 2017, Vol. 37
2017, Vol. 37文章信息
- 赵丽娅, 高丹丹, 熊炳桥, 陈涛, 李艳蔷, 李锋瑞
- ZHAO Liya, GAO Dandan, XIONG Bingqiao, CHEN Tao, LI Yanqiang, LI Fengrui.
- 科尔沁沙地恢复演替进程中群落物种多样性与地上生物量的关系
- Relationship between the aboveground biomass and species diversity of sandy communities during the process of restoring succession in the Horqin Sandy Land, China
- 生态学报. 2017, 37(12): 4108-4117
- Acta Ecologica Sinica. 2017, 37(12): 4108-4117
- http://dx.doi.org/10.5846/stxb201606161165
-
文章历史
- 收稿日期: 2016-06-16
- 修订日期: 2016-11-10





2. 中国科学院西北生态环境资源研究院, 兰州 730000
2. Northwest Institution of Eco-Environment and Resource, Chinese academy of sciences, Lanzhou 730000, China
生物多样性是生态系统结构的重要度量, 生物多样性的变化可以通过改变群落组成和结构、关键种的缺失以及改变物种对环境资源的利用方式等来影响生态系统功能[1-3]。生产力作为主要的生态系统功能, 其与生物多样性关系的研究已经成为生态学研究的热点[4]。生态学家Naeem等在控制环境的生长室建立了14个人工生态系统, 结果发现在同样的生长条件下, 物种最丰富的系统消耗最多CO2和产生最大的生物量, 即多物种比少物种具有较高生产力[5-8]。生态学家Tilman和Downing在天然草原和丢荒地的试验也说明物种多样性对于维持生产力水平的作用明显[9-10]。然而Huston、Wardle和Grime等认为这可能是由于取样效应引起的, 即从物种库中选取的物种数越多, 生产力高的物种出现的几率就会相应的增加, 使系统生产力随之升高, 并不是物种多样性的增加造成的[11-12]。在自然条件下, 生物多样性和生产力的关系具有复杂性。由于研究尺度、研究对象、研究区域等差异, 导致生物多样性与生态系统功能之间的关系结果各异。但是生态学者一致认为生物多样性对于生态系统的功能及稳定性有着重要的影响[4, 13-15]。探讨物种多样性和生产力及其之间关系, 对于阐明生物多样性与生态系统功能的关系及其内在机制有着重要意义。
科尔沁沙地地处松嫩草原与内蒙古高原的过渡地带, 由于滥垦、滥牧、滥樵导致植被大量破坏, 土地风蚀沙化严重, 生态环境脆弱, 成为我国北方农牧交错地区典型的生态脆弱带。但与我国其他沙漠、沙地相比较, 降水量在350—450mm之间, 自然条件较为优越, 本底植被为疏林草原, 大部分沙地主要由草原开垦转化而来。如果停止开垦, 沙地便有逆转的可能[16-18]。因此, 如何恢复或整治环境, 重建沙地生态系统, 已成为人们广泛关注的焦点问题。故研究掌握科尔沁沙地植被恢复演替过程中群落物种多样性和生产力的变化, 对认识该区植被恢复的过程、促进退化生态系统结构和功能重建具有重要的指导意义。
1 材料与方法 1.1 研究区概况研究区位于科尔沁沙地中南部的奈曼旗中国科学院奈曼沙漠化研究站境内。地理位置120°41′E, 42°54′N。该区属半干旱气候, 年平均降水量366mm, 年蒸发量1935mm, 年均气温6.5℃, 1月平均气温-12.7℃, 7月平均气温23.7℃, ≥10℃积温3 000℃以上, 无霜期150d。土壤类型为沙质栗钙土, 经破坏后则退化为流动风沙土, 沙土基质分布广泛, 风沙活动强烈[16]。由于沙土的干燥疏松和干旱多风等气候条件的影响, 使得该区在过度放牧和过垦等人为不合理利用的情况下, 植被遭受破坏, 土地沙化严重。但是由于沙土良好的水分储藏性能, 在大量天然种源存在的情况下和禁止放牧、禁止开垦等良好的保护措施下, 植被可以快速建立并向稳定植被类型演替的可能[17-18]。
1.2 野外调查方法以流动沙丘先锋植物——沙米群落的建立为植被恢复的起点, 沙地生态系统恢复的过程伴随着植被发育和群落演替, 也是松散沙粒基质的流动性不断降低、沙地逐渐得到固定的过程。由流动沙丘到半固定沙丘、再到固定沙丘, 生态系统恢复的每一阶段都发育着典型的群落类型, 沙地环境的变化与群落类型的演替变化相一致和相对应[19], 因此把沙地环境从流动沙丘到半固定沙丘、再到固定沙丘的几个明显的阶段作为生态系统恢复阶段。群落演替时间的确定则是从进行围封、完全停止干扰后开始计算。该文用时-空替代的方法, 在野外选取分别处于流动沙丘、半固定沙丘和固定沙丘等不同恢复阶段的6个演替时间互不相等的典型群落样地进行调查。所选的6个群落样地其围封历史清楚, 围封期间保护较好, 基本未受放牧和人为破坏等干扰。从进行围封停止干扰后开始算起, 6个群落的演替时间分别为1、3、5、12、15a和20a。从沙地环境状况判断其所处的恢复阶段, 1a和3a的群落处于流动沙丘阶段, 5a和12a的群落处于半固定沙丘阶段, 15a和20a的群落处于固定沙丘阶段。
流动沙丘恢复阶段:地表几乎被流沙覆盖, 无结皮或少量物理性结皮, 植被盖度小于20%。恢复1a的群落以沙米(Agriophyllum squarrosum)、差巴嘎蒿(Artemisia halodendron)为主, 恢复3a的群落以大果虫实(Corispermum macrocarpum)和狗尾草(Setaria viridis)为主。
半固定沙丘恢复阶段:流沙面积大于10%, 地表已有结皮, 但结皮数量较少少, 植被盖度在20%—60%。主要植物有大果虫实、狗尾草和三芒草(Aristida adscensionis)等。
