 2017, Vol. 37
2017, Vol. 37文章信息
- 马维伟, 王辉, 李广, 石万里, 赵锦梅, 罗永忠, 聂志刚, 薛景文
- MA Weiwei, WANG Hui, LI Guang, SHI Wanli, ZHAO Jinmei, LUO Yongzhong, NIE Zhigang, XUE Jingwen.
- 甘南尕海湿地退化过程中植被生物量变化及其季节动态
- Changes in plant biomass and its seasonal dynamics during degradation succession in the Gahai wetland
- 生态学报. 2017, 37(15): 5091-5101
- Acta Ecologica Sinica. 2017, 37(15): 5091-5101
- http://dx.doi.org/10.5846/stxb201604260793
-
文章历史
- 收稿日期: 2016-04-26
- 网络出版日期: 2017-03-22





2. 兰州大学草地农业生态系统国家重点实验室, 兰州 730020;
3. 甘肃农业大学信息科学技术学院, 兰州 730070;
4. 甘肃则岔自然保护区管理局, 碌曲 747200
2. State Key Laboratory of Grassland Agro-ecosystems, Lanzhou University, Lanzhou 730020, China;
3. College of Information Science and Technology, Gansu Agricultural University, Lanzhou 730070, China;
4. Gahai Zecha Nature Reserve Administration, Luqu 747200, China
作为一种特殊生态系统, 湿地系统有机碳储量、分布及其动态在全球碳循环和气候变化研究计划中受到密切关注[1-2]。其中, 植物生物量作为湿地的重要组成部分, 是湿地生态系统生产力最基本的数量特征, 也是湿地生态系统物质循环和能量流动的一个重要方面, 其微小变化都会引起湿地生态系统结构和功能的改变, 因此研究湿地生物量格局变化, 不仅对了解湿地的健康状况有重要意义, 而且对深入分析湿地生态系统固碳能力与碳循环提供重要科学依[3]。
尕海湿地位于全球气候变化最为敏感的地区——青藏高原的东部, 是我国独有青藏高原湿地重要组成部分, 是维系甘南高原生态安全的重要屏障, 已被列入国际重要湿地名录, 属于国家重点保护湿地。因地处高寒区域、常年冷湿环境使得湿地生态系统积累了大量有机物质, 成为青藏高原自然生态系统最为重要的碳库之一, 对区域碳循环以及大气温室气体平衡有着重要的功能和作用[4]。近年来, 受气候变暖、过度放牧及湿地排水疏干等作用, 尕海湿地出现不同程度退化[5-6]。在湿地退化条件下, 湿地植物生物量将如何响应对于估算本区域湿地碳库潜力十分重要。此外, 高寒湿地生态系统比较脆弱, 一旦遭到破坏, 会引起湿地土壤退化, 成为“黑土滩”, 其退化速度惊人, 且往往是不可逆的, 而恢复却很困难缓慢。因而, 详细了解高寒湿地退化过程中植被生物量变化特征, 有助于对湿地发生退化过程的认识及防止湿地退化的发生。因此, 本文从湿地退化过程中植被生物量动态变化、生物量垂直分布和生物量增长速率等方面, 对尕海高寒湿地退化过程中植被生物量变化规律进行了3a定位研究, 以期为高寒湿地退化的治理与恢复以及湿地生产力的提高, 湿地的可持续利用提供科学依据, 也为准确评估高寒湿地植物碳汇功能及其贡献奠定基础。
1 材料与方法 1.1 研究区概况位于青藏高原东北边缘甘肃省碌曲县尕海-则岔国家级自然保护区内(33°58′12″—34°32′16″N, 102°05′00″—102°05′ 00″—102°47′39″E), 海拔3430—4300 m, 区内湿地面积57846 hm2, 主要为河流湿地、沼泽化草甸和泥炭沼泽, 其中沼泽化草甸51160 hm2, 泥炭沼泽10429 hm2, 泥炭沼泽的泥炭层平均厚度约为1.94 m, 储量较大[6]。在气候分区中被划分为青藏高原带、高寒湿润气候区, 该区气候类型为大陆性季风气候类型区。年均气温1.2 ℃, 无绝对无霜期, 年均降水量781.8 mm, 主要集中在7—9月, 年蒸发量1150.5 mm。土壤类型主要包括暗色草甸土、沼泽土和泥炭土等, 植物种类以乌拉苔草(Carex meyeriana)、唐松草(Thalictrum aquilegifolium L.var. sibiricum Regel)、线叶蒿(Artenisia subulata Nakai)、问荆(Equisetum arvense L.)、珠芽蓼(Polygonum viviparum L.)、蕨麻(Potentilla anserina L.)、散穗早熟禾(Poa subfastigiata Trin.)、青藏苔草(Carex moorcroftii)、棘豆(Oxy tropis sp.)等。
