 2017, Vol. 37
2017, Vol. 37文章信息
- 韩其飞, 罗格平, 李超凡, 黄晓东
- HAN Qifei, LUO Geping, LI Chaofan, HUANG Xiaodong.
- 放牧对新疆草地生态系统碳源/汇的影响模拟研究
- Modeling the grazing effect of grassland on the carbon source/sink in Xinjiang
- 生态学报. 2017, 37(13): 4392-4399
- Acta Ecologica Sinica. 2017, 37(13): 4392-4399
- http://dx.doi.org/10.5846/stxb201603110430
-
文章历史
- 收稿日期: 2016-03-11
- 网络出版日期: 2017-02-23





2. 南京信息工程大学气象灾害预报预警与评估协同创新中心, 南京 210044;
3. 中国科学院新疆生态与地理研究所, 荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011;
4. 中国科学院南京土壤研究所, 南京 210008;
5. 兰州大学草地农业科技学院草地农业生态系统国家重点实验室, 兰州 730020
2. Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters, Nanjing University of Information Science & Technology, Nanjing 210044, China;
3. State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;
4. State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China;
5. State Key Laboratory of Grassland and Agro-ecology Systems, College of Pastoral Agricultures Science and Technology, Lanzhou University, Lanzhou 730020, China
干旱半干旱地区生态系统是陆地生态系统中最为脆弱的部分之一[1-5], 该地区分布着世界上绝大多数的牧场和天然草场。新疆是干旱半干旱生态系统分布较集中的区域之一, 草地面积5.7×107 hm2, 在全国位于第3位。近年来, 放牧活动的不断增强和气候变化的加剧, 使得新疆草地生态系统面临着巨大的社会和生态环境问题[6-7], 同时也影响着草地生态系统的结构与功能(生产力、碳储量和通量)[8-11]。
放牧作为一种高度复杂的干扰方式, 通过采食、践踏和排泄物影响植物群落生物量及结构特征和土壤理化性质, 从而影响生态系统的碳循环过程[12]。放牧通过改变牧草各器官之间固有的物质与能量分配模式来影响牧草的生长, 如放牧影响草地的根冠比, 使得地上部分的生长速度高于根系[13]。另外, 放牧改变种群动态。家畜的选择性采食能够改变牧草的竞争力。一般来说, 采食削弱被采食牧草种的竞争力, 增强未采食或少采食牧草种的竞争力[14-15]。然而过度放牧不但会破坏地表植被, 促使大气对土壤的直接影响增强, 增加地表蒸发, 减少土壤表层含水量, 损坏根系生存环境, 还能直接影响采食植物的叶片、繁殖器官和生长点等部分, 通过直接影响地上部分生长进而影响到地下部分根系的生长发育和生物量的形成及积累[13, 16]。
尽管放牧通常被认为对植物生长是不利的, 但一些研究也表明适度放牧可能促进草地的生长[15, 17-19], 即表现出补偿机制(compensatory mechanism)。有研究表明, 不同草地类型对于放牧的响应存在差异, 适度放牧可能促进天山北坡低山干旱草地的净初级生产力, 但对于森林草甸草原和高寒草甸草原的生产力均具有负面效应[20]。总之, 放牧是否促进草地植物的生长, 目前学术界还有许多争论, 放牧对草地碳平衡与碳动态的影响还具有很大的不确定性[21-22]。因此, 研究放牧对新疆草地生态系统碳源/汇的影响, 对于明确草地碳循环途径及其在区域碳循环中的作用具有重要的意义。
由于站点尺度直接观测法研究放牧对草地碳源/汇影响的局限性, 陆地生态系统模型成为研究气候变化和人类活动对生态系统结构与功能影响普遍使用的方法[20, 23-24]。其中Biome-BGC grazing模型在新疆天山北坡得到成功应用, 模拟了放牧对不同海拔草地生态系统净初级生产力的影响[20]。
因此, 本文基于空间化的Biome-BGC grazing模型, 1) 模拟新疆草地生态系统碳动态特征, 分析其碳/源汇功能和强度的时空分布;2) 量化放牧过程对新疆草地生态系统碳源/汇的贡献。本研究将有助于揭示区域草地生态系统对气候变化和放牧的响应格局与过程。
