 2017, Vol. 37
2017, Vol. 37文章信息
- 李群, 赵成章, 王继伟, 赵连春, 徐婷, 韩玲
- LI Qun, ZHAO Chengzhang, WANG Jiwei, ZHAO Lianchun, XU Ting, HAN Ling.
- 张掖湿地芦苇比叶面积和水分利用效率的关系
- Relationship analysis between specific leaf area and water use efficiency of Phragmites australis in the Zhangye wetland
- 生态学报. 2017, 37(15): 4956-4962
- Acta Ecologica Sinica. 2017, 37(15): 4956-4962
- http://dx.doi.org/10.5846/stxb201604280813
-
文章历史
- 收稿日期: 2016-04-28
- 网络出版日期: 2017-03-22

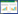




叶片作为植物的基本结构和功能单位, 是植物进行光合作用的主要器官[1], 其功能性状不仅能够反映植物对异质性生境的适应能力, 而且体现了植物与环境协同进化的生活史对策。叶片是土壤、植物与大气间物质转化和能量循环的重要载体[2], 其形态和结构决定了植物获取光照、水分和养分等限制性资源的能力, 也影响着植物的净光合速率和水分利用效率[1], 因此, 叶片功能性状及其资源配置模式, 是认识植物环境可塑性机制和光合碳同化途径的基础。叶片作为植物体暴露在环境中的表面积最大的器官, 对各种胁迫因子具有较强的感知能力, 而且能够通过叶面积、叶厚度、比叶面积等功能性状在形态和结构上的可塑性[3-4], 差异化地调控植物的生理生态功能, 间接影响植物的光合能力和营养物质的吸收水平[2]。比叶面积(SLA)是指叶面积与其干重之比(或者其倒数比叶质量), 可反映植物响应资源约束水平的光合碳积累策略[5], 与植物的光拦截效率、气孔导度密切相关[6], 能够影响植物的蒸腾速率和水分利用效率[7]。有研究表明, 不同大小SLA的叶片, 可能通过调整叶片厚度、光合组织密度等结构, 优化水分的蒸腾速率和固碳能力[8-9], 形成与群落光照环境、土壤水分供给水平相一致的资源吸收、传输和利用能力[10]。水分利用效率(WUE)是耦合植物叶片光合作用和蒸腾作用的重要指标[11], 综合反映了植物对各种胁迫因子的水资源有效利用能力, 与叶片的蒸腾速率和净光合速率密切相关。密度是自然界植物种群普遍存在的重要选择压力之一[12], 密度的变化能够改变植株可获得性资源的数量, 使植物所需空间、光照和土壤水分等生存资源可能受到邻株的物理或几何上的拥挤效应, 将促使植物调整叶片生物量分配策略, 形成与光能获取需求相适应的比叶面积;在叶片生物量有限性约束下, 植物可能选择差异化的叶片保卫结构、叶片厚度构建模式, 从而形成了与环境相适应的蒸腾速率、净光合速率和水分利用效率。因此, 比叶面积与水分利用效率的关系研究, 对于认识植物叶片功能性状与光合效率之间的协同变异规律具有重要的理论意义。
芦苇(Phragmites australis)属于多年生根茎型禾本科植物, 具有极强的环境适应能力, 广泛分布于内陆草甸区和近海河口湿地区, 也是西北荒漠区沼泽湿地生态系统的优势植物。目前, 学术界围绕植物比叶面积的生境适应性[9, 13]、植物叶片光合参数与环境因子的关系[1]、植物叶片水分利用效率[7, 14], 以及湿地植物叶片功能性状等问题展开了广泛研究[15], 有关芦苇的生境胁迫适应机制[16]、光合碳同化途径[17]、多时空尺度蒸散模拟研究[18]等也得到了重视, 但是关于密度制约下湿地植物叶片性状的空间变异规律和叶片水分利用效率的变化关系尚不明晰。鉴于此, 本研究以张掖国家级湿地公园沼泽湿地的芦苇为研究对象, 通过分析密度影响下芦苇比叶面积与水分利用效率的关系, 试图明晰:(1) 不同密度梯度芦苇比叶面积与水分利用效率存在何种关系?(2) 形成这种生长关系的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解。
