 2017, Vol. 37
2017, Vol. 37文章信息
- 王鑫, 杨德刚, 熊黑钢, 柳妍妍, 安尼瓦尔·买买提
- WANG Xin, YANG Degang, XIONG Heigang, LIU Yanyan, ANWAR Mohammat.
- 新疆干旱区4种柽柳灌丛碳贮量特征
- Carbon storage characteristics of four different Tamarix L. shrubs in the arid land of Xinjiang Province
- 生态学报. 2017, 37(13): 4384-4391
- Acta Ecologica Sinica. 2017, 37(13): 4384-4391
- http://dx.doi.org/10.5846/stxb201603070403
-
文章历史
- 收稿日期: 2016-03-07
- 网络出版日期: 2017-02-23





2. 新疆大学资源与环境科学学院, 乌鲁木齐 830046;
3. 中国科学院大学, 北京 100049;
4. 中国科学院新疆分院, 乌鲁木齐 830011
2. Resource and Environmental Science Institute of Xinjiang University, Urumqi 830046, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Xinjiang Branch, Chinese Academy of Sciences, Urumqi 830011, China
陆地生态系统的碳库包括植物和土壤两部分, 植被碳库和土壤碳库碳贮存约为大气碳库的3倍, 对气候变化和生态系统碳循环具有重要的影响[1-2]。在干旱地区, 灌丛是一种广泛分布的植被类型, 其种类繁多、生命力强、适生范围宽等特点, 在群落演替、区域生态环境保护等方面具有极其重要的作用[3]。在全球气候变化和人为因素影响下, 草地灌丛化、荒漠化日益显著[4-5], 灌丛的固碳能力和潜力研究也日益受到重视, 前人研究中, 灌丛入侵能够增加土壤有机碳储量, 引起区域碳储量的变化[6-7], 并且干旱区灌丛土壤有机碳的分布、质量和周转特征, 也影响了这一地区土壤结构、功能特性[8]。目前, 国内外对森林[9-10]、草地[11-13]的植被和土壤碳贮量研究较多, 对灌丛的碳贮量研究较少, 尤其是生物量碳和土壤有机碳一起研究的较为鲜见。
柽柳属植物对干旱生境的高度适应与极强的防风固沙能力, 是过渡带植物群落的主要建群种之一, 在干旱区生态系统结构稳定, 维护干旱区生态系统平衡中具有重要地位[14-15]。目前, 国内关于柽柳的研究主要集中于生物量[14-17]、能量平衡[18]、化学计量特征[19-20]、耗水量[21-23]等方面的研究, 关于柽柳灌丛本身有机碳贮量及其土壤有机碳贮量的研究相对较少。不同种的柽柳抗旱能力和抗旱机理不同[21], 对环境变化的响应也不一样[22-23], 本文通过对4种柽柳灌丛群落碳贮量以及土壤有机碳贮量进行调查, 分析探讨不同柽柳灌丛碳贮量差异及其与环境因子的关系, 旨在为进一步研究干旱区群落演替过程中碳收支以及生态恢复过程中的植被类型选择提供参考依据。
1 研究区域与方法 1.1 研究区概况与样地选择新疆地处73°40′—96°18′E, 34°25′—48°10′ N之间, 面积约占全国1/6。处于亚欧内陆干旱区, 受西风环流、北冰洋高纬气团以及印度洋暖湿气流的影响, 形成显著区别于非洲、美洲和大洋洲热带荒漠的水热组合区域, 年均温南疆平原10—13℃, 北疆平原低于10℃, 年降水量平均约为150 mm, 降水量北疆多于南疆, 西部多于东部。新疆是我国典型的干旱荒漠区, 全疆土地面积有79.59×l04 km2为荒漠化土地, 集中于塔里木盆地和准噶尔盆地, 主要灌丛植被类型面积有5×l04 km2。其中柽柳灌丛以柽柳为主要优势种, 单丛生长或多丛连片生长, 伴生有梭梭(Haloxylon ammodendron), 琵琶柴(Reaumuria songonica (PalL) Maxim.), 骆驼蓬(Peganum harmala L.), 沙拐枣(Calligonum arborescens Litv.), 麻黄(Ephedra sinica Stapf), 霸王(Sarcozygium xanthoxylon Bunge), 盐生草(Halogeton glomeratus)等物种。