1 材料与方法 1.1 研究区概况新疆地处欧亚大陆腹地, 属于典型的温带大陆干旱性气候, 光热资源充足, 年平均气温9—12℃, 降水量稀少, 北疆降水为150—200mm, 南疆在100mm以下。而蒸发量则相反, 北疆为1500—2300mm, 南疆为2100—3400mm[25-26], 在全球变化和人类活动的干扰下, 新疆的气候和生态已经发生了显著的变化[27]。20世纪80年代以来出现了暖干向暖湿的转型变化[28-29]。新疆草地总面积5.7×107 hm2, 占全疆土地总面积的34%[6]。不同草地类型按照海拔梯度进行划分, 包括高山草甸(Alpine meadow, AM, 2700—3500m)、中山森林草地(Mid-mountain forest-meadow, MMFM, 1650—2700m)、低山干草原(Low-mountain dry grassland, LMDG, 650—1650m)和平原荒漠草原(Plain desert grassland, PDG, <650m)[20]。
1.2 放牧数据研究所用的放牧强度数据Gridded Livestock of the World (简称GLW)来源于全球畜牧信息系统global livestock information system (简称GLIS)数据。GLIS数据是Food and Agriculture Organization of the United Nations (简称FAO)根据全球牲畜数目采样点及环境数据差值产生, 空间分辨率为5km。GLW数据中包含了主要的牲畜类型:牛、水牛、绵羊、山羊、猪和家禽类。根据中华人民共和国农业部提供的“标准羊”换算:用1只绵羊为1个标准羊单位, 1头牛为6个标准羊单位, 一头水牛为6.5个标准羊单位, 一只山羊为0.9个标准羊单位(http://www.chinaforage.com/standard/zaixuliang.htm)(图 1)。

|
| 图 1 1979—2007年新疆草地放牧率空间分布 Fig. 1 Distribution of averaged grazing intensity between 1979 and 2007 |
Biome-BGC模型是一个用来估算陆地生态系统日尺度碳、氮、水储存及通量的生态系统过程模型。输出包括总初级生产力(GPP)、净初级生产力(NPP)、净生态系统碳交换量(NEE)等。Biome-BGC grazing耦合了落叶方程模拟放牧对草地生态系统碳循环的影响, 该模型已成功的用于半干旱草原生态系统[20]。落叶方程将动物采食的碳(Dr)分为5个部分:动物的呼吸作用(占50%)、甲烷的生产(占3%)、肉的生产、尿、和粪便(占30%)。在考虑放牧活动干扰的情景下(Dr>0), 地上净初级生产力(ANPP)等于地上生物量增量与牲畜采食量之和。本文未对草地的可食性进行区分, 且由于Biome-BGC模型将草本植物划分为细根和叶两个部分, 本文假设牲畜仅采食草的叶。
因此, 在该模型中, NEE的定义为:
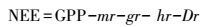
|
(1) |
式中, mr为维持呼吸, gr为生长呼吸, hr为异养呼吸, Dr为凋落速率。
模型参数采用在新疆天山北坡不同海边梯度校正过的参数[20], 每个格点的分辨率为10km×10km。为了分析气候变化与放牧对碳源/汇的影响, 本文设计了2个情景:仅气候变化(Climate change, CLIM)、气候变化和放牧共同作用(Climate change and Grazing, CLIMGRA), 则放牧对碳源/汇的影响由二者之差表示。
1.5 模型验证本文利用在东天山奇台、天山北坡不同海拔高度和平原荒漠草原区设置的围栏和观测点, 进行了生物量、土壤水分、质地和养分的观测和分析结果, 分放牧和无放牧两种情景对模型进行验证。重点对高山草甸、中山森林草地、低山干草原和平原荒漠草原年地上净初级生产力分别进行了验证。另外, 由于缺少足够覆盖新疆的实测数据, 本文部分数据通过搜集已发表的文献数据获得。同时, 为了验证模型模拟碳通量的能力, 我们在AmeriFlux网站下载了美国Kendall草原的涡度相关数据(ftp://cdiac.ornl.gov/pub/ameriflux/data), 从而验证模型输出NEE的精度。
1.6 放牧率阈值分析为了研究碳源/汇对于放牧率的响应, 从而确定不同草场类型的适宜放牧率阈值。本文首先将研究区按照牧场类型分区;然后在分区内选取放牧后(CLIMGRA)较无放牧(CLIM)情景下NEE变化为正值的格点;最后, 利用这些格点作为掩模, 统计该部分格点覆盖下牧场的平均放牧率, 作为该类型牧场的适宜放牧率阈值。