1 研究地区与研究方法 1.1 研究区域和样地概况研究区位于甘肃省张掖国家湿地公园保育区(图 1), 地理位置为100°26′50.72″E, 38°58′42.9″N, 海拔1462.7 m, 属温带大陆性气候, 年平均气温7.0 ℃, ≥0℃积温为2734 ℃, 年均降水量129 mm, 主要集中在6-9月, 年均蒸发量2047 mm, 年日照时数3085 h。由于地表经常过湿或有薄层积水, 受常年或季节性水淹的影响, 形成以沼泽土为主的自然土壤, 发育了水生和湿生植物群落, 以湿生和挺水植物为主。主要植物有:芦苇、菖蒲(AcoruscalamusL.)、香蒲(Typhaorientalis)、假苇拂子茅(Calamagrostis pseudophragmites)、千屈菜(Lythrum salicaria)、水葱(Scirpus validus)、黑三棱(Sparganium stoloniferum)等。该研究区位于湿地公园保育区, 四周建有围栏, 不存在家畜放牧和人类活动等干扰因素, 能够反映湿地的自然属性。
1.2 实验方法在2015年8月, 通过大量实地考察, 选择一块从湖边向湖心岛中部平缓延伸、面积3 hm2, 芦苇为单一优势种的沼泽湿地, 随着地势的微弱抬升, 土壤含水量和植被覆盖度发生了相应的梯度性变化。实验以芦苇分株为取样单位进行实验, 按芦苇分株的密度状况, 将芦苇种群的密度水平分为3个面积为20 m×20 m的梯度样地(图 1), 分别为:高密度(Ⅰ, 210-230株/m2)、中密度(Ⅱ, 130-150株/m2)和低密度(Ⅲ, 50-70株/m2)。按已划分的芦苇种群的密度梯度样地中按z字形从下到上的顺序依次设置6个2 m×2 m样方, 总计18个样方。首先进行群落土壤水分测定, 在每个样方内间隔10 cm用土钻(直径=2 cm)分5层采集0-50 cm土样, 之后将土样混合, 重复3次, 所有样品均剔除明显的植物根段和凋落物等杂质, 现场称鲜质量后, 装入编号的铝盒中, 带回实验室, 在105 ℃的烘箱内烘12 h, 取出称重, 计算土壤含水量。然后在每一个样方随机选择12株芦苇(株高为大中小各4株), 先用卷尺测量从基部到最高分枝的高度, 定为株高, 然后选取每个植株最外层4个方位充分伸展且健康完整的2-3个叶片, 用于芦苇叶片光合特征和叶性状的调查:

|
| 图 1 研究区概况和样地设置图 Fig. 1 Study area and locations of measured plots |
(1) 光合特征值测定:选择晴朗天气的9:00-12:00对标记过的芦苇植株进行植物气体交换参数的测定, 光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(PAR)为1200 μmol m-2s-1, CO2浓度为340 μmol/mol, 相对湿度(RH)保持在50%-60%, 流速设定为750 μmol/s。选取每个植株中端展开的4片成熟健康叶片分别测定叶片的净光合速率(Pn)、蒸腾速率(Tr)等参数, 待数据稳定后读取数据, 计算WUE(Pn/Tr)。
(2) 比叶面积测定:将测定光合作用的叶片装入信封迅速带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)扫描叶片, 利用Image软件计算扫描的叶面积, 最后将叶片装入信封, 于80 ℃烘箱中烘干48 h, 称量(精度0.0001 g)。比叶面积用叶面积和叶干重的比值表示。
(3) 叶厚度测定:使用游标卡尺测量(精确到0.01 mm), 测量时避开叶脉, 并尽量选择叶片中心部位, 每个个体在不同部位测量5次, 最后取平均值为该个体的叶片厚度。
1.3 数据处理不同密度样地湿地群落生物学特征和芦苇种群的各功能性状的所有实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析。利用线性回归的方法对不同密度条件下芦苇比叶面积和WUE的关系进行研究, 对不同密度样地植物功能性状平均值的差异比较采用单因素方差分析(one-way ANOVA), 显著性水平设为0.05。实验数据分析采用SPSS 16.0软件进行, 用SigmaPlot 10.0和Excel软件绘图。
2 结果与分析 2.1 不同密度条件下芦苇种群的生物学特性和土壤含水量的变化单因素方差分析表明, 不同密度条件下芦苇的各个叶功能性状和土壤含水量存在显著差异(P < 0.05; 表 1)。随着芦苇种群密度的逐渐降低, 0-50 cm土壤含水量呈逐渐减小趋势, 从高密度(Ⅰ)到低密度(Ⅲ)显著减小了31.00%;芦苇的株高和蒸腾速率(Tr)呈逐渐减小的趋势, 从高密度(Ⅰ)到低密度(Ⅲ), 分别减小了41.77%和59.35%;芦苇的叶厚度和净光合速率(Pn)呈逐渐增大的趋势, 由高密度(Ⅰ)到低密度(Ⅲ)分别增加了42.11%和49.28%;中密度样地(Ⅱ)是芦苇生物学特性和土壤含水量从最大值到最小值的过渡区。