本研究依据电子版《新疆植被图》(新疆遥感与地理信息系统重点实验室提供)提取了分布于南北疆荒漠区的多枝、多花、刚毛和沙生4种柽柳灌丛类型(沙生柽柳北疆未有分布), 根据不同类型灌丛所在地设置样地(图 1), 每个样地100 m×100 m大小。

|
| 图 1 4种不同柽柳灌丛类型取样点 Fig. 1 Sampling point of 4 kinds of Tamrix L. |
在每个样地内随机选取3个样方, 每个样方10 m×10 m, 记录样方内出现的柽柳株数, 测算柽柳盖度, 采用收获法测定柽柳地上生物量, 室内风干称重。每个样方内随机挖取3个1 m×1 m的样坑按照100 cm的深度进行柽柳根系取样, 取出根系带回室内洗净烘干后测定地下生物量, 并分为0—10、10—20、20—30、30—40、40—50、50—100 cm的5个土壤层, 每层土壤取3个重复装进铝盒带回室内烘干法测定含水量, 取3个重复用密封塑料袋带回室内, 分析土壤养分、pH值和电导率, 土壤有机质采用重铬酸钾容量法, 全N采用半微量开氏法, 土壤pH值采用比色法, 电导率采用浸提法测定, 测定分析由中国科学院新疆生态与地理研究所土壤分析实验室完成;环刀法采集土样测定容重。当土层厚度达不到1 m时, 则均挖至母质层, 记录土壤深度, 在后期的数据处理时, 根据实际土层厚度计算土壤有机碳的密度。本项调查于2013年的9月—10月完成。
1.3 数据分析4种柽柳灌丛的面积数据来自《新疆植被图》电子版中的土壤面积, 《新疆植被图》电子版由空间数据库和属性数据库两部分组成。属性数据库中的基础数据主要来自于《新疆植被》、《新疆土壤》以及各市县土壤志等。空间数据库由新疆1:50000土壤图经扫描、配准、矢量化、拼接等处理而成, 并且结合1:50000的地形图作为工作底图, 同时对地形图进行矢量化, 再由ArcGIS软件将其生成三维地图, 由三维地图统计出面积作为计算土壤有机碳贮量的面积依据。
本研究中参考Shi[24]和Gong[25]文献, 植物碳含量系数统一取值为0.5, 生物量碳密度计算公式(1) 如下:

|
(1) |
式中, BBCDj代表第j 种灌丛类型的生物量碳平均密度(Mg C/hm2);Bj代表植物j的生物量。
土壤剖面的有机碳密度计算公式(2) 如下:
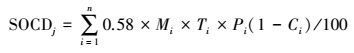
|
(2) |
式中, SOCDj为第j 种植被类型的土壤有机碳平均密度(Mg C/hm2);0.58为Blemmelen系数, 将有机质浓度转化为有机碳浓度;Pi, Mi, Ti, Ci分别表示第i层土壤容重(g/cm3)、有机质浓度(%)、土层厚度(cm)、>2 mm的砾石含量(%);n代表层数。
生物量碳贮量计算公式(3) 如下:
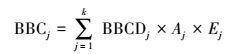
|
(3) |
式中, BBCj为第j 种灌丛类型的生物量碳贮量(Mg); Aj为第j 种灌丛类型的面积(hm2);Ej为第j 种灌丛盖度(%);k为样地数。
土壤有机碳贮量计算公式(4) 如下:
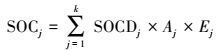
|
(4) |
式中, SOCj为第j 种灌丛类型的土壤有机碳贮量(Tg);SOCDj为第j 种土壤类型的土壤有机碳平均密度(Mg C/hm2);Aj为第j 种灌丛类型的面积(hm2);Ej为第j 种灌丛盖度(%);k为样地数。
本研究利用SPSS 16.0和SigmaPlot 10.0软件对实验数据进行分析处理、相关性分析及作图。
