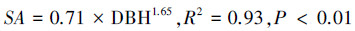
|
(1) |
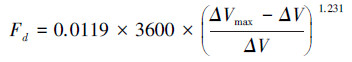
|
(2) |
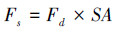
|
(3) |
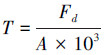
|
(4) |
式中, DBH是胸径(cm), SA是边材面积(cm2), ΔVmax是液流为0时的电压, ΔV是某一测定时刻的电压, A为样地面积(m2)。
2.2 涡动相关技术 2.2.1 仪器配置EC系统配置一体式三维超声风与红外CO2/H2O分析仪(IRGASON, Campbell Scientific, USA), 安装高度36 m。梯度气象系统包括:空气温、湿度(HMP45C, Vaisala, Finland)和风速(010C, Met One, USA)传感器, 安装高度分别为1.5、9、13、20、28和35 m;风向传感器(020C, Met One, USA), 安装高度28 m;Rn传感器(NR01, Hukseflux, Netherlands)、光合有效辐射传感器(LI190SB, Li-Cor, USA)、雨滴谱仪(Parsivel2, OTT, Germany)和雨量筒(TE525MM, Texas Electronics, USA), 安装高度23 m;大气压传感器(CS100, Campbell Scientific, USA), 安装高度23 m;土壤热通量传感器(HFP01, Hukseflux, Netherlands), 分别安装于距通量塔东、西两侧10 m远, 距离地面10 cm深处;土壤水分、温度和电导率传感器(CS650, Campbell Scientific, USA), 安装于距通量塔东侧10 m远, 距离地面5、10、20和40 cm深处。采用CR3000数据采集器(Campbell Scientific, USA)以10 Hz频率记录通量的观测数据;气象数据每隔30 min保存其平均数据;所有监测仪器均同步运行, 观测从2014年11月下旬开始。
2.2.2 环境因子数据在通量塔梯度气象系统的基础上, 该研究选取的主要环境因子包括:23 m林冠层以上的Rn和降雨量(P), 13 m林冠层的空气温度(Ta)、空气相对湿度(RH)和VPD, 36 m IRGASON所记录的风速与风向以及4层土深的平均土壤温度(Ts)和4层土深的加权平均土壤含水量(SWC)。在分析过程中, 该研究定义有降雨发生(即P>0.2 mm)的观测日为降雨日, 无降雨发生(即P=0 mm)的观测日为非降雨日, 而不以降雨发生作为条件对观测数据进行分割即为不区分降雨日。
2.2.3 通量数据使用LoggerNet软件(Campbell Scientific, USA)将观测期间10Hz原始数据按日期切割为30 min间隔的数据序列;利用Eddypro软件(Li-Cor, USA)计算通量数据, 主要步骤包括剔除野点、线性去趋势、平面拟合、超声虚温订正、密度扰动影响、湍流平稳性检验、总体湍流特征检验、数据质量评价和数据质量等级划分等。同时, 软件的处理结果需进行以下校正[32]:(1) 该研究对摩擦风速(u*)进行综合敏感性分析(0.5 m/s间隔)[33], 发现在观测期间任意时刻下, 当u*<0.20 m/s时, 湍流强度较弱并对LE的结果产生影响, 剔除该部分数据;(2) 剔除每场降雨前后各1 h的数据。最终, 缺失数据量为18%。此外, 采用Kljun等[34]的模型进行通量足迹分析, 发现通量贡献区70%和90%的平均距离分别为846和1133 m, 观测期间的主风向为东西向和东西偏北向, 观测的湍流范围包括SF法监测区域。
2.2.4 缺失数据插补采用以BP网络(BP, Back-Propagation)作为基本网络的人工神经网络法进行缺失数据的插补。输入层的环境因子以Penman-Monteith模型[35]参数为基础, 包括代表能量分配与变化的Rn、Ta和土壤热通量(G), 代表研究区域上层大气湍流变化的w和RH, 代表土壤水分状况的SWC。BP网络模型采用输入层、隐含层和输出层的3层式结构, 每层包含若干节点, 输入层的设计公式如下:
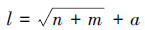
|
(5) |
式中, l为隐含层节点数, n为输入节点数(该研究为6), m为输出节点数(该研究为1);α为1—10之间的调节常数。通过改变α的值使训练误差达到最小, 从而获得隐含层最佳的节点数目;最终, l值取10, LE拟合结果的R2为0.81。