1.2 样地设置2013年5月, 通过对尕海生态环境和植被特征实地调查及相关资料论证分析, 选择刘惠斌等[7]确定的退化泥炭沼泽调查区贡巴点和退化沼泽化草甸调查区尕海湖周边, 采用空间序列代替时间序列生态学方法, 参照《天然草地退化分级指标》的国家标准以及相关湿地分级指标研究[8-9], 在研究区选择地势相对平缓坡向一致的地段, 进行定点样地样方调查。主要依据植被组成、植被总覆盖度、地表裸露面积和地下水位等特征, 将泥炭沼泽划分为未退化(PⅠ)、退化(PⅡ)类型(按照调查区泥炭沼泽退化梯度实际布设为未退化和退化两种类型), 沼泽化草甸划分为未退化(SⅠ)、轻度退化(SⅡ)、中度退化(SⅢ)及重度退化(SⅣ)4种退化类型, 每个退化阶段设立10m×10m定位研究样地(重复3次), 样地基本情况见表 1。泥炭沼泽植物地上、地下生物量于2013—2014年连续2a每年5—9月进行采样测定, 沼泽化草甸于2013—2015年连续3a每年5—9月进行采样测定。上述实验均在冬季牧场(6—9月初未放牧, 其他时段放牧)进行, 尕海湿地多年平均降水量和平均温度如图 1所示。
| 湿地类型 Wetland type |
退化阶段 Degradation stage |
海拔 Altitude /m |
植被盖度 Vegetation coverage/% |
平均均高 Mean height/cm |
基本情况 Basic conditions |
| 泥炭沼泽 Peatlands |
未退化(PⅠ) | 3612 | >90 | 9.42 | 沼泽植物为优势种, 几乎无裸班, 常年积水, 地下水位15—20 cm |
| 退化(PⅡ) | 3590 | 40—70 | 8.66 | 沼泽植物为主要伴生种, 地表裸露面积10%—20%, 无积水, 地下水位30—50 cm | |
| 沼泽化草甸 Swamp meadows |
未退化(SⅠ) | 3477 | >90 | 16.31 | 湿生植物为优势种, 枯落物和根系较多, 几乎无裸班, 有较浅的季节性积水, 地下水位20—40 cm |
| 轻度退化(SⅡ) | 3478 | 75—90 | 13.33 | 湿生植物为主要伴生种, 地表裸露面积5%—10%, 无积水, 地下水位40—70 cm | |
| 中度退化(SⅢ) | 3484 | 30—70 | 7.61 | 湿生植物为一般伴生种或偶见种, 且出现一些毒草, 地表裸露面积10%—30%, 无积水, 地下水位70 cm以下 | |
| 重度退化(SⅣ) | 3486 | 植被很稀疏, 仅有零星的旱生植物出现, 地表裸露面积90%以上 | |||

|
| 图 1 尕海湿地多年月平均温度和降水分布 Fig. 1 Mean monthly precipitation and temperature of Gahai wetland, Gansu Province |
试验期间每年5—9月的中旬左右在定位研究样地内随机选取5个50 cm×50 cm的小样方, 用剪刀将样方内的植物齐地面剪下, 装袋带回实验室, 并置于烘箱内, 在80℃温度下烘至恒重, 计算每个样方的地上生物量。
地下生物量在地上生物量调查的样方内进行。随机选取上述3个样方, 采用挖土块法取样, 土块面积大小为20 cm×20 cm, 高度为10 cm, 按照10 cm为1层, 分层进行采集, 采集深度为30 cm(研究区主要以草地为主, 而草地根系主要分布在30 cm, 且随湿地退化程度增加, 30 cm层植物根系量减小幅度最为显著)。野外采集的地下植物根系土样带回室内, 用细筛(孔径1 mm, 泰勒标准16目)筛去土, 再用细纱布包好根系, 用清水洗净, 并拣出石块和其他杂物, 在80℃恒温箱烘干至恒重。以上生物量最后都换算成单位面积地上干物质量(g/m2)。在沼泽化草甸重度退化阶段, 植被非常稀疏, 实验中未对其进行生物量调查。
1.4 地上生物量累积量和增长率计算地上生物量累积量用5—8月份测定地上干物质量差值的累积量估算, 即:
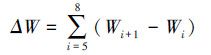
|
生物量的增长率采用生长分析方法中的绝对增长速率(AGR)和相对增长速率(RGR)用于分析和解释单位时间内生物量的净积累值。计算公式如下[10]:
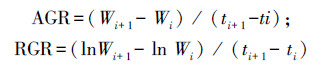
|
式中, Wi、Wi+1分别表示Wi、Wi+1月份的地上生物量。
1.5 数据处理与分析本文采用Excel 2003制图, 用SPSS 17.0软件进行数据统计分析, 采用单因素方差分析和LSD多重比较法进行差异显著性检验, 文中泥炭沼泽生物量为2013、2014年两年每月生物量均值, 沼泽化草甸为2013、2014、2015年3年每月生物量均值, 地下生物量垂直分布是2015年5—9月测定的均值。
2 结果与分析 2.1 植物地上生物量 2.1.1 地上生物量及其季节动态随着甘南尕海高寒湿地退化, 植物群落发生了显著变化, 植物盖度和地上生物量明显下降, 其变化规律如图 2所示。泥炭沼泽未退化阶段年均植被地上生物量(334.19 g/m2)>退化阶段(290.72 g/m2), 沼泽化草甸未退化(378.40 g/m2)>轻度退化(308.07 g/m2)>中度退化(261.21 g/m2)。各湿地随退化演替, 最大累积量降低幅度存在一定差异。其中, 泥炭沼泽生长季(5—8月)最大累积量从未退化505.56 g/m2降低到退化的453.08 g/m2, 降低了10.38%;而沼泽化草甸5—8月累积量从未退化641.47 g/m2分别降低到轻度、中度退化的471.85, 398.38 g/m2, 分别降低了26.47%和37.92%。说明湿地退化显著降低了高寒湿地植被地上生产力。

|