2 结果与分析 2.1 不同柽柳灌丛生物量有机碳密度对4种不同柽柳灌丛类型的生物量有机碳平均值进行了统计(图 2), 4种灌丛地上生物量碳密度在0.31—0.94 Mg C/hm2之间, 多枝柽柳灌丛地上生物量碳密度最低, 多花柽柳灌丛地上生物量碳密度最高;从地下生物量碳密度来看, 4种柽柳灌丛0—100 cm地下植物碳密度在0.28—1.49 Mg C/hm2之间, 多枝柽柳灌丛地下生物量碳密度最低, 刚毛柽柳灌丛地下生物量碳密度最高;4种柽柳总生物量碳密度由大到小的顺序是刚毛>多花>沙生>多枝, 刚毛柽柳碳密度能达到多枝柽柳的4倍。综合来看, 4种柽柳灌丛除了多枝柽柳, 其他3种灌丛地下生物量碳密度都大于地上生物量碳密度, 刚毛柽柳的根冠比最大, 为1.71, 多花柽柳为1.25, 沙生柽柳为1.21, 多枝柽柳最小, 仅为0.92。说明在干旱区, 灌丛植物通过增加地下生物量的策略来获取更多水分和养分以供生长所需, 而不同的柽柳类型, 也显现出不同的生长策略, 水分条件不同也导致柽柳根系垂直分布的差异。本研究取样深度为100 cm, 刚毛柽柳地下生物量碳密度明显大于其他, 而多枝柽柳根系可能分布更深。

|
| 图 2 4种不同柽柳生物量有机碳密度 Fig. 2 Biological biomass carbon density of 4 kinds of Tamrix L. |
数据分析表明(图 3), 4种柽柳灌丛中, 0—100 cm土壤有机碳密度在(15.00±1.40) —(63.49±7.51) Mg C/hm2之间, 土壤有机碳密度由大到小的顺序是多花>刚毛>沙生>多枝, 多花和沙生柽柳灌丛土壤有机碳密度的最大值出现在表层0—10 cm, 且随着土层深度的增大呈现降低趋势。多枝和刚毛柽柳灌丛土壤有机碳密度的最高值出现在土层深度为10—20 cm处。0—10 cm土层土壤有机碳含量在13%—43.7%之间, 地下1 m内, 0—50 cm土壤有机碳密度所占比例为82.3%—96.4%之间, 其中刚毛柽柳为96.4%, 沙生柽柳为82.3%;而50—100 cm土层土壤有机碳密度仅占3.6%—17.7%之间。总体而言, 不同柽柳灌丛0—50 cm土层深度的土壤有机碳密度高于50—100 cm土层深度土壤有机碳的密度。另外, 因为柽柳的根系较深, 1 m以下的土壤有机碳密度需要进一步深入研究。与生物量碳相比, 4种柽柳土壤有机碳密度最大的是多花柽柳, 最小的是多枝柽柳, 土壤有机碳密度分布情况基本与生物量碳分布一致, 说明柽柳灌丛土壤有机碳来源主要来自生物量。

|
| 图 3 4种不同柽柳灌丛土壤有机碳密度的垂直分布特征 Fig. 3 Vertical distribution characteristics of soil organic carbon density of 4 kinds of Tamrix L. |
根据表 1, 4种柽柳灌丛中, 总有机碳密度最大的是多花柽柳灌丛((65.60±7.83) Mg C/hm2), 其次是刚毛柽柳灌丛((63.16±5.94) Mg C/hm2), 最小的是多枝柽柳灌丛((15.59±1.48) Mg C/hm2)。生物量碳密度最大的是刚毛柽柳灌丛, 其值为(2.40±0.26) MgC/hm2, 土壤有机碳密度最大的是多花柽柳灌丛, 其值为(63.49±7.51) Mg C/hm2。4种柽柳灌丛土壤有机碳密度平均是生物量碳密度的24倍, 其中, 多花柽柳土壤有机碳密度是生物量碳密度的30倍, 沙生柽柳比值为14倍。与前人结果相比[1-2, 4, 7], 荒漠灌丛土壤有机碳密度比生物量碳密度要高出很多, 原因在于荒漠植物的生物量较小, 而土壤有机碳的积累经历了一个长期的积累过程。沙生柽柳主要分布于塔克拉玛干沙漠边缘的流动沙丘上, 在北疆固定与半固定沙丘上反而没有, 其土壤有机碳积累量与积累速度相较其余3种柽柳类型要低。从4种不同柽柳灌丛在新疆的分布面积来看, 分布面积最大的柽柳灌丛是刚毛柽柳, 其次是多枝柽柳, 最小的是多花柽柳;盖度最大的是多枝柽柳, 最小的是多花柽柳。由于有机碳密度、分布面积和盖度的不同, 总有机碳贮量以刚毛柽柳最高, 为(56.96±5.36) Tg, 多花柽柳最低, 为(0.28±0.03) Tg, 各柽柳灌丛有机碳贮量差异显著。
| 样地 Sample plots | 群落类型 Community types | 生物量碳密度/(Mg C/hm2) Biological biomass carbon density | 土壤有机碳密度 Soil carbon density/(Mg C/hm2) | 面积 Area/(104m2) | 盖度/% Coverage | 有机碳贮量/Tg Carbon storage | |