2.2.5 能量闭合分析能量平衡闭合水平是评价EC技术观测数据可靠性的主要方法[36]。森林生态系统的能量平衡通常为Rn=H + LE + G + S, S指冠层热储量[25]。郁闭森林生态系统的G和S所占的比重低于5%, 常忽略不计[37]。由于辐射平衡和能量分量的测定仪器不同, 空间范围不一致, 二者之间通常不能达到完全闭合。该研究中, 以观测期间质量控制后每小时的H + LE与Rn-G进行线性回归, 结果显示:方程斜率为1.75, 截距为18.04, R2为0.77。
3 结果和分析 3.1 环境因子特征分析观测期间, 共有12 d发生降雨, 累计P为65.8 mm, 作为典型降雨日进行分析(图 2)。Rn的波动与降雨发生有关(图 2), 降雨日的辐射能量往往较低且波动明显, 平均为6.8 MJ m-2 d-1, 而非降雨日的变化则略平缓, 平均为11.8 MJ m-2 d-1;同时, 根据Rn= 4 340.00-45.60 × Lat(纬度)[37], 该研究区域Rn年总收入约1911 MJ m-2 d-1, 而观测期间Rn总收入约317.4 MJ/m2, 占全年总收入理论值的16.6%。VPD与空气水热状况(即Ta与RH)密切相关, 能量分配与水分输入均会对其产生影响[7, 10-11], 其最低值出现在降雨日(0.055±0.005) kPa, 而最高值则出现在非降雨日(1.009±0.163) kPa(图 2)。观测期间的最高日均温出现在224 d(21.3±1.0)℃, 最低日均温出现在255 d(8.5±1.2)℃, 降雨日Ta的波动较明显, 但总体上呈下降趋势(图 2);Ts主要受Ta影响, 其从224 d时的(14.6±0.1)℃连续下降至255 d的(10.1±0.1)℃。同时, 降雨日RH和SWC分别平均可达到84.8%和0.28 m3/m3, 而非降雨日时则分别平均为77.5%和0.25 m3/m3。此外, 在对比降雨和非降雨日VPD、Rn的差异性时, 以Ta为控制变量进行偏相关分析, 结果显示P与VPD(r = -0.5)、Rn(r =-0.7) 均显著负相关(P<0.01);而方差分析表明降雨与非降雨日的VPD(F1, 31= 16.6)、Rn(F1, 31= 12.8) 均差异显著(P<0.01), 说明剔除Ta的影响后降雨和非降雨日的VPD与Rn均存在差异。

|
| 图 2 观测期间环境因子变化规律 Fig. 2 Variation of environmental factors during observation period (mean±SD) |
方差分析表明降雨和非降雨日期间Ttot(F1, 766=18.3) 和ET(F1, 766=4.8) 均差异显著(P<0.05)。图 3将各分化等级林木Fs分别按日时刻进行平均, 其昼夜变化特征均呈单峰格局。在不区分降雨日时(图 3), 优势木、中等木和劣势木的Fs峰值分别为(1185.7±93.5)、(372.3±30.5) 和(122.7±9.9) g/h;降雨日(图 3), 三者的Fs明显降低, 峰值出现在13:00, 分别为(744.1±136.6)、(229.8±45.4) 和(77.0±15.6) g/h;而非降雨日(图 3), 三者的Fs峰值较降雨日提前1 h, 峰值分别可达到(1459.3±64.3)、(461.0±21.5) 和(147.9±6.6) g/h。总体上, 非降雨日各分化等级林木均表现出高蒸腾的特征, 即日液流峰值出现时间较早、峰值较高。

|
| 图 3 观测期间各分化等级样木平均液流速率的昼夜变化 Fig. 3 Diurnal dynamics of mean sap flow velocity of sample trees during observation period (mean±SE) |
图 4为观测期内ET和T的连续变化特征。利用各分化等级林木Fs和SA对单木T进行尺度上推, 分别计算优势木蒸腾Td、中等木蒸腾Ti和劣势木蒸腾Ts以及林木总蒸腾(Ttot), 结果表明:不区分降雨日时Ttot为41.1 mm, 约为1.28 mm/d;降雨日时, Td、Ti和Ts分别为6.9、2.2和0.6 mm;而非降雨日, Td、Ti和Ts分别为23.0、7.3和2.0 mm。同时, 不区分降雨日时的ET为75.3 mm, 而降雨和非降雨日时的ET则分别为24.7、50.6 mm, 且降雨日的ET能力(2.1 mm/d)低于非降雨日(2.5 mm/d)。从能量角度分析, 观测期间LE总量为84.3 MJ/m2, 占同期Rn的28.1%;其中, 降雨日的LE为25.2 MJ/m2, 与Rn之比为31%;而非降雨日LE为59.1 MJ/m2, 占比25.1%。上述结果表明:降雨日Ttot低于非降雨日, 降雨和非降雨日时林分内Td均显著占优, 明显高于中等木和劣势木的蒸腾耗水量;降雨日的太阳辐射能量更多的分配至LE, 从而维持一定的ET需求;此外, Pearson相关性分析表明, ET和Ttot均与P呈显著的负相关关系(r分别为-0.4和-0.7), 且水分输入状况对Ttot的影响更为明显, 非降雨日的天气条件更有利于植物-大气界面的水汽通量交换。