2 结果与分析 2.1 模型验证将Biome-BGC grazing模拟得到的净初级生产力(NPP)与实测值进行比较。由图 2可以看出, 围栏及放牧条件下平均NPP实测值与模拟值具有较好的吻合性(R2=0.90, P<0.001, n=41), 说明Biome-BGC grazing模型可较好模拟新疆植被NPP。
| 项目县 Study area | 样地 Sample plot | 经纬度 Longitude/latitude | 海拔 Altitude | 措施类型 Grazing type | 草地类型 Grassland type | 备注 Remarks |
| 奇台县 Qitai | 西北旱地1 | 43°46.16′N 89°24.27′E | 1339 | 围栏、禁牧 | 低山干草原 | 2006年围栏 |
| 西北旱地2 | 43°46.06′N 89°24.38 E | 1350 | 围栏 | 低山干草原 | 2006年围封, 后破坏, 2008年5月重围 | |
| 西北旱地3 | 43°46.16′N 89°24.49′E | 1339 | 自由放牧 | 低山干草原 | ||
| 吉布库牧场 | 43°39.70′N 89°29.32′E | 1608 | 低山干草原 | 醉马草毒害草调查 | ||
| 小水山 | 43°35.99′N 89°42.25′E | 2014 | 中山森林草地 | 酸模退化草地, | ||
| 新户河 | 43°33.33′N 89°45.49′E | 2148 | 围栏 | 中山森林草地 | 2006年围栏, 以三叶草、早熟禾、千叶耆为优势种 | |
| 麻沟梁 | 43°34.59′N 89°45.22′E | 2032 | 围栏 | 中山森林草地 | 2007年围栏, 25个鼠洞/10m2 | |
| 芨芨湖 | 44°17.13′N 90°05.33′E | 717.6 | 围栏 | 荒漠草原 | 2004年围栏, 以梭梭、骆驼刺为优势种 | |
| 三工河流域 Sangong | 43°57.60′N 88°04.20′E | 1296 | 围栏 | 低山干草原 | 2004—2009年采集土壤质地、比叶面积、生物量、根冠比信息 | |
| River Basin | 43°55.20′N 88°14.40′E | 2376 | 围栏 | 中山森林草地 | ||
| 43°52.80′N 88°16.20′E | 3113 | 围栏 | 高山草甸 |

|
| 图 2 实测NPP与模拟NPP值对比 Fig. 2 Comparisons of estimated NPP and observed NPP for grassland in Xinjiang Province |
由图 3可以看出, 通量模拟结果从总体变化趋势与涡度相关数据吻合, 均表现为单峰型季节变化特征。GPP的相关系数(R2)为0.82, NEE的相关系数为0.55, 两者的均方根误差分别为1.36 g C m-2 d-1和0.84 g C m-2 d-1。

|
| 图 3 实测GPP、NEE与模拟结果对比 Fig. 3 Comparison of predicted and measured GPP, NEE |
图 4给出了新疆草地的碳源/汇(NEE)的时间变化图。从图 4可以看出, 1979—2011年新疆草地生态系统表现为碳源, 累积释放的碳量为0.38Pg C (图 4)。由时间变化趋势分析, 29年来新疆的NEE呈明显波动下降趋势, 1983年、1988年和2003年NEE出现峰值, 这与该区降水峰值出现的年份较吻合(图 4)。1990年后, NEE整体下降趋势最为明显, 产生这种现象的主要原因是放牧率的增加, 1990年前后的放牧率平均值分别为0.16头标准羊/hm2和0.76头标准羊/hm2(图 4, 图 6)。

|
| 图 4 1979—2007年新疆草地年均NEE累积NEE变化图 Fig. 4 Interannual patterns of annual averaged NEE and cumulatived NEE of Xinjiang grassland from 1979 to 2007 |
另外, 新疆草地碳源/汇的变化具有明显的区域特征(图 5)。受区域水热条件的制约, 草地植被大体上呈现出由北向南依次出现高山与亚高山草甸、平原草地、草甸、荒漠草地和高山与亚高山草地, 其NEE也逐渐由0—0.6gCm-2a-1到-10—-300gCm-2a-1, 即由碳吸收转换为碳排放。最大的碳源是高山森林草甸, 平均每年释放45gCm-2a-1。

|
| 图 5 1979—2007年新疆草地平均NEE空间分布图 Fig. 5 Distribution of the estimated carbon source/sink in Xinjiang from 1979 to 2007 |
