 2017, Vol. 37
2017, Vol. 37文章信息
- 邢树文, 朱慧, 马瑞君, 杜颖青, 孙延杰, 查广才
- XING Shuwen, ZHU Hui, MA Ruijun, DU Yingqing, SUN Yanjie, ZHA Guangcai.
- 不同生境条件与管理方式对茶园蜘蛛群落结构及多样性的影响
- Effects of different habitats and management on the spider communities in tea plantations
- 生态学报. 2017, 37(12): 4236-4246
- Acta Ecologica Sinica. 2017, 37(12): 4236-4246
- http://dx.doi.org/10.5846/stxb201604230765
-
文章历史
- 收稿日期: 2016-04-23
- 修订日期: 2016-12-07





近年来, 茶农为追求经济效益提高茶叶产量, 不断扩建茶园并且频繁施用化学农药、除草剂和化肥, 不仅导致常规管理的茶园生态环境遭到严重破坏, 而且引发茶园环境污染和茶叶农药残留超标, 茶叶品质下降。蜘蛛在茶园生态系统中种类多, 数量大, 是主要的茶园害虫天敌, 对茶园多种害虫具有重要的控制作用, 在有机生态茶园害虫调控方面发挥重要功能, 其对假眼小绿叶蝉(Empoasca vitis)具有极强的捕食作用[1-3], 对茶丽纹象甲(Myllocerinus aurolineatus)、茶二叉蚜(Toxoptera aurantii)、黑刺粉虱(Aleurocanthus spiniferus)及多种蛾类低龄幼虫及成虫也具有良好的调控效果[4-7]。因此, 保护茶园蜘蛛多样性, 以蛛控虫, 建立无污染健康的生态茶园是当前学者对茶业种植和发展最关注的热点话题之一。多年来, 为避免化学防治害虫带来的危害, 国内学者在构建茶叶种植和管理模式[8-9], 以及利用蜘蛛等天敌进行生态控制[10-11]方面进行了系统的研究。自20世纪90年代, 我国茶区在蜘蛛种类和地理分布[12-13]、茶园蜘蛛种群动态[14-15]、茶园蜘蛛区系分布[16]、茶园蜘蛛群落组成[17-19]及茶园蜘蛛多样性研究[20]等方面做了大量基础性研究, 为利用蜘蛛种群构建无污染的景观生态茶园奠定了基础, 探讨不同环境条件和管理方式对茶园蜘蛛群落结构及多样性的影响, 蜘蛛群落与茶园环境构建的关系, 对茶业种植、茶叶产业的经济发展和茶区的生态重建具有重要理论意义和实践价值。但是这些方面一直以来都缺乏深入系统研究。本研究以广东省潮州市凤凰山单丛茶区3种不同类型的茶园作为研究样区, 探讨茶园生境条件和管理方式对蜘蛛物种的组成与分布、群落结构及生物多样性的影响, 分析茶园蜘蛛群落结构的差异, 并讨论其原因, 以期为保护天敌资源, 利用蜘蛛天敌控制虫害, 建立以生物防治主的综合防治体系茶园提供基础的科学依据。
1 研究方法 1.1 研究区域概况研究样地设在广东省潮安县境内的凤凰镇, 海拔250—1391 m, 年平均气温为21.4℃, 降雨量为2119.7 mm。极端最高气温为39.6℃, 极端最低气温为-0.5℃。本研究的茶区分有机茶园、无公害茶园和普通茶园3种类型的茶园, 总面积约1000 hm2以上。主要种植茶树品种为凤凰单丛茶, 种植年限在14—16a之间。
有机茶园, 位于凤凰镇叫水坑村, 海拔850 m, 茶园周围是毛竹(Phyllostachys edulis(Carrere) J.Houz.)及阔叶混交林、灌木, 茶园伴生革命菜(Alternanthera sessilis (L.)R.Br.ex DC.)、芒萁(Dicranopteris dichotoma (Thunb.)Bernh.)、野牡丹(Paeonia delavayi Franch.)霍香蓟(Ageratum conyzoides L.)等多种植物。茶园为梯田式造田, 茶树行距2.0m, 间距40cm, 茶树盖度51.3%, 年平均气温19.23℃, 平均湿度(RH) 89.36, 年降雨量为1737.40mm, 每年春季4月人工采春茶1次, 秋后整枝和人工除草, 平均每两年施有机肥1次, 茶园不施用农药与除草剂。