|
| 图 4 观测期间蒸散发(ET)和蒸腾(T)的变化特征 Fig. 4 Variations of daily cumulative evapo-transpiration (ET) and transpiration (T) during observation periods 图中ET指蒸散发, Ttot指林木总蒸腾, Td指优势木蒸腾, Ti指中等木蒸腾, Ts指劣势木蒸腾 |
不区分降雨日时, Ttot/ET可以达到54.9%;其中, 各蒸腾组分Td/ET、Ti/ET和Ts/ET分别为38.6%、12.8%和3.5%(图 5)。然而受生态系统水分输入的影响, 该比值波动较大。降雨期日, Ttot/ET、Td/ET、Ti/ET和Ts/ET分别为38.1%、27.2%、8.5%和2.4%, 均低于其在非降雨日时的65.0%、45.5%、15.3%和4.2%。LSD多重比较分析表明各降雨状况分组的组内T与ET之比均差异显著(P<0.05)。为了直观展示不同降雨状况分组的Ttot及其蒸腾组分与ET之比的昼夜变化, 以日时间序列为分类标准对数据进行平均, 结果显示:降雨和非降雨日(图 6), Ttot及其蒸腾组分与ET之比均在夜间较低、日间较高, 呈典型的单峰格局, 与树干液流的日变化规律相似(图 3), 峰值均出现在约17:00;降雨日(图 6), Ttot/ET、Td/ET、Ti/ET和Ts/ET的日峰值分别为59.2%、42.8%、13.6%和3.74%;非降雨日(图 6), 上述比值的日峰值分别可以达到84.9%、60.2%、19.4%和5.4%。以上结果表明降雨日时的ET以自由水蒸发为主, 而非降雨日时则以Td占优。

|
| 图 5 不同降雨状况分组的林木蒸腾与蒸散发之比的平均值 Fig. 5 The average ratio of transpiration (T) to evapo-transpiration (ET) in different tree differentiation classes under different precipitation components during observation period (mean±SE) 图中不同小写字母表示不同降雨状况分组的组内Ttot及其蒸腾组分与ET比值间差异的显著性(P<0.05);ET指蒸散发, Ttot指林木总蒸腾, Td指优势木蒸腾, Ti指中等木蒸腾, Ts指劣势木蒸腾 |

|
| 图 6 观测期间不同降雨状况分组的按时刻平均的蒸腾与蒸散发之比的昼夜变化 Fig. 6 Diurnal dynamics of hourly averaged ratio of transpiration (T) to evapo-transpiration (ET) in different tree differentiation classes under different precipitation components during observation periods Ttot指林木总蒸腾, Td指优势木蒸腾, Ti指中等木蒸腾, Ts指劣势木蒸腾 |
不同降雨状况分组间, Ttot及其蒸腾组分与ET的回归方程R2以非降雨日(平均为0.72) 较高, 不区分降雨日(平均为0.56;代表降雨日与非降雨日数据的平均水平)其次, 降雨日(平均为0.51) 较低;同时, 降雨日和非降雨日Td与ET的方程斜率和R2均高于总体水平, 而Ti和Ts则均低于总体水平, 上述结果表明水分输入会影响T及ET的相对稳定性, 且不论是否存在水分输入Td对ET的贡献性均较高(表 2)。
| 降雨状况分组 Precipitation components |
蒸腾 Transpiration |
截距 Intercept |
斜率 Slope |
R2 | P |
| 不区分降雨日 | 优势木蒸腾 | 0.012 | 0.266 | 0.570 | <0.01 |
| All days | 中等木蒸腾 | 0.003 | 0.089 | 0.566 | <0.01 |
| 被压木蒸腾 | 0.001 | 0.024 | 0.552 | <0.01 | |
| 林木总蒸腾 | 0.017 | 0.379 | 0.569 | <0.01 | |
| 降雨日 | 优势木蒸腾 | 0.003 | 0.198 | 0.516 | <0.01 |