| 密度 Density | 土壤含水量 Soil moisture/% | 株高 Height/cm | 叶厚度 Leaf thick ness/mm | 净光合速率 Pn/(μmol CO2 m-2s-1) | 蒸腾速率 Tr/(mmol H2O m-2s-1) |
| 高密度 High density | 67.50±3.38a | 376.67±10.24a | 0.19±0.01c | 4.83±0.24c | 1.23±0.06a |
| 中密度 Medium density | 58.77±2.94b | 251.67±8.24b | 0.22±0.01b | 6.72±0.13b | 0.57±0.03b |
| 低密度 Low density | 46.57±2.33c | 219.33±5.24c | 0.27±0.02a | 7.21±0.36a | 0.50±0.03c |
| 同列不同小写字母表示在密度间差异显著(P < 0.05); | |||||
单因素方差分析表明, 不同密度条件下芦苇种群的水分利用效率和叶性状均存在显著差异(P < 0.05, 图 2)。随着种群密度的逐渐降低, 芦苇叶面积、叶干重和比叶面积(SLA)呈逐渐减小趋势, 水分利用效率(WUE)呈逐渐增大趋势, 从高密度(Ⅰ)到低密度(Ⅲ)样地, 叶面积减少了35.41 cm2、叶干重减少了12.21 g、比叶面积减少了1.23 cm2/g, 水分利用效率增加了7.54 μmol CO2/mmol H2O, 从最大值到最小值分别减少了84.19%、65.22%和53%, 水分利用效率(WUE)显著增加了2.69倍;中密度样地(Ⅱ)是芦苇叶性状和水分利用效率从最大值到最小值的过渡区。表明不同密度条件下芦苇叶面积大于叶干重的减小速度, 比叶面积和水分利用效率呈现相反的变化趋势。

|
| 图 2 不同密度条件下芦苇种群叶性状和水分利用效率(数值为平均值±标准误差) Fig. 2 Leaf traits and water use efficiency of Phragmites australis in different densities (mean±SE) 同列不同小写字母表示在密度间差异显著(P < 0.05) |
不同密度生境对植物叶片形态和功能的选择性塑造, 能够促使植物形成与生境高度适应的SLA, 并优化植物的净光合速率和水分利用效率。不同密度条件下芦苇比叶面积(SLA)与水分利用效率(WUE)的关系存在显著差异(P < 0.05, 图 3)。Pearson相关性分析表明, 芦苇叶片SLA与WUE在高密度(Ⅰ)和低密度(Ⅲ)均呈极显著负相关关系(yⅠ=-0.176x + 0.652, R2=0.814, P < 0.01; yⅢ=-4.912x + 1.202, R2=0.808, P < 0.01, 图 3), 在中密度样地(Ⅱ)样地, 二者呈显著负相关关系(yⅡ=-3.448x + 1.948, R2=0.587, P < 0.05)。表明随着芦苇种群密度逐渐降低, 芦苇的WUE逐渐增大, SLA逐渐减小, 二者呈显著的负相关关系, 高密度和低密度生境中芦苇植株的比叶面积和水分利用效率呈显著的权衡关系, 中密度样地是芦苇比叶面积逐渐减小和水分利用效率逐渐增加的过渡区域。

|
| 图 3 不同密度梯度下芦苇比叶面积(SLA)与水分利用效率(WUE)之间关系 Fig. 3 Relationship between specific leaf area and water use efficiency of Phragmites australis among different densities |
植物可获得性资源的有限性通常与邻株植物的物理或几何上的拥挤效应有关, 能够改变植物可获得性资源的权衡分配、组合和自我弥补[10], 迫使植株通过调整叶面积等功能性状的资源配置和叶片构型构建模式[3], 形成与生境高度适应的比叶面积, 从而改变了叶片保卫结构、叶肉细胞的组织形式[13], 提高了异质生境中植物的水分利用效率。本研究发现, 随着湿地群落芦苇种群密度的逐渐降低, 芦苇的比叶面积逐渐减小, 水分利用效率逐步增加;在高密度样地(Ⅰ)和低密度样地(Ⅲ)芦苇的比叶面积与WUE之间存在极显著负相关关系(P < 0.01), 在中密度样地二者呈显著负相关关系(P < 0.05)。湿地群落中不同密度样地中芦苇种群比叶面积和WUE关系的差异性, 既是密度制约对叶片形态和功能的选择性塑造, 也是邻体干扰下植物权衡碳投入-收益的必然结果。