固定沙丘恢复群落:流沙面积低于10%, 地表覆盖固定风沙土或沙质栗钙土或已形成较厚的土壤结皮, 植被盖度大于50%。主要植物种有画眉草(Eragrostis pilosa)、虎尾草(Chloris virgata)、达乌里胡枝子(Lespedeza davurica)、黄蒿(Artemisia scoparia)和糙隐子草(Cleistogenes squarrosa)等。
2014年8月初, 分别在6个群落样地的典型地段等距离(20m, 南北向)设置3条长200m的样带, 在每条样带上等距离(10m)设置20个固定样点, 每个样地60个样点, 共计360个样点, 在每个取样点的旁边, 设置面积1m2的草本样方, 测定每个样方的种类组成及其特征值(盖度、高度和密度), 采用齐地面刈割法调查地上生物量, 草样在85℃烘干24h称重。各样地每种植物的出现频率根据60个样点的测定数据计算。
1.3 数据处理 1.3.1 物种重要值和优势度IV=相对密度+相对频度+相对高度+相对盖度
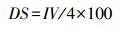
|
IV为物种重要值, DS为物种的优势度。
1.3.2 物种多样性物种多样性采用simpson指数、Shannon-Wiener多样性指数和Pielow均匀度指数来衡量[20]。
Simpson指数
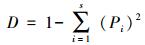
|
Shannon-Wiener多样性指数

|
Pielow均匀度指数:
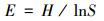
|
式中, S为物种总数, Pi为第i种植物的种数占群落中总种数的比例。鉴于群落中物种的其它信息, 如盖度、生物量、重要值等指标更能反映群落的本质特征, 上述数量数据中的个体数可以由这些指标来代替。本文用重要值代替个体数来计算。
1.3.3 统计分析采用回归分析研究群落生物多样性的地上生物量的关系, 采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较6个植物群落地上生物量之间的差异。均在SPSS18.0软件下完成。
2 结果与分析 2.1 群落基本特征表 1列出了植被恢复系列上群落种类组成及物种优势度。不难发现, 沙地群落恢复演替进程中, 物种丰富度逐渐增加, 从流动沙丘的6种到固定沙丘的30种, 增加了400%的比例。每一物种的优势度在群落中的表现不同, 优势物种也发生了变化。在恢复演替1 a的流动沙丘, 沙米(Agriophyllum squarrosum)和差巴嘎蒿(Artemisia halodendron)是群落的优势物种, 占92.56%的比例:大果虫实(Corispermum macrocarpum)、猪毛菜(Salsola collina Pall)、狗尾草(Setaria viridis)和差巴嘎蒿是演替3年流动沙丘的优势种, 占89.27%;演替5 a的半固定沙丘, 优势物种是大果虫实、狗尾草和差巴嘎蒿, 占67.52%;大果虫实、三芒草(Aristida adscensionis)、狗尾草、差巴嘎蒿和达乌里胡枝子(Lespedeza davurica)是演替12a半固定沙丘的优势物种, 占66.25%;画眉草(Eragrostis pilosa)、虎尾草(Chloris virgata)、太阳花(Erodium stephanianum)和达乌里胡枝子是演替15 a固定沙丘群落的优势物种, 占51.73%;在演替20a的固定沙丘, 画眉草、狗尾草、黄蒿(Artemisia scoparia)和糙隐子草(Cleistogenes squarrosa)是群落的优势物种, 占70.93%的比例。
| 植物种 Species |
恢复阶段Restoration stage | |||||||
| 流动沙丘 Shifting sandy dune |
半固定沙丘 Semi-fixed sandy dune |
固定沙丘 Fixed sandy dune |
||||||
| 1a | 3a | 5a | 12a | 15a | 20a | |||
| 大果虫实Corispermum macrocarpum | — | 56.00 | 21.50 | 12.78 | 3.04 | 0.91 | ||
| 猪毛菜Salsola collina | — | 6.67 | 5.94 | 4.19 | 7.00 | 0.98 | ||
| 沙米Agriophyllum squarrosum | 48.54 | 0.94 | 0.23 | — | — | — | ||
| 五星蒿Bassia dasyphylla | 2.33 | 3.88 | 1.36 | 4.23 | 0.64 | 0.11 | ||
| 灰绿藜Chenopodium glaucum | — | — | — | — | 0.10 | 1.83 | ||
| 地肤Kochia scoparia | — | — | — | — | 0.31 | 0.66 | ||
| 锋芒草Tragus mongolorum | — | — | — | — | — | 0.11 | ||
| 三芒草Aristida adscensionis | — | 1.43 | 5.59 | 13.20 | 3.46 | 4.24 | ||
| 狗尾草Setaria viridis | 4.35 | 19.84 | 17.40 | 21.42 | 6.49 | 15.06 | ||
| 画眉草Eragrostis pilosa | — | — | 0.99 | 4.20 | 13.34 | 17.60 | ||
| 虎尾草Chloris virgata | — | — | — | 0.42 | 15.45 | 2.06 | ||
| 毛马唐Digitaria cilliaris | — | — | 2.10 | 3.31 | 1.28 | 0.0005 | ||
| 野黍Eriochloa villosa | — | — | — | — | 0.05 | 6.01 | ||
| 细叶苦荬菜Ixeris denticulata | — | — | 0.86 | 1.22 | — | — | ||