无公害茶园, 位于大山镇上春村、中段村和芼岭村, 海拔510m, 茶园周边为阔叶混交林等乔木、灌木和草丛, 茶园伴生植物主要有金毛耳草(Hedyotis C hrysotricha(Palib)), 芒萁、宽叶鼠麴草(Gnaphalium adnatum Wall.ex DC.)、霍香蓟等植被。梯田式茶园, 茶行间距为2.0m, 或1.5m;茶树盖度76.3%, 平均气温24.46℃, 平均湿度(RH) 86.47, 年平均降雨量为1693.20mm, 茶园每年的4月、6月、8月和10月采茶4次。施用2.5%联苯菊酯35 mL 900倍液及24%灭多威水剂35mL 900倍液, 4—6次/a, 除草剂1次/a。
普通茶园, 位于凤凰镇周边的凤新村、下埠村, 桥头村, 地势为丘陵地带, 海拔350m, 单一茶园, 周围分布有少量小型灌木及杂草, 周边环境严重破坏。茶园伴生极少量鸡矢藤(Paederia scandens (Lour.) Merr.)及空心莲子草(Alternanthera philoxeroides (Mart.) Griseb.)等植被。非梯田式茶园, 茶树行距1.5m。茶树盖度69.32%, 年平均气温26.18℃, 平均湿度(RH) 81.47, 年平均降雨量为1748.20mm, 茶园每年采茶10次。施除草剂1次;施用2.5%联苯菊酯35 mL 900倍液及24%灭多威水剂35mL 900倍液, 12—14次/a, 除草剂2次。
1.2 调查与分析方法 1.2.1 取样时间2011年3—10月份, 对3种不同生境类型的茶园蜘蛛群落进行调查, 每月调查1次, 调查时间为每月的20—25日。标本采集方法为振落承接法和筛网法采集蜘蛛。
1.2.2 样地设置本研究是依据茶园的生境类型和管理方式不同, 在研究区域选择3种不同生境类型和管理方式的茶园(即有机茶园、无公害茶园和普通茶园)作为研究对象, 每个生境茶园各选取海拔和地形差别不大的5块茶园作为调查样地, 共15个调查样地。每个样地面积在约0.3—0.5hm2, 样地间距离均为350m以上。在每个样地采用平行跳跃法, 每隔5个茶行选取一个茶行作为调查样点, 每个样点调查10株茶丛。调查方法先地表, 后茶丛。
地表的调查方法 在每个样点的10株茶丛行间收集地表凋落物和表层土, 采用筛网法对其过筛后捕获筛选出来的蜘蛛。
茶丛的调查方法 在每个采样点的10株茶丛的茶行两侧铺以塑料薄膜承接, 用力振动和拍打茶丛6次, 收集震落下来的蜘蛛。
1.2.3 标本处理与鉴定将每个样点采集的茶丛和地表蜘蛛标本, 分别收集在盛有80%的酒精标本瓶中, 分类贴标签, 带回实验室分别鉴定和计数。所获成体蜘蛛标本鉴定到种, 幼蛛鉴定到科(属), 未鉴定到种的标本不参与数据分析。
1.2.4 优势度划分参照地Berger-Parker生态优势度指数的方法, 将物种生态优势度划分为5个等级, 具体标准为:当D≥0.1时为优势种, 用D(Dominant)表示;0.05≤D<0.1时为丰盛种, 用A(Abundant)表示;0.01≤D<0.05时为常见种, 用F(Frequent)表示;0.001≤D<0.01时为偶见种, 用O(Occasional)表示;D<0.001时为稀有种或罕见种, 用R(Rare)表示[21]。
1.3 分析方法茶园蜘蛛群落的物种多样性(Diversity)采用如下方法计算[22-23]:
Shannon-Wiener指数 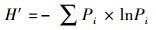
Pielou均匀性指数 E=H′/H′max, 其中, H′mas=lnS
Simpson优势度指数 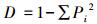
Margalef物种丰富度指数 SR=(S-1)/lnN
式中, pi为群落第i个物种个体数占总个体数比例;N为蜘蛛群落全部物种的总个体数, S为蜘蛛群落的总类群数。
Bray-Curtis距离系数 Sg=(1/S)∑[1-(Xij-Xij|)(1/Ri)]
式中, m为研究样地蜘蛛所有类群数;Xij为第j个生境第i个类群蜘蛛的个体数;Xik为第K个生境i个类群的蜘蛛个体数[24]。
1.4 数据处理本次调查统计的数据均采用Microsoft office Excel 2007和SPSS 16.0进行处理。
在每个生境的茶园蜘蛛群落调查中, 将各个月份相对应同一样地的蜘蛛标本合并计算, 获得5个样地蜘蛛的物种数和个体数, 然后再将5个样地的蜘蛛分别计算, 用以统计蜘蛛的个体数、类群数(科、属)、物种数, 以及群落物种多样性指数(H′), 物种丰富度指数(SR)、Pielou均匀度指数(E)、Simpson优势度指数(D)。参数进行Duncan新复极差多重比较检验, 取α=0.05, 比较各群落参数差异显著性。茶丛和地表蜘蛛分开统计个体数和物种数, 并按上述方法分别计算其群落物种多样性各项生态学指标和Duncan检验。