3.1 高密度样地芦苇比叶面积与WUE的关系低光照条件下植物种群为了捕捉更多的光能会倾向用有限的营养物质创造出更大的受光面积, 以最大限度的获取光照和控制叶片内水分散失的平衡[5]。相邻植株个体的构件之间对光、水分、营养和空间等资源的争夺, 导致处于竞争劣势的个体构件生长率下降、生物量减小[19]。在湿地群落中芦苇种群的高密度样地(Ⅰ)中, 芦苇具有最大的株高和盖度(表 1), 种群内部的拥挤度和隐蔽度较大, 茎叶占有的生存空间相对狭窄, 光照条件成为影响植物叶片生理生态过程的主要因素。在强烈的邻体干扰和光线竞争激烈的环境中, 芦苇将更多的生物量投资于株高, 可以提高叶片配置的机动性, 从而更有利于捕获未被充分利用的闲置空间, 在叶片生物量有限的状态下, 为了保证少量干物质投资下捕光表面积数量和光合收益的最大化, 并且能够降低自身的机械重量[20], 芦苇倾向大而薄的叶片构型模式(表 1、图 2), 以保证最大限度地满足低光条件下芦苇叶片捕捉更多光能的需求, 进而形成了较大的SLA(图 2);生长在高密度样地的芦苇拥有高的SLA意味着高的叶周转[21], 而且较小叶厚度拉近了气孔与叶片表面的距离, 减小了叶片光合作用中的水分输送阻力, 又由于该生境土壤含水量较高(表 1), 不存在干旱胁迫的压力, 环境选择偏利于具有较大水分蒸腾能力的植物, 因此芦苇形成了较高的蒸腾速率(表 1), 从而增大了叶片的呼吸能耗, 影响了植物的净光合速率(表 1), 降低了叶片的水分利用效率(图 2), 导致比叶面积和WUE间形成了极显著负相关关系(P < 0.01, 图 3)。与West GB[22]等提出的“自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化”的观点一致。
3.2 低密度样地芦苇比叶面积与WUE的关系最优化分配理论预测, 在无邻体植物竞争时, 植物应该增大获取限制资源的器官分配, 而减少获取非限制资源的器官分配[23], 即在特定的环境下, 植物为了满足生存需求会形成不同的形态结构和特定的水分供应策略[24]。在低密度样地(Ⅲ), 芦苇种群的盖度和株高最小(表 1), 在实验过程中发现芦苇株丛间的无明显的干扰和遮光现象, 土壤含水量较低(表 1), 芦苇面临光照充分和水资源相对不足的限制条件, 若将有限资源投入到叶面积上不但延缓了叶片的散热时间, 还加重了植株的蒸腾失水, 使其处于不利的竞争态势[16], 因而, 生长在低密度湿地的芦苇选择了小而厚的叶片, 伴随着比叶面积的降低(表 1、图 2), 较低的比叶面积是植物长期适应干旱环境的一种表型选择[5], 芦苇将叶片干物质更多地用于构建保卫结构、叶肉细胞密度和叶片厚度, 增加了水分运输路径的阻力[25], 从而显著降低了蒸腾速率(表 1), 在减少了单位叶面积的呼吸碳损失情形下, 获得了较高的净光合速率, 进而保证了最大的水分利用效率(图 2)。小叶具有叶边缘进行热量和物质交换的阻力更低的特性, 因此, 生长在低密度样地的芦苇依靠小叶获得了水分运输方面的选择优势, 实现了水分利用效率的最大化, 从而形成了比叶面积与WUE的极显著负相关关系(P < 0.01, 图 3)。为了满足自身生长需要和在复杂多变的应力生长环境下生存, 芦苇经过长时间的进化, 叶片的结构布局已经达到了一种自然有序的最优结构。
3.3 中密度芦苇比叶面积与WUE的关系植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争[19], 植物可以通过改变不同器官间生物量分配的可塑性反应进行表型调整[26]。在中密度样地(Ⅱ), 芦苇的高度和盖度较样地(Ⅰ)显著下降(表 1), 芦苇株丛间的干扰强度减弱、土壤水分条件和太阳辐射基本能够满足植物的生存, 若仍生长较大叶面积有可能导致呼吸和蒸腾成本更高, 不利于其光合碳获取能力最大化, 为了增强生境适合度芦苇选择了适度的叶面积和叶厚度, 在蒸腾速率降低的条件下, 获得了较高的净光合速率(表 1), 实现了水分利用效率的最大化。因此, 生长在中密度(Ⅱ)样地的芦苇的比叶面积和WUE的相关性仅达到显著水平(P < 0.05, 图 3), 是比叶面积由大到小, WUE由小到大转变的区域, 这是由于光合产物在不同构件或功能间权衡分配的结果。
4 结论植物比叶面积与WUE的关系是植物生物量分配格局的环境适应性体现, 也反映了植物叶形态与其光合产物在水分利用效率上的权衡关系[9]。高密度湿地的芦苇选择了大而薄的叶片, 形成了较高比叶面积和低水分利用效率的资源利用策略;低密度湿地的芦苇选择了小而厚的叶片, 趋向于低比叶面积和高水分利用效率的光合产物积累模式。这种随着环境变化的感应机制和“趋利避害”的表型可塑性是湿地植物与环境长期相互协同进化和适应的结果, 可以提高密度制约下植物的生存、生长和繁殖适合度, 也是顺利完成生活史的最佳生理和生态功能状态。本文仅从密度角度分析了湿地植物芦苇比叶面积与水分利用效率的关系, 它们的关系可能受土壤含水量、土壤养分及其它环境因子的影响, 还需实验验证。