| 图 2 不同退化阶段地上生物量变化及其季节动态 Fig. 2 Seasonal dynamics and Changes of above ground biomass in different degeneration types |
尕海泥炭沼泽和沼泽化草甸不同退化阶段植被地上生物量的季节变化也异常明显。2类湿地各退化阶段地上生物量季节变化趋势相同, 均表现为单峰型生长曲线。各退化阶段均在5月中下旬开始返青, 而后随气温及水分条件的改善生物量逐渐增加, 峰值出现在8月份。到9月份, 随生长季结束, 尕海湿地气温、地温显著降低(图 2), 植物光合能力逐渐减弱, 植物生长期逐渐结束, 营养物质开始溶失并向地下根系转移, 且转移能力日益增强, 致使地上生物量开始减少, 到10月中下旬左右, 植物完全停止生长, 地上部分完全枯死, 呈立枯状或枯落归还地下, 说明2类湿地各退化阶段植物生长节律与该区降雨和温度密切相关。
2.1.2 地上生物量增长速率季节与年际变化为分析尕海湿地各退化阶段在不同年份植被地上生物量累计过程, 计算了绝对增长速率(AGR)和相对增长速率(RGR)。泥炭沼泽不同退化阶段植被地上生物量AGR在2013, 2014年季节动态相同, 均在生物量达到高峰期前是正增长, 而后呈负增长变化(表 2)。但两年最快生长期出现时间并不一致, 其中, 2013年未退化和退化的绝对增长速率最大值出现在7—8月, 峰值分别为30.14, 22.17 g m-2 d-1;2014年最大值出现在5—6月, 而7—8月仍属于较快增长期, 仅比前一时间段降低了0.60 g m-2 d-1和0.11 g m-2 d-1。沼泽化草甸各退化阶段绝对增长速率最大值2013, 2014年相同, 均出现在7—8月, 且2013年明显高于2014年。在2013年, 未退化、轻度退化和中度退化峰值为54.27, 43.38, 31.57 g m-2 d-1, 但在2014年, 仅为3.26, 3.42, 1.23 g m-2 d-1, 2013年显著高于2014年。在2015年, 沼泽化草甸未退化、轻度退化和中度退化阶段峰值均出现在6—7月, 值分别为9.78, 9.45, 9.52 g m-2 d-1;同时, 在2013和2014年的5—6月期间, 沼泽化草甸各退化阶段上生物量AGR均为负值, 这是由于沼泽化草甸样地位于尕海湖边, 在5—6月湖边经常会受到放牧干扰, 样地可能受到放牧干扰引起。总体上, 从8月下旬开始, 随着气温下降和植物衰退、枯黄及营养物质转移, 引起各湿地退化阶段植被地上生物量绝对增长速率逐渐降低, AGR出现负值。由此可见, 尕海泥炭沼泽各湿地退化阶段AGR为缓慢增长—快速积累—快速折损减少等3个不同阶段, 尕海沼泽化草甸为缓慢减少—缓慢增长—快速积累—快速折损减少等4个不同阶段, 这与地上生物量的动态规律基本一致。
| 年份 Year |
退化阶段 Degradation stage |
5—6月Month | 6—7月Month | 7—8月Month | 8—9月Month | |||||||
| AGR/ (g m-2 d-1) |
RGR | AGR/ (g m-2 d-1) |
RGR | AGR/ (g m-2 d-1) |
RGR | AGR/ (g m-2 d-1) |
RGR | |||||
| 2013 | PⅠ | 1.17 | 0.019 | 4.29 | 0.030 | 30.34 | 0.054 | -27.56 | -0.043 | |||
| PⅡ | 0.96 | 0.019 | 5.62 | 0.040 | 22.17 | 0.043 | -23.17 | -0.048 | ||||
| SⅠ | -2.57 | -0.010 | 1.27 | 0.006 | 54.27 | 0.068 | -49.18 | -0.051 | ||||
| SⅡ | -4.41 | -0.025 | 3.97 | 0.026 | 43.80 | 0.063 | -32.94 | -0.034 | ||||
| SⅢ | -1.62 | -0.011 | 0.94 | 0.007 | 31.57 | 0.066 | -17.28 | -0.021 | ||||
| 2014 | PⅠ | 1.71 | 0.032 | 1.10 | 0.011 | 0.22 | 0.002 | -3.15 | -0.040 | |||
| PⅡ | 1.06 | 0.020 | 0.95 | 0.013 | 0.30 | 0.003 | -3.08 | -0.055 | ||||
| SⅠ | -0.21 | -0.002 | 2.71 | 0.014 | 3.26 | 0.020 | -1.52 | -0.008 | ||||
| SⅡ | -0.44 | -0.006 | 1.30 | 0.014 | 3.42 | 0.024 | -0.39 | -0.004 | ||||
| SⅢ | -0.33 | -0.007 | 1.08 | 0.019 | 1.23 | 0.013 | -1.00 | -0.010 | ||||
| 2015 | SⅠ | - | - | 9.78 | 0.036 | 0.93 | 0.002 | -1.21 | -0.003 | |||
| SⅡ | - | - | 9.45 | 0.035 | -7.10 | -0.021 | 2.87 | 0.010 | ||||