| 地上 Aboveground | 地下 Underground | ||||||
| 1 | 多花柽柳 Tamarix hohenackeri | 0.94±0.14a | 1.17±0.18b | 63.49±7.51a | 5.1 | 8.4 | 0.28±0.03d |
| 2 | 多枝柽柳 Tamarix ramosissima | 0.31±0.05c | 0.28±0.03d | 15.00±1.40c | 443.7 | 18.2 | 12.59±1.20b |
| 3 | 刚毛柽柳 Tamarix hispida | 0.87±0.05ab | 1.49±0.21a | 60.80±5.68a | 842.8 | 10.7 | 56.96±5.36a |
| 4 | 沙生柽柳 Tamarix taklamakanensis | 0.89±0.06ab | 1.08±0.16bc | 27.68±2.06b | 48.4 | 15.1 | 2.17±0.17c |
根据表 2可以看出, 土壤有机碳与土壤含水量、有机质、全N含量和电导率呈极显著正相关(P<0.01), 多元线性回归方程为:
| 项目 Items | 土壤含水量 Soil moisture | 有机质 Organic matter | 全N Total N | 容重 Bulk density | pH pH | 电导率 Conductivity | 土壤有机碳 Soil organic carbon | 地上生物量碳 Aboveground biomass carbon |
| 有机质Organic matter | 0.571** | |||||||
| 全N Total N | 0.462* | 0.842** | ||||||
| 容重Bulk density | -0.245 | -0.424* | -0.591** | |||||
| pH pH | 0.345 | 0.051 | 0.007 | 0.413* | ||||
| 电导率Conductivity | 0.359 | 0.582** | 0.836** | -0.644** | -0.012 | |||
| 土壤有机碳 Soil organic carbon | 0.598** | 0.835** | 0.694** | -0.139 | 0.314 | 0.536** | ||
| 地上生物量碳 Aboveground biomass carbon | 0.970* | 0.974* | 0.741 | -0.406 | -0.735 | 0.501 | 0.906* | |
| 地下生物量碳 Underground biomass carbon | 0.944* | 0.838 | 0.861 | -0.406 | -0.589 | 0.590 | 0.777 | 0.918* |
| *P<0.05, **P<0.01 | ||||||||
Cs=3.433-10.943X1+ 0.378X2-2.935X3+0.017X4
式中, Cs为土壤有机碳;X1为土壤含水量;X2为土壤有机质;X3为土壤全N;X4为土壤电导率。
生物量碳与土壤含水量、有机质呈显著正相关(P<0.05), 多元线性回归方程为:
Cp=2.042-11.930X1+0.011X2
式中, Cp为生物量有机碳;X1为土壤含水量;X2为土壤有机质。
说明在干旱环境下, 植物生物量对土壤含水量和土壤中有机质依赖较高, 而土壤有机碳主要来自土壤有机质, 并且土壤有机碳的积累与土壤中的含水量、含盐量以及全N的分解有一定的关系。
根据相关分析, 4种柽柳灌丛土壤有机碳密度均表现出与土壤有机质关系密切, 而除了多枝柽柳, 其余柽柳灌丛土壤有机碳与土壤全N含量密切相关, 多花柽柳和刚毛柽柳土壤有机碳与含水量关系密切;生物量碳密度均与土壤含水量、有机质表现出显著性相关关系(P<0.05), 多枝柽柳与刚毛柽柳地下生物量碳密度与土壤含水量呈现极显著相关关系(P<0.01), 说明二者的根系生长对水分要求更高。
