 2017, Vol. 37
2017, Vol. 37文章信息
- 陈桂莲, 谭凌照, 范春雨, 张新娜, 张春雨, 赵秀海
- CHEN Guilian, TAN Lingzhao, FAN Chunyu, ZHANG Xinna, ZHANG Chunyu, ZHAO Xiuhai.
- 吉林蛟河阔叶红松林样地种-面积关系
- Species-area relationships within sample plot in a broad-leaved Korean pine forest at Jiaohe, Jilin Province
- 生态学报. 2017, 37(14): 4770-4777
- Acta Ecologica Sinica. 2017, 37(14): 4770-4777
- http://dx.doi.org/10.5846/stxb201604220762
-
文章历史
- 收稿日期: 2016-04-22
- 网络出版日期: 2017-03-02





2. 森林资源生态系统过程北京市重点实验室, 北京 100083
2. Key Laboratory for Forest Resources & Ecosystem Processes of Beijing, Beijing Forestry University, Beijing 100083, China
种-面积关系描述了物种随取样面积的增加而变化的规律, 是群落生态学研究的核心问题之一[1]。种-面积关系可以解决许多生态问题, 诸如确定群落的最小面积, 评估区域物种多样性, 确定自然保护区范围, 预测生境破坏对物种丰富度影响等[2-6]。种-面积关系相较于用单一的物种数目, 可以更好地描述植物群落的物种多样性[7], 对区域生物保护以及物种多样性维持具有重要意义[8]。
目前为止, 生态学家们提出了上百种种-面积关系的拟合方程, 其中应用最广泛的分别是幂函数模型、对数模型和逻辑斯蒂模型, 这3种模型因其参数具有明确的生态学含义并且简单实用, 一直以来被科学家们广泛接受[9-10]。但以上3种模型对种-面积关系拟合的准确性受取样尺度和取样方法的影响较大。种-面积关系应用哪个模型拟合最优, 目前还饱受争议[10]。
大量研究结果表明, 种-面积关系会随着取样尺度不同而发生变化[11], 主要原因在于生境过滤效应以及扩散限制等生态过程影响不同尺度上的物种空间分布格局[12-14]。一些研究表明, 随着尺度变化, 在小尺度和中尺度上适合采用幂函数模型, 而在大尺度上适合采用逻辑斯蒂模型[15-16]。另一些研究, 如:Fisher则认为在小尺度上对数模型能够更好地拟合种-面积曲线[17]。进行多尺度上的种-面积关系研究对于准确反映物种丰富度和空间分布格局背后的生态学机制具有重要意义。
目前的研究着重从生态学理论角度讨论种-面积关系的尺度效应而忽略了生态学方法对种-面积关系的影响, 有研究证明, 取样方法是影响种-面积关系的重要因素[18]。目前最常用的取样方式包括巢式取样和随机取样[19-20]。巢式取样按一定规则不断扩大取样面积, 其中大样方包括小样方, 其研究尺度可从群落扩展至区域乃至大洲[21], 成为构建种-面积关系的主要方式。但Gleason等认为, 巢式取样在构建种-面积关系上易受稀有种、物种聚集以及空间异质性等因素的影响[22], 会降低预测物种数的准确性。随机取样即在研究区域随机选取规定面积的样方来构建种-面积关系曲线, 可用来估计区域种库的大小[23], 在当前的研究中应用越来越广泛。
阔叶红松林是我国温带森林典型的群落类型, 与其他同纬度的森林群落相比, 物种丰富度高, 建群种比较独特, 建立阔叶红松林的种-面积关系对反映东北主要植被类型多样性格局和多样性保护具有重要意义[24], 是当前种-面积关系研究不足之处的补充案例。本文基于吉林蛟河30 hm2阔叶红松林固定监测样地, 利用不同尺度下巢式取样和随机取样数据, 建立了基于幂函数模型、对数模型和逻辑斯蒂模型的区域种-面积关系曲线, 目的在于探讨样地内取样方法和取样尺度对种-面积关系的影响。
1 研究区概况研究区域位于吉林省蛟河林业实验区管理局, 处于吉林省蛟河市前进乡境内, 属于长白山系张广才岭山脉(地理位置:43°57.928′—43°58.214′N, 127°45.287′—127°45.790′E), 海拔为576.03—748.18 m, 最高点与最低点高差达172.15 m(图 1)。研究区域为季风性温带大陆性气候, 年均气温为3.8℃。每年7月为最热月, 平均气温21.7℃, 1月为最冷月, 平均气温-15.4℃。年均降雨量695.9 mm。样地类型为阔叶红松林, 属于长白山植被区系, 该样地的土壤类型为暗棕色森林土, 厚度为20—100 cm。林分的平均胸径为10 cm, 最大胸径达114.5 cm, DBH ≤5 cm的个体占全部个体的51.57%。