| SⅢ | - | - | 9.52 | 0.028 | -7.35 | -0.018 | -2.93 | -0.011 | ||||
| AGR:绝对增长速率Absolute growth rate;RGR相对增长速率Relative growth rate | ||||||||||||
RGR能够反映植物的生产效率, 这与AGR不同。2类湿地各退化阶段植物群落地上生物量的RGR与AGR的季节动态变化趋势上大致相同, 但年际变化存在一定差异。在2013年, 2类湿地各退化阶RGR最大值出现在7—8月, 在2014年, 最大值却出现在6—7月, 而沼泽化草甸各退化阶段地上生物量RGR最大值2015年出现在6—7月, 出现这种变化的原因是由于不同年份气候条件不同, 特别是降水发生变化引起。但当RGR到最大值后, 均变为负值, 说明在植物生长旺盛后, 植物RGR与湿地植物生长发育节律有关, 受环境因子影响相对较少。
2.2 植物地下生物量 2.2.1 地下生物量及其季节动态随着退化程度加剧, 湿地植被地下生物量呈明显下降趋势, 未退化明显高于退化(图 3)。具体为:在泥炭沼泽中, 未退化、退化地5—9月平均地下生物量分别为23081.46 g/m2和12607.72 g/m2, 退化地下生物量呈极显著减少(P<0.01), 减少了约1倍;但在沼泽化草甸中, 未退化、轻度退化、中度退化湿地植被地下生物量5—9月平均值分别为4583.16, 3008.63 g/m2和1290.73 g/m2。轻度退化、中度退化分别为未退化的65.65%和28.16%, 地下生物量显著减少(P<0.05)。

|
| 图 3 不同退化阶段地下生物量变化及其季节动态 Fig. 3 Seasonal dynamics and Changes of below ground biomass in different degeneration types |
两类典型湿地不同退化阶段植被地下生物量季节变化明显(图 3), 两类湿地不同退化阶段植被地下生物量季节变化存在一定差异, 但均表现为生长季初期, 地下生物量较大, 然后逐渐下降, 到7月达到最低值, 而后逐渐升高, 并于生长季末达到最大值(除泥炭沼泽未退化外)。泥炭沼泽未退化阶段植被地下生物量季节变化趋势基本表现为“N”型变化规律, 且变化幅度较大, 峰值出现在8月份, 为25727.17 g/m2, 最小值在9月份, 为20380.33 g/m2;受湿地退化影响, 退化泥炭沼泽植被地下生物量季节变化趋势呈“V”型变化规律, 这是由于随着植物快速生长, 地上部生长消耗开始高于地下部贮存量, 致使地下生物量减少, 到8月份以后, 随着生长季末期到来, 地上生长逐渐变慢, 多余营养转化到地下, 地下生物量逐渐增大。
沼泽化草甸各退化阶段植被地下生物量与泥炭沼泽明显不同, 这可能是两类湿地植物优势种类及所处环境条件不同所致。总体来看, 沼泽化草甸各退化阶段植被地下生物量季节变化基本表现为“V”型变化趋势, 未退化、轻度退化、中度退化地下生物量分别在生长季初期和末期达到最大值, 但初期明显高于末期, 初期分别为7549.12, 4006.69, 2932.34 g/m2, 而末期为4983.04, 3844.41, 740.77 g/m2, 最低值均出现在7月份, 值分别为1914.94, 1238.38, 287.71 g/m2, 说明在生长初期, 沼泽化草甸受低温影响, 植物地上发育比较缓慢, 主要营养物质主要转移到地下部, 使得地下生物量较高。
2.2.2 地下生物量垂直分布特征甘南尕海两类典型湿地各退化阶段植被地下生物量在不同土层均值如表 3。在泥炭沼泽中, 各退化阶段表层(0—10 cm)植被地下生物量值最大, 占其总地下生物量54%以上, 未退化、退化阶段植被地下生物量分别为8262.67, 4267.28 g/m2, 退化阶段显著降低(P<0.05);到10—20 cm土层, 地下生物量逐渐减小, 值为3825.68, 3119.50 g/m2, 未退化显著减小(P<0.05), 幅度为53.7%, 退化为26.9%;20—30 cm土层地下生物量分别为2970.66, 434.32 g/m2, 退化阶段显著降低(P<0.05), 幅度为86.1%。在沼泽化草甸, 各退化阶段表层(0—10 cm)地下生物量也最大, 占总地下生物量75%以上, 未退化地下生物量显著高于轻度、中度退化(P<0.05), 未退化、轻度退化和中度退化地下生物量分别为2076. 81, 1123.67, 653.57 g/m2;10—20 cm土层植被地下生物量分别为523.09, 208.41, 98.96 g/m2, 生物量迅速减小, 其中未退化减少幅度在70%以上, 其他退化阶段在80%以上;各退化阶段20—30 cm土层地下生物量差异不显著(P>0.05), 分别为157.21, 111.29, 38.81 g/m2, 生物量也相应减少。地下生物量0—30 cm各土层变化趋势与总地下生物量变化趋势基本相同, 均随湿地退化加剧, 地下生物量逐渐降低。在泥炭沼泽, 除10—20 cm外, 其他两层未退化生物量都显著高于退化(P<0.05)。沼泽化草甸各退化阶段0—10 cm地下生物量间均存在显著差异(P<0.05);10—20 cm地下生物量未退化显著高于轻度和中度(P<0.05), 轻度与中度退化地下生物量差异不显著(P>0.05);20—30 cm地下生物量未退化显著高于中度退化, 但与轻度退化差异不显著(P>0.05)。从地下生物量垂直分布看, 随土层深度增加生物量逐渐减小, 总体表现为“T”字形下降的锯齿状分布(图 4)。两类湿地各退化阶段植被地下生物量均在表层0—10 cm最大, 随着土层增加逐渐减小, 泥炭沼泽各退化阶段70%以上的地下生物量主要集中在0—20 cm土层, 而沼泽化草甸70%以上的地下生物量主要集中在0—10 cm土层。