| 大籽蒿Artemisia sieversiana | — | — | — | — | — | 0.96 | ||
| 黄蒿Artemisia scoparia | 0.38 | — | — | — | 0.33 | 19.69 | ||
| 鸡眼草Kummerowia striata | — | — | — | — | 0.29 | 0.39 | ||
| 褐虱Lappula myosotis | — | 0.66 | 4.32 | 6.34 | 2.12 | 0.15 | ||
| 地锦Euphorbia humifusa | — | 3.84 | — | — | 0.02 | 0.09 | ||
| 蒺藜Tribulus terrestris | — | — | — | — | 0.07 | — | ||
| 太阳花Erodium stephanianum | — | — | — | — | 9.14 | — | ||
| 马齿苋Portulaca oleracea | — | — | — | — | 4.34 | 0.21 | ||
| 糙隐子草Cleistogenes squarrosa | — | — | — | — | 5.63 | 18.57 | ||
| 白草Pennisetum centrasiaticum | — | — | — | 0.01 | — | 4.58 | ||
| 赖草Leymus secalinus | — | — | 0.67 | — | 2.51 | 0.21 | ||
| 芦苇Phragmites communis | — | — | 0.15 | — | 0.14 | |||
| 扁蓿豆Melissitus ruthenicus | — | — | 2.33 | 3.91 | 3.70 | 0.80 | ||
| 狭叶米口袋Gueldenstaedtia stenophylla | — | — | — | — | — | 0.13 | ||
| 白山蓟Olgaea leucophylla | — | — | 1.31 | 0.38 | — | 0.58 | ||
| 苔草Carex dispalata | — | — | — | — | — | 0.08 | ||
| 地稍瓜Cynanchum theisiodes | 0.39 | — | 3.42 | 0.25 | 0.37 | 0.76 | ||
| 乳桨大戟Euphorbia esula | — | — | 0.38 | — | — | — | ||
| 柳穿鱼Linaria vulgaris | — | — | 0.70 | 3.45 | — | — | ||
| 防风草Saposhnikovis divaricata | — | — | — | 0.57 | 3.42 | 0.30 | ||
| 沙葱Allium mongolicum | — | — | — | — | — | 0.95 | ||
| 差巴嘎蒿Artemisia halodendrom | 44.02 | 6.76 | 28.62 | 12.23 | 0.06 | — | ||
| 达乌里胡枝子Lespedeza davurica | — | — | 2.12 | 6.62 | 13.80 | 1.88 | ||
| 山竹岩黄芪Hedysarum fruticosum | — | — | — | 1.29 | 3.04 | — | ||
| 总种数Sum of species | 6 | 9 | 19 | 19 | 26 | 30 | ||
从生活型结构来看(表 2), 一年生草本植物在各群落中占绝对优势, 均在50%以上。但随着演替的进行, 生活型结构呈现多样化, 多年生植物种类明显增加, 固定沙丘(演替20a)阶段, 多年生草本植物达到12种, 物种生活型组成上的这种变化反映了恢复演替过程中植物群落与环境之间的变化特征。
| 演替时间/a Succession time |
一年生草本Annuals | 二年生草本Biennials | 多年生草本Perennials | 灌木类Shrubs | |||||||
| 种数Ns | 优势度DS | 种数Ns | 优势度DS | 种数Ns | 优势度DS | 种数Ns | 优势度DS | ||||
| 1 | 4 | 55.59 | — | — | 1 | 0.39 | 1 | 44.02 | |||
| 3 | 8 | 93.24 | — | — | — | — | 1 | 6.76 | |||
| 5 | 9 | 59.44 | 1 | 0.86 | 7 | 8.96 | 2 | 30.75 | |||
| 12 | 9 | 70.08 | 1 | 1.22 | 6 | 8.57 | 3 | 20.13 | |||
| 15 | 15 | 53.67 | 2 | 4.67 | 6 | 24.77 | 3 | 16.90 | |||
| 20 | 15 | 50.18 | 2 | 19.9 | 12 | 28.04 | 1 | 1.88 | |||
| Ns: Number of species; Ds: Dominance of species | |||||||||||
表 3给出植被恢复系列上群落种类组成的科属结构。伴随着演替的发生, 科属种类增加, 多样化特征明显。藜科植物在流动沙丘阶段优势度明显, 在50%以上, 到半流动沙丘阶段, 藜科植物的优势度下降到30%以下, 固定沙丘阶段, 藜科植物的优势度下降到11%以下。藜科植物之所以在流动沙丘占优势, 是因为沙米、大果虫实等藜科植物对严酷的沙地环境适应性强。随着演替的进行, 禾本科植物的种数和优势度均在增加, 种数由1种上升到11种, 优势度从4.35增加到68.65, 禾本科植物在群落中的地位逐渐增强。菊科植物差巴嘎蒿在流动沙丘阶段的优势度明显, 随着演替的进行, 逐渐退出固定沙丘群落。在固定沙丘阶段(20a), 菊科植物的优势度为21.33, 贡献大的物种主要是黄蒿(Artemisia scoparia)。恢复演替过程中, 豆科植物的种数增加, 但优势度波动变化。