乔木树种主要包括红松(Pinus koraiensis)、裂叶榆(Ulmus laciniata)、紫椴(Tilia amurensis)、色木槭(Acer mono)、白牛槭(Acer mandshuricum)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、白桦(Betula platyphylla)等, 灌木主要包括暴马丁香(Syringa reticulata)、毛榛(Corylus mandshurica)、瘤枝卫矛(Euonymus pauciflorus)和东北鼠李(Rhamnus davurica)等, 草本主要包括白花碎米荠(Cardamine leucantha)、北重楼(Paris verticillata)、荷青花(Hylomecon Japonicum)、东北延胡索(Corydalis ambigua)、多被银莲花(Anemone raddeana)等。

|
| 图 1 蛟河30 hm2阔叶红松林样地地形模拟图 Fig. 1 Terrain simulation diagram of the 30 hm2 broad-leaved korean pine forest sample area in Jiaohe |
2010年建立30 hm2(500 m×600 m)阔叶红松林固定监测样地。将样地划分为750个20 m×20 m的连续样方。在每个样方中调查样地内所有DBH ≥1 cm的木本植物, 记录其种名、胸径、树高、冠幅(东西冠幅及南北冠幅)和其在整个样地内相对应的位置坐标, 并进行挂牌标记。
2.2 取样尺度基于研究样地面积, 在30 hm2范围内随机选取10、20 hm2的研究区域, 以10、20、30 hm2分别作为小尺度、中尺度、大尺度上构建种-面积关系的上限。
2.3 取样方法以30 hm2取样尺度为例:
巢式取样:在研究区域(500 m×600 m)内随机选取一点, 按照5 m×5 m、10 m×10 m、15 m×15 m、…、495 m ×495 m、500 m×500 m、500 m×540 m、500 m×580 m、500 m×600 m的嵌套方法进行取样, 在抽取的样方中随机选取30个, 分别统计各样方中的物种数, 构建巢式取样的种-面积关系。在取样时若取样单元超出研究区域, 则对无效取样进行剔除, 重新取样。
随机取样:在研究区域(500 m×600 m)内以5 m×5 m、10 m×10 m、15 m×15 m…495 m×495 m、500 m×500 m、500 m×540 m、500 m×580 m、500 m×600 m为规格进行随机取样, 在抽取的样方中随机选取30个, 记录每个样方内的物种数, 构建随机取样的种-面积关系。
由于取样面积变化梯度小, 样方较多而树种变化小, 为降低因随机取样带来的误差, 每次取样重复1000次, 其结果平均值为该面积下的物种数。
2.4 拟合模型对数模型
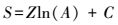
|
幂函数模型

|
逻辑斯蒂模型

|
式中, S为落在样方内的物种数, A为取样面积, C、B和Z均为模型参数。
2.5 种-面积关系拟合优度评价采用最小二乘法对对数模型进行检验, 用高斯牛顿算法(Gauss-Newton)对逻辑斯蒂模型以及幂函数模型进行检验。采用赤池信息准则(AIC)对3种模型的拟合优度进行评价, AIC数值越小, 模型拟合越好, 预测值越准确。
本研究中所有数据分析、作图采用R-3.2.2、Excel 2010完成。
3 结果分析 3.1 不同尺度植物群落组成在10、20、30 hm2 3个研究尺度下, 监测到DBH ≥1的木本植物15610株、32342株、49678株, 分别隶属于19科, 29属, 45种;19科, 31属, 47种;19科, 31属, 50种。蔷薇科(Rosaceae)在3个尺度下物种数最多, 其次是槭树科(Aceraceae)和桦木科(Betulaceae)。而在不同尺度下物种最多的属为槭属(Acer), 其次为桦木属(Betula)、忍冬属(Lonicera)和杨属(Populus)。单种属在3个尺度下分别为17属、19属和20属, 占总属数的58.62%、55.93%、64.5%。
10、20、30 hm2研究尺度下的胸高断面积分别为9.97(m2/hm2)、20.45(m2/hm2)、30.99(m2/hm2), 10 hm2尺度下胸高断面积最大的5个树种依次为:色木槭(Acer mono)、裂叶榆(Ulmus laciniata), 紫椴(Tilia amurensis)、胡桃楸(Juglans mandshurica)、红松(Pinus koraiensis), 占总胸高断面积的57.76%。20 hm2尺度下胸高断面积最大的5个树种依次为:色木槭、裂叶榆、紫椴、红松、胡桃楸, 占总胸高断面积的55.93%。30 hm2尺度下胸高断面积最大的5个树种依次为裂叶榆、色木槭、紫椴、红松、枫桦(Betula costata), 占总胸高断面积的58.08%, 是样地内的先锋树种。