| 湿地类型 Wetland types |
退化类型 Degeneration types |
0—10 cm | 10—20 cm | 20—30 cm | |||||
| 生物量 Biomass/ (g/m2) |
% | 生物量 Biomass/ (g/m2) |
% | 生物量 Biomass/ (g/m2) |
% | ||||
| 泥炭沼泽 Peat | PⅠ | 8262.67Aa | 54.87% | 3825.68Ab | 25.40% | 2970.66Ab | 19.73% | ||
| PⅡ | 4267.28Ba | 54.56% | 3119.50Aa | 39.89% | 434.32Bb | 5.55% | |||
| 沼泽化草甸 Swamp meadow |
SⅠ | 2076. 81Aa | 75.33% | 523.09Ab | 18.97% | 157.21Abc | 5.70% | ||
| SⅡ | 1123.67Ba | 77.85% | 208.41Bb | 14.44% | 111.29ABb | 7.71% | |||
| SⅢ | 653.57Ca | 82.59% | 98.96Bb | 12.50% | 38.81BCb | 4.90% | |||
| 标有不同小写字母(同一阶段)或大写字母(同一土层)表示有显著性差异(P<0.05), 否则无显著差异 | |||||||||

|
| 图 4 不同退化阶段地下生物量垂直分布 Fig. 4 Vertical distribution of the belowground biomass in different degeneration types |
甘南尕海两类典型湿地不同退化演替阶段的地下与地上物量比值(根冠比)差异较大, 且季节变化明显如图 5具体为:在泥炭沼泽中, 未退化、退化地5—9月平均根冠比为85.33和61.05, 统计分析表明差异不显著(P>0.05);在沼泽化草甸中, 未退化、轻度退化、中度退化湿地平均根冠比分别为18.06, 14.21和8.79, 统计分析表明差异不显著(P>0.05)。

|
| 图 5 不同退化阶段根冠比季节动态 Fig. 5 Seasonal dynamics of root-shoot ratio in different degeneration types |
从季节变化来看, 两类湿地各退化阶段变化趋势存在一定差异。在泥炭沼泽中, 未退化样地最大值出现在生长季的第2月, 而退化样地最大值在生长末期, 总体趋势在生长初期较大, 根冠比随着植物生长旺盛时间的到来, 开始减小, 到8月份达到最低值, 然后迅速回升, 方差分析显示, 除8, 9月份两种退化类型根冠比差异不显著外(P>0.05), 其他月份存在显著差异(P<0.05)。在沼泽化草甸中, 除轻度退化在6月份异常外, 其他时间各退化阶段变化趋势基本一致, 均在生长季初期的返青时期最大, 到8月出现最小, 而后又逐渐增加的趋势, 经方差分析, 未退化和中度退化根冠比在7, 8差异不显著(P>0.05), 其他各月均差异显著(P<0.05)。
3 讨论 3.1 尕海湿地退化对植被地上生物量的影响湿地退化已成为我国重大的资源和环境问题之一, 尕海湿地生态系统脆弱、抗干扰能力差, 湿地退化尤为突出, 这与其所在的青藏高原生态系统环境具有一致性。湿地退化会改变湿地植被地上初级生产力, 进而影响本区域湿地碳汇能力, 也进一步加速湿地土壤发生退化[6]。受全球气候变暖、湿地排水修路, 特别近年来过度放牧的影响, 尕海湿地出现一定程度退化[5], 致使湿地生态系统植被地上生产力显著下降。本研究显示, 随着尕海湿地退化程度的加剧, 植被地上生物量显著减少。在草地生态系统研究表明, 放牧一方面通过伤害植物的正常组织、形态, 抑制植物生长, 另一方面通过移去植物顶端组织和衰老组织, 刺激植物生长[11], 而植物受损程度在其阈值范围以内可以通过自身的生理机制进行恢复, 若超过该阈值则会发生退化, 引起生产力下降[12], 这说明尕海湿地生态系统植物受损程度已经超过了其受损程度所在的阈值范围。此外, 尕海湿地退化过程主要是过度利用植被而导致其生产力降低乃至完全丧失过程, 有研究已表明这一过程能使土壤板结、物质循环放慢、地表覆盖物和土壤的枯枝落叶减少[13]。