| 演替时间/a Succession time |
总属数 Ng |
总科数 Nf |
藜科Chenopodiaceae | 菊科Compositae | 禾本科Gramineae | 豆科Leguminosae | |||||||||||
| 属数 Ng |
种数 Ns |
优势度 Ng |
属数 Ns |
种数 Ng |
优势度 Ns |
属数 Ng |
种数 Ns |
优势度 Ng |
属数 Ns |
种数 Ng |
优势度 Ns |
||||||
| 1 | 5 | 4 | 2 | 2 | 50.86 | 1 | 2 | 44.40 | 1 | 1 | 4.35 | — | — | — | |||
| 3 | 9 | 5 | 4 | 4 | 67.49 | 1 | 1 | 6.76 | 2 | 2 | 21.27 | — | — | — | |||
| 5 | 18 | 6 | 4 | 4 | 29.03 | 2 | 3 | 30.79 | 6 | 6 | 26.90 | 2 | 2 | 4.45 | |||
| 12 | 19 | 8 | 3 | 3 | 21.20 | 2 | 2 | 12.61 | 6 | 6 | 42.56 | 3 | 3 | 11.82 | |||
| 15 | 25 | 11 | 5 | 5 | 11.09 | 1 | 2 | 0.39 | 8 | 8 | 48.21 | 4 | 4 | 20.83 | |||
| 20 | 29 | 11 | 5 | 5 | 4.49 | 2 | 3 | 21.33 | 11 | 11 | 68.65 | 4 | 4 | 3.20 | |||
| Ng: Number of genera;Nf: Number of families;Ns: Number of species;Ds: Dominance of species | |||||||||||||||||
表 4从物种多样性指数和均匀度等方面反映群落物种组成的结构水平。结果发现, 群落物种多样性随着演替而增加, 演替到12a的群落时, 达到最大, 之后随着演替呈降低态势。流动沙丘是植被恢复演替的起点, 多样性测度值最小, 演替到12a的半流动沙丘, 多样性测度值达到最大, 之后在固定沙丘出现下降。科尔沁沙地植被恢复演替过程中, 演替机制决定于物种间的竞争能力, 沙米种群的退出, 差巴嘎蒿种群的消亡, 沙地环境发生了变化, 为其他禾本科和豆科植物的入侵创造了条件, 物种数不断增加, 生物多样性也维持在较高的水平[21]。
| 物种多样性 Species diversity |
恢复阶段Restoration stage | |||||||
| 流动沙丘 Shifting sandy dune |
半固定沙丘 Semi-fixed sandy dune |
固定沙丘 Fixed sandy dune |
||||||
| 1a | 3a | 5a | 12a | 15a | 20a | |||
| Simpson指数 Simpson index |
0.5682 | 0.6347 | 0.8298 | 0.8869 | 0.8788 | 0.8638 | ||
| Shannon-Wiener指数 Shannon-Wiener index |
0.4251 | 0.6066 | 0.9287 | 1.0484 | 1.0399 | 1.0114 | ||
| 均匀度指数 Evenness index |
0.5463 | 0.6356 | 0.7263 | 0.8198 | 0.7349 | 0.6847 | ||
沙地植被恢复演替系列上, 群落地上生物量呈增加趋势, 从流动沙丘到固定沙丘(3个恢复阶段的均值计算), 干物质量增加了580%(表 5)。方差分析结果表明:在沙地植被恢复系列上群落地上生物量存在显著的差异(F=18.926, n=360, P<0.001)。此外, 沙地植被恢复演替进程中, 不同植物种的地上生物量变化方式也不同。沙米、大果虫实的生物量逐渐减少, 差巴嘎蒿、达乌里胡枝子以及禾本科植物等呈现波动变化。
| 植物种 Species |
恢复阶段Restoring stages | F | P | |||||||
| 流动沙丘 Shifting sandy dune |
半固定沙丘 Semi-fixed sandy dune |
固定沙丘 Fixed sandy dune |
||||||||
| 1a | 3a | 5a | 12a | 15a | 20a | |||||
| 大果虫实Corispermum macrocarpum | — | 12.94 ± 3.85ab | 17.83 ± 3.92a | 13.40 ± 2.23ab | 6.18 ± 2.89b | 1.24 ± 0.71c | 8.7 | <0.001 | ||
| 猪毛菜Salsola collina | — | 2.49 ± 1.61a | 4.42 ± 2.20a | 4.97 ± 2.39a | 24.32 ± 7.52b | 1.67 ± 0.85a | 7.8 | <0.001 | ||
| 沙米Agriophyllum squarrosum | 1.63 ± 0.20a | 0.02 ± 0.01b | 0.01 ± 0.01b | — | — | — | 43.1 | <0.001 | ||
| 五星蒿Bassia dasyphylla | 0.03 ± 0.01a | 1.15 ± 0.98a | 0.20 ± 0.07a | 4.73 ± 2.91b | 0.38 ± 0.20a | 0.01 ± 0.01a | 2.5 | <0.05 | ||
| 灰绿藜Chenopodium glaucum | — | — | — | — | 0.02 ± 0.01a | 1.27 ± 0.61b | 4.7 | <0.001 | ||
| 地肤Kochia scoparia | — | — | — | — | 0.02 ± 0.01a | 0.07 ± 0.02b | 6.8 | <0.001 | ||