由图 6可以看出, 在无放牧情景下(CLIM), 1979—2011年新疆草地生态系统吸收和释放的碳量相当, 平均每年释放约0.0005Pg C。而在放牧情境下(CLIMGRA), 平均每年释放的碳量为0.013 Pg C a-1, 即在过去29年, 放牧导致新疆释放0.37Pg碳。与无放牧情景情况相比较, 放牧后NEE呈现出随着放牧梯度的增加而降低的趋势, 随放牧时间的延续, 当载畜率超过0.4头标准羊/hm2时, NEE开始显著降低。这表明对于草地生态系统来说, 随着放牧梯度的增加, 降低了草地的初级生产力, 从而使生态系统固定的碳减少。

|
| 图 6 1979—2007年放牧前后新疆草地NEE及放牧率变化图 Fig. 6 Interannual patterns of mean NEE of Xinjiang grassland with and without grazing from 1979 to 2007 |
放牧对NEE变化的区域影响如图 7所示, 大部分地区的NEE在放牧后减少, 即放牧增加了碳的排放。但是, 除冬季牧场外, 其他草场在适宜的放牧率条件下, 都表现出了补偿机制, 平均阈值放牧率约为(0.24±1.63) 头标准羊/hm2, 不同草场类型的阈值放牧率如表 2所示。本研究表明, 春秋牧场相比于其他牧场能有效地补偿因严重采食而失去的地上组织, 主要是因为该类型牧场主要分布在低山干草原(LMDG, 650—1650m), 放牧能够一定程度上降低水分胁迫对干旱草原生态系统净初级生产的负面效应。

|
| 图 7 放牧前后新疆草地NEE变化 Fig. 7 The difference of NEE before and after grazing scenarios |
| 草场类型 Grassland type | 平均放牧率 Grazing intensity | 标准差 Standard deviation |
| 春季牧场Spring pasture | 0.37 | 1.80 |
| 秋季牧场Autumn pasture | 0.38 | 2.81 |
| 全年牧场yearlong grazing | 0.12 | 0.34 |
| 夏季牧场Summer pasture | 0.17 | 0.38 |
| 冬季牧场Winter pasture | - | - |
本研究实现了Biome-BGC模型在新疆区域尺度的应用, 突破了由于区域试验站点有限、草地类型多样而无法实现空间动态评估与预估的瓶颈。但是研究中仍然存在一些问题:1) 放牧模型中许多关于理想条件的假设可能与实际情况不符, 从而导致模拟结果的失真。例如在放牧模型中, 未对植被物种的可食性进行区分, 从而高估了动物的采食能力;另外, 未考虑放牧对物种多样性变化、植被演替因素的影响等均可导致模型的不确定性。2) 模型预测的准确程度受到诸多因素的影响。涉及群落的一些生理生态参数的精确参数化较难, 相应实验研究也比较少;模型是在假定植被组成不变的前提下得出的模拟结果;随着实验研究的深入和相应数据源的增多, 将可以通过改进模型结构和参数化过程更真实全面地了解生态系统过程和结构对气候变化的响应和适应。3) 气象数据的空间分辨率较大, 相对于站点水平的研究还不能一一对应, 导致模拟结果存在不确定性。
3.2 放牧对新疆草地碳源/汇的影响放牧管理影响下草原碳循环和分布的生态过程还没有被完全认知, 从已有的文献中很难得出放牧管理与碳源/汇之间明确的关系。本研究结果表明, 尽管在新疆的大部分地区, 放牧导致草地碳释放增加, 但是当放牧率低于一定阈值时, 出现了补偿生长现象。影响牧草生长发育的因素有放牧制度、放牧家畜的种类、放牧强度、放牧时期、放牧周期和草地的植被与土壤特征[30]。20世纪40年代植物界就有补偿现象的报道, 董世魁等[19]研究发现, 重牧状态(70%的采食率)下, 草群盖度较低, 叶片截光量少, 牧草的补偿性再生生长弱, 草地牧草的现存量较低。中牧及轻牧状态(<50%的采食率)下, 草地的补偿作用较为明显。汪诗平等[31]通过研究发现, 当放牧率较低时, 如其研究的1.33头标准羊/hm2的放牧率, 表现出较高的超补偿性生长, 但随着放牧率的增大, 地上净初级生产力的补偿性生长随着降低。本文的研究发现, 新疆草地的在平均放牧率低于0.24头标准羊/hm2时, 可以促进草地碳固定, 但在历史实际的放牧强度下, 放牧是新疆草地生态系统碳释放的主要原因。
但是, 草地群落对放牧的补偿反应除了通过可食牧草的快速生长, 以补偿草地群落的功能损失之外, 还可能通过毒害草繁衍生长来实现[32]。虽然该种补偿形式同样可以增加草地的生产力, 但增加部分多为家畜不可采食的成分, 是一种功能上的冗余。因此, 在后续的研究中, 可以通过改进Biome-BGC grazing放牧模型, 增加表征种间竞争的“植被演替”模块, 通过模拟物种的竞争, 深入研究草地群落对放牧的补偿反应。