| Rainy days | 中等木蒸腾 | 0.001 | 0.064 | 0.500 | <0.01 |
| 被压木蒸腾 | 0.000 | 0.018 | 0.510 | <0.01 | |
| 林木总蒸腾 | 0.003 | 0.280 | 0.513 | <0.01 | |
| 非降雨日 | 优势木蒸腾 | 0.009 | 0.298 | 0.728 | <0.01 |
| Non-rainy days | 中等木蒸腾 | 0.003 | 0.099 | 0.721 | <0.01 |
| 被压木蒸腾 | 0.001 | 0.027 | 0.710 | <0.01 | |
| 林木总蒸腾 | 0.013 | 0.424 | 0.726 | <0.01 |
以降雨的发生情况为固定变量, VPD和Rn为协变量进行方差分析, 结果表明降雨和非降雨日的Ttot和ET均受Rn、VPD的显著影响(P<0.01);同时, 选择代表林木蒸腾耗水总体水平的Ttot以及生态系统ET作为对环境因子响应分析的因变量。降雨日和非降雨日时的Ttot对VPD响应程度均较高(R2均为0.69;图 7), 表明降雨和非降雨日时的VPD都是Ttot的限制性因子;非降雨日, Ttot随VPD的增加而逐渐增强的范围较降雨日明显, 说明生态系统水分输入较低时利于维持林木的蒸腾需求。降雨日, ET受VPD的影响较小(R2为0.23;图 7), 而在非降雨日则略高(R2为0.32);虽然降雨日的VPD总体水平较低, 但ET的离散程度较大、数值范围较广, 在非降雨日具有相似的表现。同时, Ttot和ET与VPD的回归方程斜率均在降雨日时较高, 说明降雨日生态系统水汽通量对VPD的响应更为敏感。此外, Ttot对VPD的响应水平明显高于ET, 且该现象在降雨日更为明显。

|
| 图 7 降雨和非降雨日的蒸腾和蒸散发与主要环境因子的相关性 Fig. 7 Reponses of transpiration (T) and evapo-transpiration (ET) to the vapor pressure deficit (VPD) and net radiation (Rn) under different precipitation conditions R:降雨日 Rainy days, NR:非降雨日 Non-rainy days;图中方程均显著(P<0.05), 降雨日的n=288, 非降雨日的n=480 |
降雨和非降雨日Ttot与Rn均呈正相关关系(图 7), 且非降雨日时的R2较高;其中, 降雨日Ttot的回归方程斜率较非降雨日低, 说明非降雨日的Ttot随Rn的变化敏感度略强。ET对Rn的响应与Ttot较为相似(图 7), 其在非降雨日的方程斜率和R2均较降雨日高;同时, 降雨日Rn数值变化范围较小, 而ET的离散程度较非降雨日高。此外, 降雨日Ttot与Rn的响应水平较高时, ET的响应水平却较低, 但非降雨日则不存在该现象。上述结果表明, ET会随着水分输入状况对水汽通量组分进行调节, Rn对生态系统水分平衡的影响更为明显。
4 讨论EC系统的能量闭合状况直接影响到对ET的评估效果, 与FLUXNET相比(R2范围0.64—0.95)[36], 该研究站点的能量闭合水平(R2为0.77) 在可靠范围内, 可保证较为理想的数据质量。同时, EC技术容易低估夜间湍流不充分时的通量[24], 该研究通过对u*的敏感性测试剔除了湍流强度较弱时的数据[32-33], 保证EC系统的测定是处于强湍流条件下的结果, 并采用基于Penman-Monteith模型[35]的BP神经网络对缺失数据进行插补, 拟合结果的R2在0.80以上, 保证数据的完整性和连续性。整个观测期间的Rn总收入占该区域年总收入的16.6%(图 2), 约为观测天数(32 d)与全年天数(365 d)之比(8.8%)的两倍, 主要与该研究区域的地理位置特点有关[37]。不区分降雨日时的兴安落叶松林蒸腾能力为1.28 mm/d, 略高于张彦群等[38]总结的北方针叶林生长季1.24 mm/d的蒸腾能力, 表明大兴安岭北部的兴安落叶松天然林的蒸腾耗水能力较强, 对于区域水量平衡具有重要意义, 同时, 由于VPD和Rn是林木蒸腾耗水的主要驱动力[16-17, 19], 环境因子的年际差异以及微生境差异会影响观测结果。降雨日和非降雨日的Td均明显高于Ti和Ts(图 3), 与优势木处于林内竞争优势地位有关, 其冠层位于林分平均冠层之上[31], 可充分获取太阳辐射能量以驱动树干液流[17], 对环境因子的响应也相应的敏感[21]。