采用非线性多维标度法(non-metric multidimensional scaling, NMDS)对3种茶园蜘蛛群落进行了分类排序。在本文中, NMDS排序所使用的数据库是每个样方蜘蛛的活动密度, 排序分析过程选用Sorensen (Bray-Curtis)指数来比较蜘蛛群落结构的差异。排序分析结果给出了Stress值, Stress < 5, 表示排序结果很好;Stress < 10, 表示排序结果较好;Stress < 20, 表示排序结果尚可;Stress > 20, 则表示排序结果较差[25]。利用非参数多元方差分析比较3种茶园蜘蛛群落结构的整体差异。非参数的多元方差分析也称为基于置换的多元方差分析(Permutation based MANOVA, perMANOVA)是一种利用置换方法来检验多元方差分析的非参数程序, 适用于动物生态学群落分析。本文perMANOVA用于检验3种不同茶园蜘蛛群落整体上是否存在差异, 以茶园类型为分组变量, 用Sorensen (Bray-Curtis)做距离测量。perMANOVA的结果以传统方差分析表的形式输出。以上分析使用国际通用的排序软件PC-ORD 5.0软件包[26]。
2 结果与分析 2.1 不同生境条件与管理方式的茶园蜘蛛群落组成与数量特征调查统计结果如表 1所示, 3种茶园共采集蜘蛛标本15992头, 隶属18科62属79种。其中, 有机茶园蜘蛛个体数8076头, 隶属18科58属73种, 占总个体数的50.22%, 占总物种数的92.41%;无公害茶园蜘蛛个体数4911头, 隶属17科49属64种, 占总个体数的30.88%, 占总物种数的81.01%;普通茶园蜘蛛个体数3005头, 隶属14科42属46种, 占总个体数的18.90%, 占总物种数的58.22%。
| 科Family | 有机茶园 Organic tea plantation |
无公害茶园 Pollution-free tea plantation |
普通害茶园 Common tea plantation |
|||||||||||
| 物种数 Species number |
优势度/% Dominance |
个体数 Individual Number |
优势度/% Dominance |
物种数 Species number |
优势度/% Dominance |
个体数 Individual Number |
优势度/% Dominance |
物种数 Species number |
优势度/% Dominance |
个体数 Individual Number |
优势度/% Dominance |
|||
| 球蛛科Theridiidae | 10 | 13.70 | 1091 | 13.66 | 9 | 14.06 | 553 | 11.26 | 7 | 15.22 | 198 | 6.59 | ||
| 园蛛科Araneidae | 3 | 4.11 | 565 | 7.07 | 3 | 4.69 | 233 | 4.74 | 2 | 4.35 | 126 | 4.19 | ||
| 皿蛛科Linyphiidae | 8 | 10.96 | 228 | 2.85 | 7 | 10.94 | 212 | 4.32 | 5 | 10.87 | 193 | 6.42 | ||
| 肖蛸科Tetragnathidae | 3 | 4.11 | 377 | 4.72 | 3 | 4.69 | 390 | 7.94 | 2 | 4.35 | 131 | 4.36 | ||
| 异纺蛛科Hexathelidae | 1 | 1.37 | 136 | 1.70 | 1 | 1.56 | 144 | 2.93 | 0 | 0.00 | 0 | 0.00 | ||
| 漏斗蛛科Agelenidae | 2 | 2.74 | 301 | 3.77 | 2 | 3.13 | 265 | 5.40 | 1 | 2.17 | 132 | 4.39 | ||
| 栅蛛科Hahniidae | 1 | 1.37 | 495 | 6.20 | 1 | 1.56 | 367 | 7.47 | 1 | 2.17 | 406 | 13.51 | ||