3.2 不同取样方式下的种-面积关系(以30 hm2为例) 3.2.1 巢式取样的种-面积关系在巢式取样下, 根据模型的AIC值, 逻辑斯蒂模型拟合最优, 其次是对数模型。幂函数模型的预测值与观察值之间有一定的差异, 在0—10 hm2之间, 预测物种数低于观察值;在15—30 hm2, 预测值高于观察值, 估计的预测物种数较高, 其拟合效果在3个模型中最差(AIC分别为85.58, 115.20, 167.74)(图 2)。依据种-面积曲线来看, 当取样面积在0—10 hm2之间, 随着取样面积的逐渐增大, 物种数迅速增加, 当取样面积大于10 hm2后, 曲线斜率趋于平缓, 物种数随着取样面积的增加而缓慢增加。当取样面积为8 hm2时, 涵盖了42个种, 占总物种数的84%, 确定为该群落的最小面积。

|
| 图 2 30 hm2样地巢式取样法获得的3种模型的种-面积曲线 Fig. 2 Three species-area curves for nest sample design in a 30-hm2 plot |
随机取样下的3个模型相比较, 逻辑斯蒂模型的拟合效果最好, 其次是对数模型, 幂函数模型的拟合效果最差(AIC分别为76.91、87.40、134.35)。种-面积关系曲线变化规律与巢式取样结果一致(图 3)。

|
| 图 3 30 hm2样地随机取样法获得的3种模型的种-面积曲线 Fig. 3 Three species-area curves for random sample design in a 30-hm2 plot |
不同取样方式下的拟合结果表明, 巢式取样和随机取样下逻辑斯蒂模型的拟合效果都是最优的, 而幂函数模型的拟合效果始终相对较差(表 1), 可能与幂函数增长的趋势易高估预测物种数有关[25]。两种取样方法相比, 随机取样下的逻辑斯蒂模型可以很好地描述该阔叶红松林内的种-面积关系。
| 取样方法 Sampling design | 种-面积模型 Species-area model | 模型参数Model parameters | |||
| C | Z | B | AIC | ||
| 随机取样 | 对数模型 | -18.37*** | 5.27*** | — | 87.40 |
| Random sample design | 幂函数模型 | 4.82*** | 0.19*** | — | 134.35 |
| 逻辑斯蒂模型 | 0.02*** | 0.40*** | 1.44*** | 76.91 | |
| 巢式取样 | 对数模型 | -15.52*** | 5.22*** | — | 115.20 |
| Nest sample design | 幂函数模型 | 7.18*** | 0.16*** | — | 167.74 |
| 逻辑斯蒂模型 | 0.01*** | 0.51*** | 0.75*** | 85.58 | |
| C、Z、B是常数项;***P<0. 001;AIC为赤池信息准则, 值越小代表拟合效果越好 | |||||
基于样地范围划分大、中、小3个尺度范围, 取样上限分别为30、20、10 hm2。不同尺度下种-面积关系的结果表明(表 2), 在小尺度上, 随机取样与巢式取样下的对数模型和逻辑斯蒂模型拟合效果相差不大, 均优于两种取样方法下的幂函数模型。在中尺度上, 以逻辑斯蒂模型最优, AIC值分别为74.59和79.90。大尺度上, 两种取样方法下的逻辑斯蒂模型拟合效果优于对数模型和幂函数模型, 其中随机取样优于巢式取样。
| 取样方法 Sampling design | 种-面积模型 Sampling design | AIC | ||
| 10 hm2 | 20 hm2 | 30 hm2 | ||
| 随机取样 | 对数模型 | 73.91 | 78.11 | 87.40 |
| Random sample design | 幂函数模型 | 106.22 | 124.36 | 134.35 |
| 逻辑斯蒂模型 | 80.51 | 74.59 | 76.91 | |
| 巢式取样 | 对数模型 | 85.45 | 104.61 | 115.20 |
| Nest sample design | 幂函数模型 | 139.11 | 155.98 | 167.74 |
| 逻辑斯蒂模型 | 73.30 | 79.90 | 85.58 | |
近年来, 国内外的学者对种-面积关系进行了多方面的研究, 例如:相对物种丰富度(RSA)、β多样性与种-面积关系的联系[26]、种-面积和种-个体间的关系[27]等, 结果表明种-面积关系对反映地区生物多样性规律有基础性的意义。该研究基于吉林蛟河30 hm2阔叶红松林大样地, 分析了作为东北地区典型植被的种-面积关系变化规律, 讨论了取样方式和取样尺度对蛟河阔叶红松林种-面积关系变化的影响。