对于兴安落叶松林生态系统, 降雨日的辐射能量相对非降雨日更多的分配至LE, 表现为降雨日的LE/Rn略高, 因而降雨日时代表Ta和Ts增温效应的能量组分H、G比例相应减少, 表明该时间段的生态系统会维持相应的ET需求, 保证生态系统水分平衡[15];然而, 非降雨日的ET明显更高(图 4), 表明该时期的环境条件更有利于植物-大气界面的水汽通量交换;同时, 结果显示降雨日ET主要以E占优势, 而非降雨日则以Td为主, 与以往的研究结果相符[14-15], 说明生态系统会随着水分条件的改变对ET组分(E和T所占比例)进行动态调节[15], 主要原因是叶片气孔对空气湿度增加的响应较为强烈从而限制了植物的蒸腾耗水[18-19]。不区分降雨日时的Ttot约占ET的54.9%(图 5), 低于Schlesinger等[5]对北方针叶林的统计结果(平均为65%), 主要原因是该研究观测期间的降雨事件较多, 不利于维持林木的蒸腾需求[19], 但对E的影响相对较弱[15, 18-19]。
值得注意的是, 尽管以往研究多以林分Td代表林木平均蒸腾能力进行尺度上推[20-21], 但主要原因是中等木和劣势木的边材宽度较小, 利用SF法进行观测时的不确定性较大;该研究通过生长芯法的实际测量结果, 将液流探针切向插入中等木和劣势木边材[9], 保证SF法的观测精度;而结果表明不考虑降雨时Ti与Ts之和仅占ET的20%, 因而若仅以优势木的蒸腾耗水状况进行尺度上推容易高估林分的蒸腾能力。在研究Ttot和ET对主要环境因子的响应关系时, 需要考虑水分条件的差异[5, 15-16]。Ttot对VPD的响应水平明显高于ET, 该现象在降雨日更为明显, 表明无论降雨与否, VPD都是生态系统水汽通量的限制性因子, 而降雨引发的“增湿降温”现象对Ttot的影响更为明显[15];同时, Ttot和ET与Rn的相关性均在非降雨日略高, 与以往研究结果略有不同[15], 可能与研究树种的木质部结构、树干储水性能和树形结构等因素有关[39], ;相对于VPD, Rn与水汽通量的相关性更强, 可能与太阳辐射是驱动生态系统水分循环的首要条件有关[17]。
5 结论降雨和非降雨日各分化等级兴安落叶松Fs的日变化均呈单峰格局;Td明显高于Ti和Ts, 且该现象在非降雨日更为明显。降雨日的ET较低, 而LE/Rn略高, 且连续降雨期间ET的日间变化略明显, 表明非降雨日环境条件更有利于植物-大气界面水汽交换。同时, 各蒸腾组分与ET之比在降雨日较低, 说明降雨日的ET以自由水蒸发为主, 而非降雨日时则以Td占优;不区分降雨日时, Ti与Ts之和约占ET的20%, 因而仅以优势木的蒸腾耗水状况进行尺度上推会高估林分的蒸腾能力。总体上, 降雨和非降雨日Ttot与VPD、Rn的相关性均较ET高, 且Rn与Ttot、ET的相关性均较VPD高, 表明Ttot对环境因子的响应略敏感, 而Rn是驱动生态系统水汽通量的首要条件。该研究为进一步了解大兴安岭北部寒温带兴安落叶松天然林的水汽通量特征提供了科学依据和数据支持。
| [1] | Xiao J F, Sun G, Chen J Q, Chen H, Chen S P, Dong G, Gao S H, Guo H Q, Guo J X, Han S J, Kato T, Li Y L, Lin GH, Lu W Z, Ma M G, McNulty S, Shao C L, Wang X F, Xie X, Zhang X D, Zhang Z Q, Zhao B, Zhou G S, Zhou J. Carbon fluxes, evapotranspiration, and water use efficiency of terrestrial ecosystems in China. Agricultural and Forest Meteorology, 2013, 182-183: 76–90. DOI:10.1016/j.agrformet.2013.08.007 |
| [2] | Jasechko S, Sharp Z D, Gibson J J, Birks S J, Yi Y, Fawcett P J. Terrestrial water fluxes dominated by transpiration. Nature, 2013, 496(445): 347–359. |
| [3] | Avissar R, Schmidt T. An evaluation of the scale at which ground-surface heat flux patchiness affects the convective boundary layer using large-eddy simulations. Journal of the Atmospheric Sciences, 1998, 55(16): 2666–2689. DOI:10.1175/1520-0469(1998)055<2666:AEOTSA>2.0.CO;2 |