4 结论本文利用改进的Biome-BGC grazing模型对1979—2007年新疆草地生态系统长期碳源/汇动态(NEE)进行了模拟和验证, 分析了放牧活动对新疆草地生态系统碳动态的影响。结果表明, 1979—2007年新疆草地生态系统为碳源, 释放碳的总量为0.38PgC, 其中由放牧导致的碳释放为0.37PgC。当平均放牧率小于0.24头标准羊/hm2时, 放牧能够促进草地碳固定。本研究从增加碳汇的角度为草地适应性管理提供了重要的理论依据, 新疆草地通过合理控制放牧梯度能够达到增加草地固碳潜力, 实现碳增汇的目的。
| [1] | Letnic M. Dispossession, degradation and extinction:environmental history in arid Australia. Biodiversity & Conservation, 2000, 9(3): 295–308. |
| [2] | Pringle H J R, Watson L W, Tinley K L. Landscape improvement, or ongoing degradation-reconciling apparent contradictions from the arid rangelands of Western Australia. Landscape Ecology, 2006, 21(8): 1267–1279. DOI:10.1007/s10980-006-0015-x |
| [3] | Qi S Z, Luo F. Water environmental degradation of the Heihe River basin in arid Northwestern China. Environmental Monitoring and Assessment, 2005, 108(1): 205–215. |
| [4] | Jabbar M T, Chen X L. Land degradation due to salinization in arid and semi-arid regions with the aid of geo-information techniques. Geo-Spatial Information Science, 2008, 11(2): 112–120. DOI:10.1007/s11806-008-0013-z |
| [5] | Beyene S T. Rangeland degradation in a semi-arid communal savannah of swaziland:long-term dip-tank use effects on woody plant structure, cover and their indigenous use in three soil types. Land Degradation & Development, 2015, 26(4): 311–323. |
| [6] | 赵万羽, 李建龙, 齐家国, 刘旭. 新疆草地生态安全问题、现状与对策分析. 干旱区研究, 2005, 22(1): 45–50. |
| [7] | 柴军. 新疆牧民生产决策行为与草地退化问题研究[D]. 北京: 中国农业科学院, 2008. |
| [8] | Wang Y H, Zhou G S, Jia B R. Modeling SOC and NPP responses of meadow steppe to different grazing intensities in Northeast China. Ecological Modelling, 2008, 217(1/2): 72–78. |
| [9] | Riedo M, Gyalistras D, Fuhrer J. Net primary production and carbon stocks in differently managed grasslands:simulation of site-specific sensitivity to an increase in atmospheric CO2 and to climate change. Ecological Modelling, 2000, 134(2/3): 207–227. |
| [10] | Schaldach R, Wimmer F, Koch J, Volland J, Geißler K, Köchy M. Model-based analysis of the environmental impacts of grazing management on Eastern Mediterranean ecosystems in Jordan. Journal of Environmental Management, 2013, 127: S84–S95. DOI:10.1016/j.jenvman.2012.11.024 |