3 讨论绿色植物通过光合作用吸收大气中的CO2固定为植物本身的有机碳, 在稳定全球气候、减缓温室效应方面发挥了重要作用。植物碳含量是植物碳贮量的一种度量, 反映绿色植物的固碳能力[26], 不同植物固碳能力有差异, 不同研究得出的结果也不同[27]。灌木生物量碳是植被碳库的度量, 是评价荒漠生态系统功能和进行碳循环研究的基础, 对研究生态系统的营养物质分配和碳循环具有重要意义[21]。柽柳是荒漠主要建群种之一, 对荒漠生态系统稳定具有重要作用, 其生物量研究一直受到学者的重视。安尼瓦尔等[14]、董道瑞等[15]、彭守璋等[16]都对柽柳属植物地上生物量进行过研究, 海依沙尔·哈力木江等[17]对柽柳地下生物量进行了研究, 而李丕军等[28]对3种柽柳含碳量进行了研究, 平均结果为48.67%。本研究对地上生物量和地下0—100 cm生物量进行了取样, 并根据碳含量系数(0.5) 进行了生物量碳分析, 结果显示4种柽柳灌丛地上生物量碳密度和地下生物量碳密度相差不是很大, 生物量碳密度在0.59—2.35 Mg/hm2之间, 根冠比在0.92—1.71之间, 但干旱区柽柳根系分布较深, 本研究取样深度不够导致根冠比要比实际小。生物量碳与土壤含水量、土壤有机质关系密切, 在干旱区, 土壤水分是植物生长最大的限制因子, 也是影响生物量有机碳库的重要因子, 单立山等[21], 王会提等[22]、张道远等[23]、都证明了水分对柽柳的生长和分布具有显著的影响。类似生境下, 4种柽柳表现出不同的适应性, 刚毛柽柳生长得更好, 表现为更大的生物量碳库, 沙生柽柳则主要生长于流动沙丘。因此, 在防风固沙的物种选择上也有一定的参考作用。
土壤有机碳对于生态系统过程、大气组成及气候变化速率的作用及其重要意义已得到了普遍的认同[10]。其积累是长期过程的结果, 本研究结果表明, 土壤有机碳与含水量、有机质、电导率有极显著正相关关系。土壤有机质是有机碳的主要来源, 土壤有机质来自地上植被的枯落物、植物根系、动物和微生物遗体等。其输入量在很大程度上取决于气候条件、土壤水分状况与养分有效性、植被生长及人为扰动等因素, 而有机物质的分解速率则依赖于有机物质的化学组成、土壤水热状况及物理化学等因[29]。土壤水分通过对土壤孔隙的通透性、植物根系和微生物产生影响, 从而影响土壤有机碳的积累[30]。根系对离子的选择吸收和吸收速率不同造成根际土壤盐离子变化[31], 柽柳通过泌盐作用富集盐离子, 进而影响土壤根际有机碳的积累。
灌丛“肥岛”有利于土壤有机碳的积累, 但“肥岛”现象随深度而逐渐变弱, 据研究, 柽柳灌丛“肥岛”的深度可达50 cm[32], 李龙[33]等按0—20, 20—60, 60—100 cm划分了3个层级分层取样研究, 结果显示土壤有机碳含量呈现先增加后降低的趋势, 与本文多枝与刚毛柽柳土壤有机碳含量变化一致;罗永清等[34]在科尔沁沙地不同坡位半固定沙丘的不同坡位土壤碳含量均随深度增加显著降低, 主要变异层发生在0—40 cm, 与本文多花与沙生柽柳土壤有机碳含量变化一致, 且多花与多枝柽柳土壤有机碳在40—50 cm深度活跃, 说明该层根系分布较多, 其死亡分解、分泌物以及根系与环境的共同作用, 如水分再分配、水力提升等导致该层较为活跃[30]。
4 结论新疆干旱区4种柽柳灌丛有机碳密度和碳贮量因其生物量差异以及与环境因子的适应性不同表现出了显著的差异性:有机碳密度从大到小的顺序是多花((65.60±7.83) Mg C/hm2)>刚毛((63.16±5.94) Mg C/hm2)>沙生((29.65±2.28) Mg C/hm2)>多枝((15.59±1.48) Mg C/hm2)。4种柽柳灌丛总有机碳贮量为75.00±6.76Tg, 以刚毛柽柳最高, 多花柽柳最低, 各柽柳灌丛有机碳贮量差异显著;土壤含水量、有机质对生物量碳和土壤有机碳有显著影响, 说明在干旱环境下, 柽柳的生长对土壤含水量和土壤中有机质依赖较高, 但不同类型的柽柳对水分和有机质的要求则有所不同。总之, 柽柳灌丛的碳汇作用显著, 在荒漠与绿洲过渡带生态系统中发挥着重要的作用, 但荒漠灌丛的类型较多, 碳汇能力和对荒漠生态系统的作用也不同。
| [1] | Schlesinger W H. Biogeochemistry, An Analysis of Global Change.2nd ed. San Diego, CA: Academic Pres, 1997. |