尕海湿地植被由于受低温的影响, 植物的返青期都比较晚, 一般在5月中旬湿地植被才陆续返青, 但在初期由于受外界放牧和环境因素影响, 生物量并未显著增加, 到6于中旬, 随着牧场转移, 各湿地地上生物量开始积累, 并随着植物生长发育节律、气温回升和降水量增加而逐步增长, 呈单峰型生长曲线, 生物量峰值出现在降雨和温度最佳的8—9月份, 揭示了水分、温度等环境因子对湿地植物生活史的影响。这与祁连山北坡天然草地[14]、青海高寒草甸[15]生长规律一致。生长高峰期过后, 受气温下降, 湿地群落植物光合作用减弱, 加之植物个体逐渐衰老, 枯落量增加, 营养物质不断流失并向地下根系转移, 导致地上生物量日趋减少, 这与三江平原典型湿地生态系统生物量变化规律一致, 但是衰退期晚于三江平原[16], 可能是由于两湿地所处地理位置不同引起。对地上生物量绝对增长速率(AGR)和相对增长速率(RGR)季节动态分析表明尕海湿地植物生物量积累大致可分为3个阶段缓慢增长—快速积累—快速折损减少, 这与生物量动态变化具有一致性。但不同年份AGR和相对RGR存在一定差异, 在2013年和2014年变化基本相同, 而2015年差别较大。在2015年7—8月, AGR和RGR均为负值, 这可能于2015年降水量较少有关。研究发现, AGB与降水量有显著正相关关系, 并且认为除降水外, AGB还与上一年地上生产力变化有关[17];也有研究认为, 地上生产力与年际降水量有密切关系, 地上生产力随着年降水量增加而增大[18], 本研究结果与上述学者研究结果基本一致;但在自然状态下湿地群落AGB受温度的影响显著, 而扰动状态下湿地群落AGB受温度影响规律不明显, 说明湿地退化过程中植被地上部分生长节律发生改变, 致使地上部分与温度之间的关系变得更复杂。
3.2 尕海湿地退化对植被地下生物量的影响地下生物量是湿地植被碳蓄积的重要组成部分, 是联系地上和地下生态系统过程的主要纽带[19], 准确测定地下生物量是确定湿地植被碳源汇功能的基础。受气候变暖、长期超载过牧等人为活动的影响, 青藏高原湿地生物多样性逐渐降低、湿地植被严重退化[5], 湿地生产力急剧下降, 致使其地下生物量的研究存在较大不确定性。同时, 由于目前对地下生物量研究缺乏统一的采样标准和方法[20], 导致对湿地地下生物量生态过程的研究存在一些偏差。在关于地下生物量研究报道中, 主要研究集中在天然未退化生态系统的地下生物量[21-22], 或者对不同草地类型、不同干扰强度地下生物量分布规律进行分析[23-26], 但青藏高原湿地已发生一定程度退化, 以未退化湿地植被地下生物量结果对湿地生态系统物质循环和功能分析, 有时会带来较大偏差, 不能很好地反映真实情况。为此, 本研究针对湿地不同退化程度高寒湿地, 分析了湿地演替各阶段的植被地下生物量季节动态及垂直分布格局, 为准确估算湿地地下生物量提供了有效的数据基础和参考价值。
在尕海湿地, 随着湿地退化程度的加剧, 植被地下生物量呈明显下降趋势, 未退化明显高于退化, 这是由于湿地退化程度加剧, 引起地上植被数量减少, 致使植物光合面积降低, 尤其在植物返青生长期, 其光合产物不能满足其自身生长发育, 需要地下根系提供营养物质, 这不仅显著影响植物根系生长发育, 而且制约地上、地下干物质积累, 久而久之, 有些植物种不能完成其生命全过程, 逐渐衰退和消失, 最终导致地下生物量减少, 这与西藏那曲地区高寒矮嵩草草甸植被退化过程中地下生物量变化一致[24]。
地下生物量也是时间函数, 研究地下生物量在时间序列上变化具有十分重要的理论和现实意义[20, 27]。本研究中尕海两类湿地各退化阶段植被地下生物量季节变化存在差异, 但均表现出愈接近生长季的始末其值愈大(除未退化泥炭沼泽外)。泥炭沼泽各未退化阶段植被地下生物量季节变化趋势基本表现为“N”型变化曲线, 与祁连山山地草甸、山地草甸草原、山地草原和山地荒漠草原地下生物量季节动态颇为相似[14], 而沼泽化草甸各退化阶段植被地下生物量表现为“V”型变化曲线, 曲线最低值出现在7月, 表明同一地区, 不同湿地类型各植被退化阶段地下生物量动态趋势存在差异。与地上生物量相比, 其动态趋势变化较为复杂。
此外, 前人对草原地下生物量研究表明, 草原地下生物量主要集中在土壤表层, 随着深度的增加, 生物量急剧降低, 呈“T”形分布[14, 24, 27-29], 本研究也表明, 不同退化程度湿地植被地下生物量由表层向下逐渐降低, 垂直分布也呈“T”形分布。各退化阶段植物根系主要集中在0—10 cm土壤表层, 泥炭沼泽占总地下生物量50%以上, 而沼泽化草甸占70%以上。随湿地退化程度加剧, 引起土壤质地粗化, 土壤肥力降低, 进而影响植被生长发育, 导致植被地下生物量大幅下降。未退化湿地植被生长有充足地下土壤养分补给, 而退化湿地植被生存的土壤环境已发生一定程度恶化, 土壤养分匮乏[17], 植物根系获取养分越来越少。由于湿地类型不同, 致使2类湿地各退化阶段植被地下生物量垂直分布变化幅度存在一定差异。特别指出的是, 高寒湿地地下生物量往往是由土壤质地、温度、水分和养分等多因子相互关联综合作用, 随着湿地退化程度加剧, 环境因素影响植被地下生物量的大小及其生态系统功能将明显增强。
3.3 湿地退化对植被地下、地上生物量分配的影响植物体的各部分是一个统一的整体, 地上部分对地下部分的生长发育有重要影响, 地上和地下生物量分配关系支持等速生长模式[30]。地下生物量与地上生物量的比值(根冠比)反映了光合作用物质在植物体内的分布[31]。尕海湿地未退化阶段根冠比显著高于退化阶段, 并随退化程度加剧根冠比降低幅度逐渐增大, 这是由于植物体在受到生长限制因素限制时, 生长策略往往是将受限制的资源优先分配给受限制因素影响最大的器官或组织。随尕海湿地退化程度加剧, 湿地水分含量显著降低, 植物会将更多的水分和生物量分配给地下部分[32]。2类湿地各退化阶段根冠比季节动态均为越接近生长季始末值越大, 这与柴曦等人研究结果一致[33]。植物在生长季末, 地下生物量要远大于地上生物量, 根系中积累的营养物质用于维持植物冬天呼吸消耗, 进入返青期(5—6月份)后, 随着温度和降雨升高, 植物地上冠层逐渐恢复生长, 光合产物积累越来越多地上部生长速率超过了地下部生长速率, 根冠比开始减小, 但是2类湿地根冠比峰值出现时间不同, 泥炭沼泽出现在生长季末9月, 沼泽化草甸出现在生长季开始5月, 由于泥炭沼泽样地属于公共放牧地, 牛羊在生长季末期、早期的大量啃食致使地上现存生物量较低, 引起根冠比较大。