种-面积曲线(图 2, 图 3)确定了吉林蛟河阔叶红松林的最小面积为8 hm2。物种增加速率随取样面积的增加而降低, 当取样面积为8 hm2时, 出现42个物种, 占总物种数的84%。而长白山次生杨桦林的研究表明, 在取样面积为5 hm2时, 曲线趋于稳定, 出现近35个物种, 占总物种数的80%[28]。比较蛟河样地和长白山样地发现, 蛟河森林群落的最小取样面积大于长白山森林群落的最小取样面积, 可能是因为蛟河森林群落物种分布相对较为分散。
种-面积模型具有尺度效应, 不同尺度下模型的拟合优度不同。研究表明, 研究区域的种-面积关系在大尺度(30 hm2)和中尺度(20 hm2)上适合采用逻辑斯蒂模型, 而小尺度(10 hm2)上适合用对数模型。在中尺度和大尺度上, 逻辑斯蒂模型优于对数模型和幂函数模型, 为最佳拟合模型, 说明在取样范围足够大且包括样地所有物种条件下, 逻辑斯蒂模型能够更好地描述物种和面积的关系[29]。幂函数模型拟合优度虽然在中尺度和大尺度上低于逻辑斯蒂模型, 但模型参数具有明确的生物学意义, 反映了研究样地中的α多样性(C值)(随机取样时α=4.82;巢式取样时α=7.18) 和β多样性(Z值)(随机取样时β=0.19;巢式取样时β=0.16)。本文结果中, 幂函数模型在不同尺度的拟合均未表现出理想的拟合效果。而在许多其他研究中, 小尺度、中尺度上幂函数模型拟合效果优于其他模型[17, 30]。这可能是因为本研究样地处于温带森林, 物种数相对较少且聚集程度相对更低, 而幂函数曲线趋于无限增长形式, 容易造成预测误差[25]。在小尺度上对数模型的拟合效果优于其他模型, 可能是因为小尺度上群落的环境条件及物种组成相似, 新物种数增加缓慢, 更适合用对数模型拟合[29-31]。
种-面积关系表现出的尺度效应与群落物种组成格局密切相关。Reed认为, 生境异质性影响了物种在不同空间尺度上的分布格局, 是影响种-面积关系的主要推动力[32-34]。小尺度上, 选取的样方之间生境差异表现明显, 故物种组成上也会出现较大的差异, 呈现出种-面积曲线的较大变异;而大尺度上, 生境异质性降低, 样本间差异小, 种-面积曲线更为稳定[16]。代力民等对二道白河河岸植物群落及森林群落对比研究也发现, 河岸带生境异质性强、物种丰富度高, 二者的种-面积曲线差异较大, 表现为河岸带群落的最小面积小于森林群落[35]。环境梯度是形成生境异质性的主要成分之一[36], 阔叶红松林样地海拔梯度变化大、坡度变化明显, 群落生境异质性高, 生境过滤作用强, 生态位相近的物种表现出相似的分布格局[37], 物种聚集程度高。随着研究尺度的增大, 出现的新物种数少, 大部分为稀有种或偶见种, 这种分布格局与生境异质性显著相关[38]。
另外, 由于研究林分演替阶段不同, 群落空间格局显著不同, 影响不同尺度上的取样结果。姜俊等在蛟河42 hm2针阔混交林的研究也证明了种-面积关系具有尺度效应, 但与该结果相异的是小尺度和中尺度以幂函数模型最优, 在大尺度上以逻辑斯蒂模型最优, 造成两个研究结果差异的原因可能是, 处于演替中后期的阔叶红松林在物种组成以及物种丰富度增加速率上与演替中期的针阔混交林显著不同[7, 39], 张春雨等人的研究也表明不同演替阶段物种随取样面积增加的速率不同, 其中中后期的阔叶红松林的物种增加速率小于中期的针阔混交林[40]。
在蛟河30 hm2阔叶红松林样地中, 取样方法对种-面积关系影响显著, 其中随机取样优于巢式取样。两种取样方法构建的种-面积关系不同, 会影响对局域物种数预测的准确性[41]。随机取样时按设置样方的规格进行等概率的抽样, 样地中各区域被抽中的概率相等, 减少了因生境异质性、种子传播、物种萌生能力强等带来的物种聚集分布的情况。与随机取样相比, 巢式取样方法简单, 但同时小样方中的稀有种也出现在大样方, 容易高估物种数, 降低预测物种数的准确性。倪瑞强等对长白山森林群落的研究也表明, 在取样方法上随机取样优于巢式取样[42]。Collins等的研究也表明, 巢式样方的种-面积关系不适合外推估算大尺度的物种数量[43]。此外, 一些研究还表明种-面积关系也受到取样面积、样方数以及样方形状等影响[2, 35]。
本研究基于演替阶段中后期的阔叶红松林样地, 认为区域种-面积关系同时受研究尺度、取样方法的影响。在实际应用时, 要充分考虑研究区域的实际群落结构和样地内环境差异, 在相应的尺度上选取合适的取样方法, 从而利用种-面积关系对森林经营给予合理建议。
| [1] | Lomolino M V. Ecology's most general, yet protean1 pattern: the species-area relationship. Journal of Biogeography, 2000, 27(1): 17–26. DOI:10.1046/j.1365-2699.2000.00377.x |
| [2] | Palmer M W, White P S. Scale dependence and the species-area relationship. The American Naturalist, 1994, 144(5): 717–740. DOI:10.1086/285704 |