| [2] | Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 2000, 10(2): 423–436. DOI:10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2 |
| [3] | 胡会峰, 王志恒, 刘国华, 傅伯杰. 中国主要灌丛植被碳储量. 植物生态学报, 2006, 30(4): 539–544. |
| [4] | 张宏, 史培军, 郑秋红. 半干旱地区天然草地灌丛化与土壤异质性关系研究进展. 植物生态学报, 2001, 25(3): 366–370. |
| [5] | 郑敬刚, 张本昀, 何明珠, 保平, 张宝华, 李新荣. 灌丛化对贺兰山西坡草场土壤异质性的影响. 干旱区研究, 2009, 26(1): 26–31. |
| [6] | Connin S L, Virginia R A, Chamberlain C P. Carbon isotopes reveal soil organic matter dynamics following arid land shrub expansion. Oecologia, 1997, 110(3): 374–386. DOI:10.1007/s004420050172 |
| [7] | Goodale C L, Davidson E A. Carbon cycle:uncertain sinks in the shrubs. Nature, 2002, 418(6898): 593–594. DOI:10.1038/418593a |
| [8] | 姜广争, 宫渊波, 陈林武, 左琴, 刘金鑫, 岳艳杰, 张兴华. 岷江上游干旱河谷区灌丛植被土壤有机碳稳定性特征. 水土保持学报, 2011, 25(5): 209–214. |
| [9] | 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 2000, 24(5): 518–522. |
| [10] | 王百群, 吴金水, 吴振海. 子午岭次生林区植被中有机碳的储量. 西北植物学报, 2004, 24(10): 1870–1876. DOI:10.3321/j.issn:1000-4025.2004.10.018 |
| [11] | Ni J. Forage yield-based carbon storage in grasslands of China. Climatic Change, 2004, 67(2/3): 237–246. |
| [12] | 安尼瓦尔·买买提, 杨元合, 郭兆迪, 方精云. 新疆天山中段巴音布鲁克高山草地碳含量及其垂直分布. 植物生态学报, 2006, 30(4): 545–552. |
| [13] | 郭然, 王效科, 逯非, 段晓男, 欧阳志云. 中国草地土壤生态系统固碳现状和潜力. 生态学报, 2008, 28(2): 862–867. |
| [14] | 安尼瓦尔, 尹林克. 柽柳属植物的生物量研究. 新疆环境保护, 1997, 19(1): 46–50. |
| [15] | 董道瑞, 李霞, 万红梅, 林海军. 塔里木河下游柽柳灌丛地上生物量估测. 西北植物学报, 2012, 32(2): 384–390. |
| [16] | 彭守璋, 赵传燕, 彭焕华, 郑祥霖, 许仲林. 黑河下游柽柳种群地上生物量及耗水量的空间分布. 应用生态学报, 2010, 21(8): 1940–1946. |
| [17] | 海依沙尔·哈力木江, 努尔巴依·阿布都沙力克, 王山. 塔里木河中游地区柽柳地下生物量研究. 安徽农业科学, 2013, 41(12): 5539–5540, 5543-5543. DOI:10.3969/j.issn.0517-6611.2013.12.129 |
| [18] | 马虹, 陈亚宁, 李卫红. 荒漠河岸柽柳(Tamarix chinensis)灌丛的能量平衡特征. 中国沙漠, 2014, 34(1): 108–117. DOI:10.7522/j.issn.1000-694X.2013.00134 |