| [1] | Knight C A, Ackerly D D. Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size:Congeneric species from desert and coastal environments. New Phytologist, 2003, 160(2): 337–347. DOI:10.1046/j.1469-8137.2003.00880.x |
| [2] | Deng J M, Li T, Wang G X, Liu J, Yu Z L, Zhao C M, Ji M F, Zhang Q. Trade-offs between the metabolic rate and population density of plants. PLoS ONE, 2008, 3(3): e1799. DOI:10.1371/journal.pone.0001799 |
| [3] | Westoby M, Wright I J. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species. Oecologia, 2003, 135(4): 621–628. DOI:10.1007/s00442-003-1231-6 |
| [4] | 施宇, 温仲明, 龚时慧, 宋光, 郑颖, 丁曼. 黄土丘陵区植物功能性状沿气候梯度的变化规律. 水土保持研究, 2012, 19(1): 107, 116–111, 116. |
| [5] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [6] | Milla R, Reich P B. The scaling of leaf area and mass:The cost of light interception increases with leaf size. Proceedings of the Royal Society B:Biological Sciences, 2007, 274(1622): 2109–2115. DOI:10.1098/rspb.2007.0417 |
| [7] | Wright I J, Westoby M, Reich P B. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. Journal of Ecology, 2002, 90(3): 534–543. DOI:10.1046/j.1365-2745.2002.00689.x |
| [8] | Warren C R, Tausz M, Adams M A. Does rainfall explain variation in leaf morphology and physiology among populations of red ironbark (Eucalyptus sideroxylon subsp. tricarpa) grown in a common garden?. Tree Physiology, 2005, 25(11): 1369–1378. |
| [9] | Gago J, Douthe C, Florez-Sarasa I, Escalona J M, Galmes J, Fernie A R, Flexas J, Medrano H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Science, 2014, 226: 108–119. DOI:10.1016/j.plantsci.2014.04.007 |
| [10] | Kenney A M, McKay J K, Richards J H, Juenger T E. Direct and indirect selection on flowering time, water-use efficiency (WUE, δ 13C), and WUE plasticity to drought in Arabidopsis thaliana. Ecology and Evolution, 2014, 4(23): 4505–4521. DOI:10.1002/ece3.2014.4.issue-23 |