| [3] | Chave J, Muller-Landau H C, Levin S A. Comparing classical community models: theoretical consequences for patterns of diversity. The American Naturalist, 2002, 159(1): 1–23. DOI:10.1086/324112 |
| [4] | Zurlini G, Grossi L, Rossi O. Spatial-accumulation pattern and extinction rates of Mediterranean flora as related to species confinement to habitats in preserves and larger areas. Conservation Biology, 2002, 16(4): 948–963. DOI:10.1046/j.1523-1739.2002.99547.x |
| [5] | Pimm S L, Russell G J, Gittleman J L, Brooks T M. The future of biodiversity. Science, 1995, 269(5222): 347–350. DOI:10.1126/science.269.5222.347 |
| [6] | Thomas C D, Cameron A, Green R E, Bakkenes M, Beaumont L J, Collingham Y C, Erasmus B F N, de Siqueira M F, Grainger A, Hannah L, Hughes L, Huntley B, van Jaarsveld A S, Midgley G F, Miles L, Ortega-Huerta M A, Peterson A T, Phillips O L, Williams S E. Extinction risk from climate change. Nature, 2004, 427(6970): 145–148. DOI:10.1038/nature02121 |
| [7] | Lepš J, Štursa J. Species-area curve, life history strategies, and succession: a field test of relationships. Vegetatio, 1989, 83(1/2): 249–257. |
| [8] | 唐志尧, 乔秀娟, 方精云. 生物群落的种-面积关系. 生物多样性, 2009, 17(6): 549–559. |
| [9] | Connor E F, McCoy E D. The statistics and biology of the species-area relationship. The American Naturalist, 1979, 113(6): 791–833. DOI:10.1086/283438 |
| [10] | Tjørve E. Shapes and functions of species-area curves: a review of possible models. Journal of Biogeography, 2003, 30(6): 827–835. DOI:10.1046/j.1365-2699.2003.00877.x |
| [11] | Turner W R, Tjørve E. Scale-dependence in species-area relationships. Ecography, 2005, 28(6): 721–730. DOI:10.1111/eco.2005.28.issue-6 |
| [12] | Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees. PLoS Biology, 2006, 4(11): e344. DOI:10.1371/journal.pbio.0040344 |
| [13] | 饶米德, 冯刚, 张金龙, 米湘成, 陈建华. 生境过滤和扩散限制作用对古田山森林物种和系统发育β多样性的影响. 科学通报, 2013, 58(13): 1204–1212. |
| [14] | 王世雄. 黄土高原子午岭植物群落物种多样性的时空格局与过程[D]. 西安: 陕西师范大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10718-1014177265.htm |
| [15] | Williamson M, Gaston K J, Lonsdale W M. The species-area relationship does not have an asymptote. Journal of Biogeography, 2001, 28(7): 827–830. |