| [4] | Zhang B Z, Xu D, Liu Y, Li F S, Cai J B, Du L J. Multi-scale evapotranspiration of summer maize and the controlling meteorological factors in north China. Agricultural and Forest Meteorology, 2016, 216: 1–12. DOI:10.1016/j.agrformet.2015.09.015 |
| [5] | Schlesinger W H, Jasechko S. Transpiration in the global water cycle. Agricultural and Forest Meteorology, 2014, 189-190: 115–117. DOI:10.1016/j.agrformet.2014.01.011 |
| [6] | Fuentes J D, Barr J G. Mangrove forests and carbon and water cycling. Agricultural and Forest Meteorology, 2015, 213: 263–265. DOI:10.1016/j.agrformet.2015.08.243 |
| [7] | 孙慧珍, 周晓峰, 康绍忠. 应用热技术研究树干液流进展. 应用生态学报, 2004, 15(6): 1074–1078. |
| [8] | Wilson K B, Hanson P J, Mulholland P J, Baldocchi D D, Wullschleger S D. A comparison of methods for determining forest evapotranspiration and its components:sap-flow, soil water budget, eddy covariance and catchment water balance. Agricultural and Forest Meteorology, 2001, 106(2): 153–168. DOI:10.1016/S0168-1923(00)00199-4 |
| [9] | Lu P, Urban L, Zhao P. Granier's thermal dissipation probe (TDP) method for measuring sap flow in trees:theory and practice. Acta Botanica Sinica, 2004, 46(6): 631–646. |
| [10] | Vertessy R A, Benyon R G, O'Sullivan S K, Gribben P R. Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiology, 1995, 15(9): 559–567. DOI:10.1093/treephys/15.9.559 |
| [11] | Wullschleger S D, King A W. Radial variation in sap velocity as a function of stem diameter and sapwood thickness in yellow-poplar trees. Tree Physiology, 2000, 20(8): 511–518. DOI:10.1093/treephys/20.8.511 |
| [12] | Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems:past, present and future. Global Change Biology, 2003, 9(4): 479–492. DOI:10.1046/j.1365-2486.2003.00629.x |
| [13] | Wofsy S C, Goulden M L, Munger J W, Fan S M, Bakwin P S, Daube B C, Bassow S L, Bazzaz F A. Net exchange of CO2 in a mid-latitude forest. Science, 1993, 260(5112): 1314–1317. DOI:10.1126/science.260.5112.1314 |
| [14] | 白岩, 朱高峰, 张琨, 马婷. 基于树干液流及涡动相关技术的葡萄冠层蒸腾及蒸散发特征研究. 生态学报, 2015, 35(23): 7821–7831. |