| 锋芒草Tragus mongolorum | — | — | — | — | — | 0.02 ± 0.02 | — | — | ||
| 三芒草Aristida adscensionis | 0.01 ± 0.01a | 0.68 ± 0.35a | 4.58 ± 1.22b | 1.77 ± 0.88ac | 2.59 ± 0.71c | 7.8 | <0.001 | |||
| 狗尾草Setaria viridis | 0.10 ± 0.04a | 4.40 ± 2.11ac | 11.55 ± 2.14b | 18.36 ± 3.63d | 9.86 ± 1.49bc | 20.52 ± 3.08d | 14.1 | <0.001 | ||
| 画眉草Eragrostis pilosa | — | — | — | 1.82 ± 1.03a | 8.76 ± 1.37b | 23.54 ± 4.51c | 25.4 | <0.001 | ||
| 虎尾草Chloris virgata | — | — | — | — | 20.01 ± 4.84a | 3.14 ± 2.70b | 13.8 | <0.001 | ||
| 毛马唐Digitaria cilliaris | — | — | 0.66 ± 0.23a | 2.60 ± 2.14a | 0.40 ± 0.15a | 6.48 ± 1.32b | 7.1 | <0.001 | ||
| 细叶苦荬菜Ixeris denticulata | — | — | 0.74 ± 0.50a | 0.96 ± 0.52a | — | — | 不显著 | |||
| 大籽蒿Artemisia sieversiana | — | — | — | — | — | 1.38 ± 0.95 | — | — | ||
| 黄蒿Artemisia scoparia | 0.001 ± 0.001a | — | — | — | 0.14 ± 0.05a | 19.93 ± 3.64b | 2.3 | <0.05 | ||
| 鸡眼草Kummerowia striata | — | — | — | — | — | 0.07 ± 0.05 | 32.8 | <0.001 | ||
| 地锦Euphorbia humifusa | — | 0.03 ± 0.02a | 0.84 ± 0.23a | 1.61 ± 0.20a | 2.07 ± 0.71a | 0.07± 0.04a | 不显著 | |||
| 蒺藜Tribulus terrestris | — | 1.91 ± 1.75a | — | — | 0.01 ± 0.01a | 0.09± 0.08a | 不显著 | |||
| 太阳花Erodium stephanianum | — | — | — | — | 0.01 ± 0.01a | 0.18± 0.10b | 3.6 | <0.01 | ||
| 马齿苋Portulaca oleracea | — | — | — | — | 0.01 ± 0.01 | — | — | — | ||
| 糙隐子草Cleistogenes squarrosa | — | — | — | — | 38.65 ± 8.69a | 42.63± 3.64b | 33.4 | <0.01 | ||
| 白草Pennisetum centrasiaticum | — | — | — | — | — | 5.55± 1.55 | — | — | ||
| 赖草Leymus secalinus | — | — | 0.17 ± 0.14a | — | 0.01 ± 0.01a | 0.20± 0.20a | 不显著 | |||
| 芦苇Phragmites communis | — | — | 0.06 ± 0.06a | — | — | 0.11± 0.11a | 不显著 | |||
| 扁蓿豆Melissitus ruthenicus | — | — | 1.94 ± 1.61a | 4.91 ± 2.87a | 2.66 ± 2.27a | 0.65± 0.35a | 不显著 | |||
| 白山蓟Olgaea leucophylla | — | — | 1.63 ± 0.37a | 0.26 ± 0.14a | — | 1.56± 1.18a | 不显著 | |||
| 苔草Carex dispalata | — | — | — | — | — | 0.02± 0.02 | — | — | ||
| 地稍瓜Cynanchum theisiodes | 0.01 ± 0.01a | — | 1.24 ± 0.28b | 0.08 ± 0.04ac | 0.04 ± 0.03a | 0.67± 0.45bc | 6.1 | <0.001 | ||
| 乳桨大戟Euphorbia esula | — | — | 0.31 ± 0.31 | — | — | — | — | — | ||
| 柳穿鱼Linaria vulgaris | — | — | 0.38 ± 0.27a | 2.86 ± 1.33b | — | — | 4.9 | <0.001 | ||
| 防风草Saposhnikovis divaricate | — | — | — | 0.47 ± 0.47a | 0.23 ± 0.12a | 0.09± 0.06a | 不显著 | |||
| 沙葱Allium mongolicum | — | — | — | — | — | 0.28± 0.12 | — | — | ||
| 差巴嘎蒿Artemisia halodendrom | 28.25 ± 5.55a | 0.76 ± 0.40b | 1.86 ± 1.43b | 14.70 ± 4.77c | 0.02 ± 0.02b | — | 11.8 | <0.001 | ||
| 达乌里胡枝子Lespedeza davurica | — | — | 46.04 ± 11.53b | 5.65 ± 2.55a | 81.56 ± 11.73c | 3.84± 1.85a | 27.4 | <0.001 | ||