| [16] | 姜俊, 张春雨, 赵秀海. 吉林蛟河42hm2针阔混交林样地植物种-面积关系. 植物生态学报, 2012, 36(1): 30–38. |
| [17] | Fisher R A, Corbet A S, Williams C B. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology, 1943, 12(1): 42–58. DOI:10.2307/1411 |
| [18] | 臧婧. 河南宝天曼落叶阔叶林种-面积关系研究[D]. 哈尔滨: 东北林业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10225-1015655775.htm |
| [19] | Drakare S, Lennon J J, Hillebrand H. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecology Letters, 2006, 9(2): 215–227. DOI:10.1111/j.1461-0248.2005.00848.x |
| [20] | Scheiner S M. Six types of species-area curves. Global Ecology and Biogeography, 2003, 12(6): 441–447. DOI:10.1046/j.1466-822X.2003.00061.x |
| [21] | Arrhenius O. Species and area. Journal of Ecology, 1921, 9(1): 95–99. DOI:10.2307/2255763 |
| [22] | Gleason H A. Species and area. Ecology, 1925, 6(1): 66–74. DOI:10.2307/1929241 |
| [23] | Ugland K I, Gray J S, Ellingsen K E. The species-accumulation curve and estimation of species richness. Journal of Animal Ecology, 2003, 72(5): 888–897. DOI:10.1046/j.1365-2656.2003.00748.x |
| [24] | 王雨茜. 吉林蛟河老龄林群落特征及其与地形关系[D]. 北京: 北京林业大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10022-1013212047.htm |
| [25] | Ulrich W, Buszko J. Sampling design and the shape of species-area curves on the regional scale. Acta Oecologica, 2007, 31(1): 54–59. DOI:10.1016/j.actao.2006.03.005 |
| [26] | Grilli J, Azaele S, Banavar J R, Maritan A. Spatial aggregation and the species-area relationship across scales. Journal of Theoretical Biology, 2012, 313: 87–97. DOI:10.1016/j.jtbi.2012.07.030 |
| [27] | Condit R, Hubbell S P, Lafrankie J V, Sukumar R, Manokaran N, Foster R B, Ashton P S. Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots. Journal of Ecology, 1996, 84(4): 549–562. DOI:10.2307/2261477 |
| [28] | 郝占庆, 李步杭, 张健, 王绪高, 叶吉, 姚晓琳. 长白山阔叶红松林样地(CBS):群落组成与结构. 植物生态学报, 2008, 32(2): 238–250. |
| [29] | He F L, Legendre P. On species-area relations. The American Naturalist, 1996, 148(4): 719–737. DOI:10.1086/285950 |
| [30] | Archibald E E A. The specific character of plant communities: Ⅱ. A quantitative approach. Journal of Ecology, 1949, 37(2): 274–288. DOI:10.2307/2256609 |