| [15] | 刘晨峰, 张志强, 孙阁, 查同刚, 朱金兆, 申李华, 陈军, 方显瑞, 陈吉泉. 基于涡度相关法和树干液流法评价杨树人工林生态系统蒸发散及其环境响应. 植物生态学报, 2009, 33(4): 706–718. |
| [16] | 徐世琴, 吉喜斌, 金博文. 典型固沙植物梭梭生长季蒸腾变化及其对环境因子的响应. 植物生态学报, 2015, 39(9): 890–900. DOI:10.17521/cjpe.2015.0085 |
| [17] | O'Brien J J, Oberbauer S F, Clark D B. Whole tree xylem sap flow responses to multiple environmental variables in a wet tropical forest. Plant, Cell and Environment, 2004, 27(5): 551–567. DOI:10.1111/pce.2004.27.issue-5 |
| [18] | Whitehead D. Regulation of stomatal conductance and transpiration in forest canopies. Tree Physiology, 1998, 18(8/9): 633–644. |
| [19] | Meinzer F C, Goldstein G, Jackson P, Holbrook N M, Gutiérrez M V, Cavelier J. Environmental and Physiological regulation of transpiration in tropical forest gap species:the influence of boundary layer and hydraulic properties. Oecologia, 1995, 101(4): 514–522. DOI:10.1007/BF00329432 |
| [20] | 孙慧珍, 孙龙, 王传宽, 周晓峰. 东北东部山区主要树种树干液流研究. 林业科学, 2005, 41(3): 36–42. DOI:10.11707/j.1001-7488.20050306 |
| [21] | 李振华, 王彦辉, 于澎涛, 王艳兵, 韩新生, 曹恭祥. 华北落叶松液流速率的优势度差异及其对林分蒸腾估计的影响. 林业科学研究, 2015, 28(1): 8–16. |
| [22] | 赵平, 曾小平, 余作岳. 华南丘陵人工马占相思林下灌木湿润季节蒸腾与小气候的关系. 应用生态学报, 1997, 8(4): 365–371. |
| [23] | 赵平, 饶兴权, 马玲, 蔡锡安, 曾小平. 马占相思(Acacia mangium)树干液流密度和整树蒸腾的个体差异. 生态学报, 2006, 26(12): 4050–4058. DOI:10.3321/j.issn:1000-0933.2006.12.018 |
| [24] | Fisher J B, Baldocchi D D, Misson L, Dawson T E, Goldstein A H. What the towers don't see at night:nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California. Tree Physiology, 2007, 27(4): 597–610. DOI:10.1093/treephys/27.4.597 |
| [25] | 胡海清, 罗碧珍, 魏书精, 魏书威, 文正敏, 孙龙, 罗斯生, 王立明, 马洪斌. 大兴安岭5种典型林型森林生物碳储量. 生态学报, 2015, 35(17): 5745–5760. |
| [26] | 玉宝, 乌吉斯古楞, 王百田, 王立明. 大兴安岭兴安落叶松(Larix gmelinii)天然林分级木转换特征. 生态学报, 2008, 28(11): 5750–5757. DOI:10.3321/j.issn:1000-0933.2008.11.062 |
| [27] | 玉宝, 乌吉斯古楞, 王百田, 王立明. 兴安落叶松天然林树冠生长特性分析. 林业科学, 2010, 46(5): 41–48. DOI:10.11707/j.1001-7488.20100507 |