4 结论(1) 随着甘南尕海高寒湿地退化程度的加剧, 植物盖度和地上生物量明显下降, 最大累积量也逐渐降低, 但降低幅度趋于减小。2类湿地各退化阶段植被地上生物量的季节动态均为单峰型曲线, 峰值出现在8月份, 地上生物量生长节律与该区降雨和温度密切相关。2类湿地各退化阶段地上生物量绝对增长速率和相对增长速率季节动态相同, 均在达到高峰期前是正增长, 而后呈负增长变化, 但不同年份最快生长期出现时间不同, 泥炭沼泽2013年在7—8月, 2014年在5—6月, 而沼泽化草甸2013, 2014年均在7—8月, 2015年在5—6月。
(2) 随着尕海2类湿地退化程度加剧, 植被地下生物量呈明显下降趋势, 表现为未退化显著高于其他退化阶段(P<0.05)。但2类湿地不同退化阶段植被地下生物量季节变化存在差异, 泥炭沼泽未退化阶段植被地下生物量季节变化趋势表现为“N”型变化规律, 且变化幅度较大, 地下生物量峰值出现在8月份, 为25727.17 g/m2, 最小值在9月份, 为20380.33 g/m2;沼泽化草甸各退化阶段植被地下生物量季节变化趋势基本表现为“V”型变化规律, 未退化、轻度退化、中度退化地下生物量分别在生长季初、末期达到最大值, 最低值均出现在7月份, 分别为1914.94, 1238.38, 287.71 g/m2。2类湿地地下生物量随土层深度的增加逐渐减小, 总体表现为“T”字形下降趋势的锯齿状分布, 且地下生物量在0—10 cm层最大。
(3) 尕海2类高寒湿地5—9月平均根冠比均表现未退化高于退化, 2类湿地根冠比的季节变化存在差异, 泥炭沼泽值, 未退化样地最大值出现在生长季的第2月, 而退化样地最大值生长末期9月;沼泽化草甸为生长季初期的返青时期最大, 到8月出现最小, 而后又逐渐增加。
| [1] | Lund M, Lindroth A, Christensen T R, Ström L. Annual CO2 balance of a temperate bog. Tellus B, 2007, 59(5): 804–811. DOI:10.1111/j.1600-0889.2007.00303.x |
| [2] | Sutton-Grier A E, Megonigal J P. Plant species traits regulate methane production in freshwater wetland soils. Soil Biology and Biochemistry, 2011, 43(2): 413–420. DOI:10.1016/j.soilbio.2010.11.009 |
| [3] | 谢长永, 陈波, 徐同凯. 杭州西溪湿地植物生物量和有机碳季节变化及碳的积累. 杭州师范大学学报:自然科学版, 2015, 14(3): 285–291. |
| [4] | 马维伟, 王辉, 李广, 赵锦梅, 王跃思. 尕海湿地CH4、CO2和N2O通量特征初步研究. 草业学报, 2015, 24(8): 1–10. |
| [5] | 刘帆, 刘晖, 徐凌翔, 程玉, 李伟. 青藏高原东部尕海湿地种子库沿水位梯度的分布格局. 科学通报, 2013, 58(14): 1332–1339. |
| [6] | 马维伟, 王辉, 李广, 石万里. 甘南尕海湿地不同植被退化阶段土壤有机碳含量及动态. 水土保持学报, 2015, 29(5): 254–259. |
| [7] | 刘惠斌, 李俊臻. 甘肃尕海湿地退化泥炭地恢复技术评价. 湿地科学与管理, 2010, 6(2): 26–29. |
| [8] | 张晓龙, 李培英. 湿地退化标准的探讨. 湿地科学, 2004, 2(1): 36–41. |
| [9] | 刘育红, 李希来, 李长慧, 孙海群, 芦光新, 潘根兴. 三江源区高寒草甸湿地植被退化与土壤有机碳损失. 农业环境科学学报, 2009, 28(12): 2559–2567. DOI:10.3321/j.issn:1672-2043.2009.12.019 |
| [10] | 郭雪莲, 吕宪国, 戴国华. 三江平原不同水位梯度湿地地上生物量动态特征. 生态环境, 2008, 17(5): 1739–1742. |
| [11] | Huhta A P, Hellström K, Rautio P, Tuomi J. Grazing tolerance of Gentianella amarella and other monocarpic herbs:why is tolerance highest at low damage levels?. Plant Ecology, 2003, 166(1): 49–61. DOI:10.1023/A:1023278502972 |
| [12] | 方楷, 宋乃平, 魏乐, 安慧. 不同放牧制度对荒漠草原地上生物量及种间关系的影响. 草业学报, 2012, 21(5): 12–22. DOI:10.11686/cyxb20120502 |
| [13] | 高雪峰, 韩国栋, 张功, 赵萌莉, 卢萍. 放牧对荒漠草原土壤微生物的影响及其季节动态研究. 土壤通报, 2007, 38(1): 145–148. |