| 山竹岩黄芪Hedysarum fruticosum | — | — | — | 1.48 ± 0.98a | 28.91 ± 11.45b | — | 6.8 | <0.001 | ||
| 小计Totals | 29.79 ± 5.82ab | 23.70 ± 10.74b | 90.56 ± 25.64ac | 83.43 ± 29.43ac | 226.04 ± 54.47d | 137.87± 28.92e | 18.93 | <0.001 | ||
回归分析表明, 沙地植被恢复演替系列上, 群落物种多样性和地上生物量之间均存在显著的相关关系, 拟合曲线为负二次函数关系, 即单峰型函数关系(图 1)。物种多样性与生物量之间的单峰函数关系可能是异质性生境中生态位特化选择的结果[22-23]。物种数、Simpson指数、Shannon-Wiener指数和均匀度指数与生物量的拟合系数分别为0.9354、0.873、0.8448和0.4736(P<0.001)。可以得出沙地植被恢复过程中相对高生产力的群落具有高物种多样性的结论。固定沙丘群落具有高的生物量和高物种多样性倾向, 而流动沙丘群落具有低的生物量和低物种多样性的倾向。

|
| 图 1 植被恢复演替进程中群落物种多样性与地上生物量的关系 Fig. 1 Relationship between species diversity and aboveground biomass during the process of restoring succession |
科尔沁沙地植被恢复过程中, 每一群落对环境的改变都对下一阶段群落的形成和物种入侵创造了条件。群落环境实现了从重度沙漠化到潜在沙漠化的变化, 植物种数由流动沙丘的6种增加到固定沙丘的30种, 藜科植物在流动沙丘阶段优势度明显, 之后逐渐下降;禾本科植物的种数和优势度逐渐增加, 意味着禾本科植物在群落中的地位逐渐增强。群落地上生物量也呈增加趋势, 从流动沙丘到固定沙丘(3个恢复阶段的均值计算)地上生物量增加了580%, 这为自然状态下的植被恢复和生物多样性保护提供了佐证。土地沙漠化实质就是人类不合理的利用造成的土地退化和生态系统破坏, 因此首先要改变不合理的利用方式, 使沙米等先锋植物定居下来, 植被恢复得以开始, 来保护沙地植被[18-19, 21]。
物种多样性与生产力关系的单峰格局目前主要有3种解释:一种认为高生产力导致高速的种群增长, 使潜在的竞争排除得以迅速实现;一种认为中等干扰是关键因素, 单峰关系并非生产力本身造成, 可能是一些其他干扰因子和生产力平行变化;另一种解释认为高生产力环境中资源空间异质性下降, 从而导致竞争排除增加[22-23]。Guo和Berry在Chilhuuan沙漠研究认为, 影响物种丰富度与生产力关系的因素除空间尺度外, 还有被测群落环境梯度变化的幅度, 即环境梯度从贫瘠到丰富的生境变化时, 物种丰富度与生产力的关系为单峰形式, 环境条件范围越大, 单峰形式越明显[24]。从研究结果看, 沙地植被恢复演替进程中, 群落的物种多样性与地上生物量呈负二次函数关系, 支持随着群落生产力的增加, 中等生产力水平物种多样性高的理论[25]。科尔沁沙地群落物种多样性的变化, 即严重沙漠化类型具有低的生产力和低的物种多样性倾向, 而轻度和中度沙漠化类型具有中等生产力水平和最高的物种多样性。中等生产力水平物种多样性最高, 生产力与多样性之间的这种关系可能是异质性生境中生态位特化选择的结果。尤其是水分条件所引起的环境变化、物种更迭和竞争等是形成单峰关系的主要原因[4, 26-30]。
生物多样性的变化以及多样性和生产力关系受多种因素影响。相关研究表明, 除了人类干扰外, 多样性和生产力的关系会随着环境因子以及不同管理方式的改变而变化[31]。在自然环境中, 不同的影响因素是共存的, 需要找到在影响多样性和生产力关系中起到关键作用的影响因素, 这样才能就生物多样性与生产力关系的机制进行准确而深入的探讨。在20a的时间里, 沙地环境实现了从极端退化的流动沙地到固定沙地的转化, 群落环境、物种多样性和生产力均发生了显著的变化。对于这种由于人类不合理利用造成的生态系统退化, 重要的就是停止不合理的土地利用方式, 植被休养生息得以恢复, 获得较高的草地生物多样性和生产力。
群落物种多样性与生产力关系的研究是阐明物种多样性对生物群落功能作用的重要途径之一。物种多样性的变化与群落的环境密切相关。物种多样性不仅能够表征群落的组成结构, 而且也能对环境状况做出指示[32]。群落生产力是衡量群落功能最重要的指标, 其变化能够反映群落结构及其功能的变化。在科尔沁沙地恢复演替进程中, 沙质草地环境的改变对某一阶段群落的形成和物种侵入创造了条件, 一定的群落环境对应着一定的物种多样性水平和群落的生产力, 严酷环境条件下总是能够支持较少的物种, 而环境条件的改善则允许更多物种的共存。可以说, 一定条件下的物种多样性和群落生产力与其所处的环境状况和环境容量紧密相关。因此, 除了考虑不合理的人为干扰因素外, 更重要的是要考虑科尔沁沙地的环境容纳量, 以及其供给水平, 这才是保护沙地生物多样性和提高群落生产力的正道, 也是多样性与生产力关系研究中值得深入探讨的问题。
4 结论(1) 科尔沁沙地恢复演替过程中, 物种丰富度逐渐增加, 从流动沙丘的6种到固定沙丘的30种, 增加了400%的比例。一年生草本植物在各群落中都占绝对优势, 但随着演替的进行, 生活型结构呈现多样化, 多年生植物种类明显增加。藜科植物在流动沙丘阶段优势度明显, 之后逐渐下降;禾本科植物的种数和优势度逐渐增加, 意味着禾本科植物在群落中的地位逐渐增强。群落地上生物量也呈增加趋势, 从流动沙丘到固定沙丘(3个恢复阶段的均值计算)地上生物量增加了580%。
(2) 回归分析表明, 沙地植被恢复演替系列上, 群落生物多样性和地上生物量之间均存在显著的相关关系, 拟合曲线为负二次函数关系, 即单峰型函数关系(P<0.001)。表明沙地植被恢复过程中, 相对高生产力的群落具有高物种多样性, 固定沙丘群落具有高的生物量和高物种多样性的倾向, 而流动沙丘群落具有低的生物量和低物种多样性的倾向。