| [31] | Williams C B. Area and number of species. Nature, 1943, 152(3853): 264–267. DOI:10.1038/152264a0 |
| [32] | Reed T. The number of breeding land bird species on British Islands. Journal of Animal Ecology, 1981, 50(2): 613–624. DOI:10.2307/4078 |
| [33] | 沈国春. 生境异质性和扩散限制对亚热带和热带森林物种多样性维持的作用[D]. 杭州: 浙江大学, 2010. http://cdmd.cnki.com.cn/Article/CDMD-10335-1011066742.htm |
| [34] | Guo Y L, Lu J M, Franklin S B, Wang Q G, Xu Y Z, Zhang K H, Bao D C, Qiao X J, Huang H D, Lu Z J, Jiang M X. Spatial distribution of tree species in a species-rich subtropical mountain forest in central China. Canadian Journal of Forest Research, 2013, 43(9): 826–835. DOI:10.1139/cjfr-2013-0084 |
| [35] | 代力民, 王青春, 邓红兵, 陈高, 王庆礼. 二道白河河岸带植物群落最小面积与物种丰富度. 应用生态学报, 2002, 13(6): 641–645. |
| [36] | Redon M, Bergès L, Cordonnier T. Effects of increasing landscape heterogeneity on local plant species richness: how much is enough?. Landscape Ecology, 2014, 29(5): 773–787. DOI:10.1007/s10980-014-0027-x |
| [37] | Debski I, Burslem D F R P, Palmiotto P A, Lafrankie J V, Lee H S, Manokaran N. Habitat preferences of Aporosa in two Malaysian forests: implications for abundance and coexistence. Ecology, 2002, 83(7): 2005–2018. DOI:10.1890/0012-9658(2002)083[2005:HPOAIT]2.0.CO;2 |
| [38] | Hubbell S P. Tree dispersion, abundance, and diversity in a tropical dry forest. Science, 1979, 203(4387): 1299–1309. DOI:10.1126/science.203.4387.1299 |
| [39] | 刘万德, 苏建荣, 李帅锋, 张志钧, 郎学东. 南亚热带季风常绿阔叶林不同演替阶段物种-面积关系. 应用生态学报, 2011, 22(2): 317–322. |
| [40] | 张春雨, 赵秀海, 赵亚洲. 长白山温带森林不同演替阶段群落结构特征. 植物生态学报, 2009, 33(6): 1090–1100. |
| [41] | Fattorini S. To fit or not to fit? A poorly fitting procedure produces inconsistent results when the species-area relationship is used to locate hotspots. Biodiversity and Conservation, 2007, 16(9): 2531–2538. DOI:10.1007/s10531-006-9029-6 |
| [42] | 倪瑞强. 长白山典型针阔混交林群落结构与动态研究[D]. 北京: 北京林业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324722.htm |
| [43] | Collins M D, Vázquez D P, Sanders N J. Species-area curves, homogenization and the loss of global diversity. Evolutionary Ecology Research, 2002, 4(3): 457–464. |