| [11] | Farquhar G D, Richards R A, Juenger T E. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Australian Journal of Plant Physiology, 1984, 11(6): 539–552. DOI:10.1071/PP9840539 |
| [12] | Japhet W, Zhou D W, Zhang H X, Zhang H X, Yu T. Evidence of phenotypic plasticity in the response of Fagopyrum esculentum to population density and sowing date. Journal of Plant Biology, 2009, 52(4): 303–311. DOI:10.1007/s12374-009-9037-7 |
| [13] | 胡耀升, 么旭阳, 刘艳红. 长白山森林不同演替阶段比叶面积及其影响因子. 生态学报, 2015, 35(5): 1480–1487. |
| [14] | 全先奎, 王传宽. 帽儿山17个种源落叶松针叶的水分利用效率比较. 植物生态学报, 2015, 39(4): 352–361. DOI:10.17521/cjpe.2015.0034 |
| [15] | 党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚. 祁连山高寒草地甘肃臭草叶性状与坡向间的关系. 植物生态学报, 2015, 39(1): 23–31. DOI:10.17521/cjpe.2015.0003 |
| [16] | 庄瑶, 孙一香, 王中生, 杨琳璐, 邓自发, 姚志刚, 安树青. 芦苇生态型研究进展. 生态学报, 2010, 30(8): 2173–2181. |
| [17] | 郑学平, 张承烈, 陈国仓. 河西走廊芦苇的光合碳同化途径对生境条件的适应. 植物生态学与地植物学学报, 1993, 17(1): 1–8. |
| [18] | 于文颖, 纪瑞鹏, 冯锐, 武晋雯, 赵先丽, 张玉书. 芦苇湿地多时空尺度蒸散模拟研究进展. 生态学杂志, 2014, 33(5): 1388–1394. |
| [19] | 黎磊, 周道玮. 红葱种群地上和地下构件的密度制约调节. 植物生态学报, 2011, 35(3): 284–293. |
| [20] | 罗震, 陈立平, 黄玉盈, 张云清. 连续体结构的拓扑优化设计. 力学进展, 2004, 34(4): 463–476. DOI:10.6052/1000-0992-2004-4-J2003-111 |
| [21] | 薛伟, 李向义, 朱军涛, 林丽莎, 王迎菊. 遮阴对疏叶骆驼刺叶形态和光合参数的影响. 植物生态学报, 2011, 35(1): 82–90. |
| [22] | West G B, Brown J H, Enquist B J. A general model for the origin of allometric scaling laws in biology. Science, 1997, 276(5309): 122–126. DOI:10.1126/science.276.5309.122 |
| [23] | Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4): 207–215. DOI:10.1078/1433-8319-00083 |
| [24] | 龚容, 高琼. 叶片结构的水力学特性对植物生理功能影响的研究进展. 植物生态学报, 2015, 39(3): 300–308. |
| [25] | 祁建, 马克明, 张育新. 北京东灵山不同坡位辽东栎(Quercusliao tungensis)叶属性的比较. 生态学报, 2008, 28(1): 122–128. |
| [26] | Cheplick G P. A modular approach to biomass allocation in an invasive annual (Microstegium vimineum; Poaceae). American Journal of Botany, 2006, 93(4): 539–545. DOI:10.3732/ajb.93.4.539 |
| [27] | 刘明虎, 辛智鸣, 徐军, 孙非, 窦立军, 李永华. 干旱区植物叶片大小对叶表面蒸腾及叶温的影响. 植物生态学报, 2013, 37(5): 436–442. |