| [1] | Huston M A. Hidden treatments in ecological experiments:re-evaluating the ecosystem function of biodiversity. Oecologia, 1997, 110(4): 449–460. DOI:10.1007/s004420050180 |
| [2] | Tilman D. Causes, consequences and ethics of biodiversity. Nature, 2000, 405(6783): 208–211. DOI:10.1038/35012217 |
| [3] | Hector A, Bazeley-White E, Loreau M, Otway S, Schmid B. Overyielding in grassland communities:testing the sampling effect hypothesis with replicated biodiversity experiments. Ecology Letters, 2002, 5(4): 502–511. DOI:10.1046/j.1461-0248.2002.00337.x |
| [4] | 彭少麟, 黄忠良. 生产力与生物多样性之间的相互关系研究概述. 生态科学, 2000, 19(1): 1–9. |
| [5] | Naeem S, Thompson L J, Lawler S P, Lawton J H, Woodfin R M. Declining biodiversity can alter the performance of ecosystems. Nature, 1994, 368(6473): 734–737. DOI:10.1038/368734a0 |
| [6] | Naeem S, Thompson L J, Lawler S P, Lawton J H, Woodfin R M. Empirical evidence that declining species diversity may alter the performance of terrestrial ecosystems. Philosophical Transactions of the Royal Society B, 1995, 347(1321): 249–262. DOI:10.1098/rstb.1995.0025 |
| [7] | Naeem S, Håkansson K, Lawton J H, Crawley M J, Thompson L J. Biodiversity and plant productivity in a model assemblage of plant species. Oikos, 1996, 76(2): 259–264. DOI:10.2307/3546198 |
| [8] | Wardle D A. Reply:A more reliable design for biodiversity study?. Nature, 1998, 394(6688): 30–30. |
| [9] | Tilman D, Downing J A. Biodiversity and stability in grasslands. Nature, 1994, 367(6461): 363–365. DOI:10.1038/367363a0 |
| [10] | Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 1996, 379(6567): 718–720. DOI:10.1038/379718a0 |
| [11] | Huston M A, Aarssen L W, Austin M P, cade B S, Fridley J D, Garnier E, Grime J P, Hodgson J, Lauenroth W K, Thompson K, Vandermeer J H, Wardle D A, Hector A, Schmid B, Beierkuhnlein C, Caldeira M C, Diemer M, Dimitrakopoulos P G, Finn J A, Freitas H, Giller P S, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen A, Körner C, Leadley P W, Loreau M, Minns A, Mulder C P H, O'donovan G, Otway S J, Pereira J S, Prinz A, Read D J, Scherer-Lorenzen M, Schulze E D, Siamantziouras A S D, Spehn E, Terry A C, Troumbis A Y, Woodward F I, Yachi S, Lawton J H. No consistent effect of plant diversity on productivity. Science, 2000, 289(5483): 1255–1255. DOI:10.1126/science.289.5483.1255a |
| [12] | Wardle D A, Huston M A, Grime J P, Berendse F, Garnier E, Lauenroth W K, Setälä H, Wilson S D. Biodiversity and ecosystem functioning:an issue in ecology. Bulletin of the Ecological Society of America, 2000, 81(3): 235–239. |