| 跳蛛科Salticidae | 12 | 16.44 | 1102 | 13.80 | 10 | 15.63 | 513 | 10.45 | 9 | 19.57 | 263 | 8.75 | ||
| 逍遥蛛科Philodromidae | 1 | 1.37 | 81 | 1.01 | 1 | 1.56 | 30 | 0.61 | 0 | 0.00 | 0 | 0.00 | ||
| 管巣蛛科Clubionidae | 3 | 4.11 | 1129 | 14.14 | 2 | 3.13 | 511 | 10.41 | 1 | 2.17 | 452 | 15.04 | ||
| 猫蛛科Oxyopidae | 2 | 2.74 | 213 | 2.67 | 2 | 3.13 | 214 | 4.36 | 1 | 2.17 | 123 | 4.09 | ||
| 蟹蛛科Thomisidae | 7 | 9.59 | 1058 | 13.25 | 6 | 9.38 | 529 | 10.77 | 4 | 8.70 | 215 | 7.15 | ||
| 平腹蛛科Gnaphosidae | 2 | 2.74 | 154 | 1.93 | 2 | 3.13 | 124 | 2.52 | 2 | 4.35 | 136 | 4.53 | ||
| 狼蛛科Lycosidae | 10 | 13.70 | 935 | 11.70 | 9 | 14.06 | 641 | 13.05 | 8 | 17.39 | 438 | 14.58 | ||
| 圆颚蛛科Corinnidae | 3 | 4.11 | 4 | 0.05 | 2 | 3.13 | 2 | 0.04 | 0 | 0.00 | 0 | 0.00 | ||
| 米图蛛科Miturgidae | 2 | 2.74 | 61 | 0.76 | 2 | 3.13 | 13 | 0.26 | 1 | 2.17 | 51 | 1.70 | ||
| 长纺蛛科Hersiliidae | 1 | 1.37 | 6 | 0.8 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | ||
| 卵形蛛科Oonopidae | 2 | 2.74 | 41 | 0.51 | 2 | 3.13 | 170 | 3.46 | 2 | 4.35 | 141 | 4.69 | ||
| 合计Total | 73 | 100.00 | 8076 | 100.00 | 64 | 100.00 | 4911 | 100.00 | 46 | 100.00 | 3005 | 100 | ||
| 优势度的划分:优势类群(Dominant Order):>10.0%;丰富类群(Rich groups):5%―10%;常见类群(Frequent Order):1%―5%;稀有类群(Rare Order):<1% | ||||||||||||||
在科的水平上, 本次调查的3类茶园共有优势类群是管巢蛛科和狼蛛科, 占总科数的11.11%。其中, 有机茶园与无公害茶园优势类群相同, 包括球蛛科、跳蛛科、管巢蛛科、蟹蛛科和狼蛛科, 分别占总科数的27.78%和29.41%;普通茶园优势类群3科, 包括管巢蛛科、狼蛛科和栅蛛科, 占总科数的23.08%。优势类群、丰盛类群、常见类群和稀有类群之比分别是有机茶园5:2:7:4, 无公害茶园5:3:6:3, 普通茶园3:4:7:0, 由此看出, 有机茶园和无公害茶园蜘蛛群落的组成与结构优势于普通茶园。
在属的水平上, 球蛛科、皿蛛科、跳蛛科和狼蛛科的属较多, 属的优势度均高于10%以上。有机茶园蜘蛛比无公害茶园多9个属, 比普通茶园多12个属。有机茶园占总属数的93.55%, 无公害茶园占总属数的79.03%, 普通茶园占总属数的67.74%。
在物种的水平上, 有机茶园比无公害茶园多9种, 比普通茶园多27种。3类茶园的球蛛科、跳蛛科、狼蛛科和皿蛛科的蜘蛛种类最多, 物种优势度均大于10%。其次, 蟹蛛科物种优势度较高, 有机茶园9.59%, 无公害茶园9.38%和普通茶园8.70%。物种名称详见附表:广东潮州凤凰山茶园蜘蛛资源名录。
| 科Family-属genus | 种Species | OP | PP | CP | |||||
| 茶丛 | 地表 | 茶丛 | 地表 | 茶丛 | 地表 | ||||
| 球蛛科Theridiidae | |||||||||
| 鞘腹蛛属Coleosoma | 1.滑鞘腹蛛* Coleosoma blandum O. P. -Cambridge | D | A | A | |||||