| [19] | 高培, 李从娟, 唐俊妍. 塔克拉玛干沙漠公路人工防护林植被的C、N化学计量特征及其固存能力研究. 沙漠与绿洲气象, 2015, 9(2): 63–68. |
| [20] | 许文强, 罗格平, 陈曦. 干旱区绿洲——荒漠过渡带灌丛土壤属性研究. 应用生态学报, 2006, 17(4): 583–586. |
| [21] | 单立山, 张希明, 王有科, 王蕙, 闫海龙, 魏疆, 许浩. 水分条件对塔里木沙漠公路防护林植物幼苗生长及生物量分配的影响. 科学通报, 2008, 53(S2): 82–88. |
| [22] | 王会提, 曾凡江, 张波, 刘波, 李尝君, 高欢欢, 郭自春. 不同灌溉量对绿洲——荒漠过渡带多枝柽柳幼苗根系生长和分布的影响. 西北植物学报, 2013, 33(12): 2521–2528. |
| [23] | 张道远, 尹林克, 潘伯荣. 柽柳属植物抗旱性能研究及其应用潜力评价. 中国沙漠, 2003, 23(3): 252–256. |
| [24] | Shi L, Zhao S Q, Tang Z Y, Fang J Y. The changes in China's forests:an analysis using the forest identity. PLoS One, 2011, 6(6): e20778. DOI:10.1371/journal.pone.0020778 |
| [25] | Gong Y M, Hu Y K, Fang F, Liu Y Y, Li K H, Zhang G M. Carbon storage and vertical distribution in three shrubland communities in Gurbantünggüt Desert, Uygur Autonomous Region of Xinjiang, Northwest China. Chinese Geographical Science, 2012, 22(5): 541–549. DOI:10.1007/s11769-012-0561-x |
| [26] | 鲍雅静, 李政海, 韩兴国, 宋国宝, 杨晓慧, 吕海燕. 植物热值及其生物生态学属性. 生态学杂志, 2006, 25(9): 1095–1103. |
| [27] | 郑帷婕, 包维楷, 辜彬, 何晓, 冷俐. 陆生高等植物碳含量及其特点. 生态学杂志, 2007, 26(3): 307–313. |
| [28] | 李丕军, 刘华, 王文月, 贾瑞琪, 董玲. 干旱区灌木树种柽柳含碳量分析. 南方农业学报, 2013, 44(11): 1874–1877. DOI:10.3969/j:issn.2095-1191.2013.11.1874 |
| [29] | 武小钢, 郭晋平, 杨秀云, 田旭平. 芦芽山典型植被土壤有机碳剖面分布特征及碳储量. 生态学报, 2011, 31(11): 3009–3019. |
| [30] | 弋良朋, 马健, 李彦. 荒漠盐生植物根际土壤盐分和养分特征. 生态学报, 2007, 27(9): 3565–3571. |
| [31] | 陈全胜, 李凌浩, 韩兴国, 阎志丹. 水分对土壤呼吸的影响及机理. 生态学报, 2003, 23(5): 972–978. |
| [32] | 李君, 赵成义, 朱宏, 王锋. 柽柳(Tamarix spp.)和梭梭(Haloxylon ammodendron)的"肥岛"效应. 生态学报, 2007, 27(12): 5138–5147. DOI:10.3321/j.issn:1000-0933.2007.12.023 |
| [33] | 李龙, 姚云峰, 秦富仓, 郭月峰, HaburaB, 常伟东. 半干旱区不同土层深度土壤有机碳变化. 水土保持通报, 2014, 34(4): 118–122. |
| [34] | 罗永清, 丁杰萍, 赵学勇, 李玉强, 周欣, 朱阳春, 岳祥飞, 张腊梅. 科尔沁沙地半固定沙丘不同坡位土壤C、N特征. 水土保持通报, 2015, 35(2): 94–100. |