| [14] | 黄德青, 于兰, 张耀生, 赵新全. 祁连山北坡天然草地地下生物量及其与环境因子的关系. 草业学报, 2011, 20(5): 1–10. DOI:10.11686/cyxb20110501 |
| [15] | 刘文辉, 周青平, 颜红波, 梁国玲. 青海扁茎早熟禾种群地上生物量积累动态. 草业学报, 2009, 18(2): 18–24. DOI:10.11686/cyxb20090204 |
| [16] | 杨永兴, 王世岩, 何太蓉, 田昆, 杨波. 三江平原典型湿地生态系统生物量及其季节动态研究. 中国草地, 2002, 24(1): 1–7. |
| [17] | 金云翔, 徐斌, 杨秀春, 李金亚, 马海龙, 高添, 于海达. 不同沙化程度草原地下生物量及其环境因素特征. 草业学报, 2013, 22(5): 44–51. DOI:10.11686/cyxb20130506 |
| [18] | Fang J Y, Piao S L, Tang Z Y, Peng C H, Ji W, Knapp A K, Smith M D. Interannual variability in net primary production and precipitation. Science, 2001, 293(5536): 1723–1723. DOI:10.1126/science.293.5536.1723a |
| [19] | 吴琴, 尧波, 朱丽丽, 幸瑞新, 胡启武. 鄱阳湖典型苔草湿地生物量季节变化及固碳功能评价. 长江流域资源与环境, 2012, 21(2): 215–219. |
| [20] | 宇万太, 于永强. 植物地下生物量研究进展. 应用生态学报, 2001, 12(6): 927–932. |
| [21] | 周华坤, 周立, 赵新全, 沈振西, 李英年, 周兴民, 严作良, 刘伟. 金露梅灌丛地下生物量形成规律的研究. 草业学报, 2002, 11(2): 59–65. |
| [22] | 何池全. 毛果苔草湿地枯落物及其地下生物量动态. 应用生态学报, 2003, 14(3): 363–366. |
| [23] | 余欣超, 姚步青, 周华坤, 金艳霞, 杨月娟, 王文颖, 董世魁, 赵新全. 青藏高原两种高寒草甸地下生物量及其碳分配对长期增温的响应差异. 科学通报, 2015, 60(4): 379–388. |
| [24] | 鄢燕, 张建国, 张锦华, 范建容, 李辉霞. 西藏那曲地区高寒草地地下生物量. 生态学报, 2005, 25(11): 2818–2823. DOI:10.3321/j.issn:1000-0933.2005.11.004 |
| [25] | Molyneux D E, Davies W J. Rooting pattern and water relations of three pasture grasses growing in drying soil. Oecologia, 1983, 58(2): 220–224. DOI:10.1007/BF00399220 |
| [26] | Scurlock J M O, Johnson K, Olson R J. Estimating net primary productivity from grassland biomass dynamics measurements. Global Change Biology, 2002, 8(8): 736–753. DOI:10.1046/j.1365-2486.2002.00512.x |
| [27] | 胡中民, 樊江文, 钟华平, 韩彬. 中国草地地下生物量研究进展. 生态学杂志, 2005, 24(9): 1095–1101. |
| [28] | 李凯辉, 王万林, 胡玉昆, 高国刚, 公延明, 尹伟. 不同海拔梯度高寒草地地下生物量与环境因子的关系. 应用生态学报, 2008, 19(11): 2364–2368. |
| [29] | 王艳芬, 汪诗平. 不同放牧率对内蒙古典型草原地下生物量的影响. 草地学报, 1999, 7(3): 198–203. DOI:10.11733/j.issn.1007-0435.1999.03.004 |
| [30] | Cheng D L, Niklas K J. Above-and below-ground biomass relationships across 1534 forested communities. Annals of Botany, 2007, 99(1): 95–102. DOI:10.1093/aob/mcl206 |
| [31] | 沈禹颖, 阎顺国, 朱兴运, 赵银. 河西走廊几种盐化草地第一性生产力的研究Ⅰ:地上生物量与地下生物量季节动态及分配. 草业学报, 1995, 4(2): 44–50. |
| [32] | 王敏, 苏永中, 杨荣, 杨晓. 黑河中游荒漠草地地上和地下生物量的分配格局. 植物生态学报, 2013, 37(3): 209–219. |
| [33] | 柴曦, 梁存柱, 梁茂伟, 韩伟华, 李智勇, 苗百岭, 王炜, 王立新. 内蒙古草甸草原与典型草原地下生物量与生产力季节动态及其碳库潜力. 生态学报, 2014, 34(19): 5530–5540. |