| [11] | Piñeiro G, Paruelo J M, Oesterheld M, Jobbágy E G. Pathways of grazing effects on soil organic carbon and nitrogen. Rangel and Ecology and Management, 2010, 63(1): 109–119. DOI:10.2111/08-255.1 |
| [12] | 侯扶江, 杨中艺. 放牧对草地的作用. 生态学报, 2006, 26(1): 244–264. |
| [13] | 张雪, 乌云娜, 林璐, 赵晨羽, 王雪婷, 白佳忆. 放牧梯度上草原植被——土壤系统碳截存特征. 中国沙漠, 2013, 33(6): 1789–1795. DOI:10.7522/j.issn.1000-694X.2013.00266 |
| [14] | 宋彦涛. 松嫩草地植物功能生态学研究[D]. 长春: 东北师范大学, 2012. |
| [15] | 汪诗平, 李永宏, 王艳芬, 陈佐忠. 不同放牧率对内蒙古冷蒿草原植物多样性的影响. 植物学报, 2001, 43(1): 89–96. |
| [16] | 王仁忠, 李建东. 羊草草地放牧退化演替中种群消长模型的研究. 植物生态学报, 1995, 19(2): 170–174. |
| [17] | 张璐璐, 周晓松, 李英年, 袁芙蓉, 樊瑞俭, 朱志红. 刈割、施肥和浇水对矮嵩草补偿生长的影响. 植物生态学报, 2011, 35(6): 641–652. |
| [18] | 朱志红, 席博, 李英年, 臧岳铭, 王文娟, 刘建秀, 郭华. 高寒草甸不同生境粗喙薹草补偿生长研究. 植物生态学报, 2010, 34(3): 348–358. |
| [19] | 董世魁, 江源, 黄晓霞. 草地放牧适宜度理论及牧场管理策略. 资源科学, 2002, 24(6): 35–41. |
| [20] | 周德成, 罗格平, 韩其飞, 尹昌应, 李龙辉, 胡玉昆. 天山北坡不同海拔梯度山地草原生态系统地上净初级生产力对气候变化及放牧的响应. 生态学报, 2012, 32(1): 81–92. |
| [21] | 孔玉华, 姚风军, 鹏爽, 刘艳, 董文轩, 白龙. 不同利用方式下草地土壤碳积累及汇/源功能转换特征研究. 草业科学, 2010, 27(4): 40–45. |
| [22] | 何念鹏, 韩兴国, 于贵瑞. 内蒙古放牧草地土壤碳固持速率和潜力. 生态学报, 2012, 32(3): 844–851. |
| [23] | 李东. 基于CENTURY模型的高寒草甸土壤有机碳动态模拟研究[D]. 南京: 南京农业大学, 2011. |
| [24] | 张存厚. 内蒙古草原地上净初级生产力对气候变化响应的模拟[D]. 呼和浩特: 内蒙古农业大学, 2013. |
| [25] | 丹利, 季劲钧, 马柱国. 新疆植被生产力与叶面积指数的变化及其对气候的响应. 生态学报, 2007, 27(9): 3582–3592. |
| [26] | 杨红飞, 刚成诚, 穆少杰, 章超斌, 周伟, 李建龙. 近10年新疆草地生态系统净初级生产力及其时空格局变化研究. 草业学报, 2014, 23(3): 39–50. DOI:10.11686/cyxb20140305 |
| [27] | 陈曦, 罗格平, 夏军, 周可法, 娄少平, 叶民权. 新疆天山北坡气候变化的生态响应研究. 中国科学D辑:地球科学, 2004, 34(12): 1166–1175. |
| [28] | 何清, 杨青, 李红军. 新疆40a来气温、降水和沙尘天气变化. 冰川冻土, 2003, 25(4): 423–427. |
| [29] | 施雅风, 沈永平, 胡汝骥. 西北气候由暖干向暖湿转型的信号、影响和前景初步探讨. 冰川冻土, 2002, 24(3): 219–226. |
| [30] | 朱玲玲, 戎郁萍, 王伟光, 马磊. 放牧对草地生态系统CO2净气体交换影响研究概述. 草地学报, 2013, 21(1): 3–10. DOI:10.11733/j.issn.1007-0435.2013.01.001 |
| [31] | 汪诗平, 王艳芬, 李永宏, 陈佐忠. 不同放牧率对草原牧草再生性能和地上净初级生产力的影响. 草地学报, 1998, 6(4): 275–281. DOI:10.11733/j.issn.1007-0435.1998.04.006 |
| [32] | 赵彬彬, 牛克昌, 杜国祯. 放牧对青藏高原东缘高寒草甸群落27种植物地上生物量分配的影响. 生态学报, 2009, 29(3): 1596–1606. |