| [28] | Granier A, Biron P, Lemoine D. Water balance, transpiration and canopy conductance in two beech stands. Agricultural and Forest Meteorology, 2000, 100(4): 291–308. DOI:10.1016/S0168-1923(99)00151-3 |
| [29] | James S A, Clearwater M J, Meinzer F C, Goldstein G. Heat dissipation sensors of variable length for the measurement of sap flow in trees with deep sapwood. Tree Physiology, 2002, 22(4): 277–283. DOI:10.1093/treephys/22.4.277 |
| [30] | 金鹰, 王传宽, 桑英. 三种温带树种树干储存水对蒸腾的贡献. 植物生态学报, 2011, 35(12): 1310–1317. |
| [31] | 刘家霖, 王传宽, 张全智. 不同分化等级兴安落叶松树干心材和边材的空间变异. 林业科学, 2014, 50(12): 114–121. |
| [32] | 徐自为, 刘绍民, 宫丽娟, 王介民, 李小文. 涡动相关仪观测数据的处理与质量评价研究. 地球科学进展, 2008, 23(4): 357–370. |
| [33] | Goulden M L, Munger J W, Fan S M, Daube B C, Wofsy S C. Measurements of carbon sequestration by long-term eddy covariance:methods and a critical evaluation of accuracy. Global Change Biology, 1996, 2(3): 169–182. DOI:10.1111/gcb.1996.2.issue-3 |
| [34] | Kljun N, Calanca P, Rotach M W, Schmid H P. A simple parameterisation for flux footprint predictions. Boundary-Layer Meteorology, 2004, 112(3): 503–523. DOI:10.1023/B:BOUN.0000030653.71031.96 |
| [35] | Monteith J L. Evaporation and environment. Symposia of the Society for Experimental Biology, 1965, 19: 205–234. |
| [36] | Wilson K, Goldstein A, Falge E, Aubinet M, Baldocchi D, Berbigier P, Bernhofer C, Ceulemans R, Dolman H, Field C, Grelle A, Ibrom A, Law B E, Kowalski A, Meyers T, Moncrieff J, Monson R, Oechel W, Tenhunen J, Valentini R, Verma S. Energy balance closure at FLUXNET sites. Agricultural and Forest Meteorology, 2002, 113(1/4): 223–243. |
| [37] | 谭正洪, 于贵瑞, 周国逸, 韩士杰, 夏禹九, 前田高尚, 小杉绿子, 山野井克己, 李胜功, 太田岳史, 平田竜一, 安田幸生, 中野隆志, 小南裕志, 北村兼三, 溝口康子, 廖志勇, 赵俊福, 杨廉雁. 亚洲东部森林的小气候特征:1.辐射和能量的平衡. 植物生态学报, 2015, 39(6): 541–553. DOI:10.17521/cjpe.2015.0052 |
| [38] | 张彦群, 王传宽. 北方和温带森林生态系统的蒸腾耗水. 应用与环境生物学报, 2008, 14(6): 838–845. |
| [39] | Phillips N G, Ryan M G, Bond B J, Mcdowell N G, Hinckley T M, Čermák J. Reliance on stored water increases with tree size in three species in the Pacific Northwest. Tree Physiology, 2003, 23(4): 237–245. DOI:10.1093/treephys/23.4.237 |