| [13] | Nguyen H, Herbohn J, Firn J, Lamb D. Biodiversityproductivity relationships in small-scale mixed-species plantations using native species in Leyte province, Philippines. Forest Ecology and Management, 2012, 274: 81–90. DOI:10.1016/j.foreco.2012.02.022 |
| [14] | Shanafelt D W, Dieckmann U, Jonas M, Franklin O, Loreau M, Perrings C. Biodiversity, productivity, and the spatial insurance hypothesis revisited. Journal of Theoretical Biology, 2015, 380: 426–435. DOI:10.1016/j.jtbi.2015.06.017 |
| [15] | Carnicer J, Sardans J, Stefanescu C, Ubach A, Bartrons M, Asensio D, Peñuelas J. Global biodiversity, stoichiometry and ecosystem function responses to human-induced C-N-P imbalances. Journal of Plant Physiology, 2015, 172: 82–91. DOI:10.1016/j.jplph.2014.07.022 |
| [16] | 刘新民, 赵哈林, 赵爱芬. 科尔沁沙地风沙环境与植被. 北京: 科学出版社, 1996. |
| [17] | 蒋德明, 刘志民, 曹成有, 寇振武, 王汝槦. 科尔沁沙地荒漠化过程与生态恢复. 北京: 中国环境科学出版社, 2003. |
| [18] | 赵哈林, 赵学勇, 张铜会, 周瑞莲. 沙漠化的生物过程及退化植被的恢复机理. 北京: 科学出版社, 2007. |
| [19] | 李胜功, 赵爱芬, 常学礼. 科尔沁沙地植被演替的几个问题. 中国沙漠, 1997, 17(S1): 25–33. |
| [20] | 马克平. 生物群落多样性的测度方法Ⅰ.α多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. DOI:10.17520/biods.1994027 |
| [21] | 吕朋, 左小安, 张婧, 周欣, 连杰, 刘良旭. 放牧强度对科尔沁沙地沙质草地植被的影响. 中国沙漠, 2016, 36(1): 34–39. DOI:10.7522/j.issn.1000-694X.2015.00197 |
| [22] | 郑晓翾, 王瑞东, 靳甜甜, 木丽芬, 刘国华. 呼伦贝尔草原不同草地利用方式下生物多样性与生物量的关系. 生态学报, 2008, 28(11): 5392–5400. DOI:10.3321/j.issn:1000-0933.2008.11.022 |
| [23] | 杨利民, 周广胜, 李建东. 松嫩平原草地群落物种多样性与生产力关系的研究. 植物生态学报, 2002, 26(5): 589–593. |
| [24] | Guo Q F, Berry W L. Species richness and biomass:dissection of the hump-shaped relationships. Ecology, 1998, 79(7): 2555–2559. DOI:10.1890/0012-9658(1998)079[2555:SRABDO]2.0.CO;2 |
| [25] | Kassen R, Buckling A, Bell G, Rainey P B. Diversity peaks at intermediate productivity in a laboratory microcosm. Nature, 2000, 406(6795): 508–512. DOI:10.1038/35020060 |
| [26] | 马文静, 张庆, 牛建明, 康萨如拉, 刘朋涛, 何欣, 杨艳, 张艳楠, 邬建国. 物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例. 植物生态学报, 2013, 37(7): 620–630. |
| [27] | 王长庭, 龙瑞军, 丁路明. 高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响. 生物多样性, 2004, 12(4): 403–409. |
| [28] | 王国杰, 汪诗平, 郝彦宾, 蔡学彩. 水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响. 生态学报, 2005, 25(7): 1649–1656. |
| [29] | 左小安, 赵学勇, 赵哈林, 郭轶瑞, 李玉强, 赵玉萍. 科尔沁沙地草地退化过程中的物种组成及功能多样性变化特征. 水土保持学报, 2006, 20(1): 181–185. |
| [30] | 白永飞, 李凌浩, 王其兵, 张丽霞, 张焱, 陈佐忠. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667–673. |
| [31] | Rajaniemi T K. Explaining productivity-diversity relationships in plants. Okios, 2003, 101(3): 449–457. DOI:10.1034/j.1600-0706.2003.12128.x |
| [32] | 张继义, 赵哈林, 张铜会, 赵学勇. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态. 植物生态学报, 2004, 28(1): 86–92. DOI:10.17521/cjpe.2004.0013 |