| 2.佛罗鞘腹蛛*Coleosoma floridanum Banks | O | O | |||||||
| 3.八斑鞘腹蛛*Coleosoma octomaculatum (Boesenberg & Strand) | A | A | F | ||||||
| 丽蛛属Chrysso | 4.黑丽蛛* Chrysso nigra (O. P. -Cambridge) | F | F | ||||||
| 格蛛属Coscinida | 5.亚洲格蛛* Coscinida asiatica Zhu & Zhang | Rs | O | R | R | ||||
| 球蛛属Theridion | 6.漏斗球蛛* Theridion chonetum Zhu | F | O | O | F | ||||
| 后丘蛛属Dipoenura | 7.旋转后丘蛛* Dipoenura cyclosoides (Simon, ) | F | O | ||||||
| 丘腹蛛属Episinus | 8.驼背丘腹蛛* Episinus gibbus Zhu & Wang | A | A | A | |||||
| 圆腹蛛属Phycosoma | 9.花圆腹蛛*Phycosoma hana (Zhu) | F | O | F | F | ||||
| 10.玛丁圆腹蛛*Phycosoma martinae (Roberts) | O | ||||||||
| 斯坦蛛属Stemmops | 11.日斯坦蛛*Stemmops nipponicus Yaginuma | F | R | F | O | R | |||
| 蟹蛛科Thomisidae | |||||||||
| 蟹蛛属Thomisus | 12.角红蟹蛛Thomisus labefactus Karsch | F | O | F | O | F | O | ||
| 13.冲绳蟹蛛* Thomisus okinawensis Strand | F | O | R | O | O | O | |||
| 微蟹蛛属Lysiteles | 14.梅微蟹蛛* Lysiteles maius Ono | A | O | F | O | F | O | ||
| 花蟹蛛属Xysticus | 15.千岛花蟹蛛* Xysticus kurilensis Strand | F | O | F | O | F | O | ||
| 16.波纹花蟹蛛Xysticus croceus Fox | A | O | A | A | |||||
| 锯足蛛属Runcinia | 17.近亲锯足蛛* Runcinia affinis Simon | F | O | O | O | ||||
| 三门蟹蛛属Pharta | 18.郑氏三门蟹蛛* Pharta brevipalpus (Simon) | R | O | R | O | R | |||
| 逍遥蛛科Philodromidae | |||||||||
| 逍遥蛛属Philodromus | 19.胡氏逍遥蛛* Philodromus hui Yang & Mao | F | F | O | |||||
| 栅蛛科Hahniidae | |||||||||
| 栅蛛属Hahnia | 20.栓栅蛛Hahnia corticiola Boesenberg et strand | D | O | A | A | ||||
| 异纺蛛科Hexathelidae | |||||||||
| 大疣蛛属Macrothele | 21.触形大疣蛛* Macrothel palpator Pocock | R | O | ||||||
| 肖峭科Tetragnathidae | |||||||||
| 银鳞蛛属Leucauge | 22.肩斑银鳞蛛Leucauge blanda L.koch | A | A | A | |||||
| 肖峭属Tetragnatha | 23.羽斑肖峭* Tetragnatha pinicola L. Koch | A | A | A | |||||
| 24.前齿肖峭Tetragnatha praedonia L.Koch | R | R | O | ||||||
| 平腹蛛科Gnaphosidae | |||||||||
| 平腹蛛属Gnaphosa | 25.金比罗平腹蛛* Gnaphosa kompirensis Boesenberg & Strand | F | F | F | |||||
| 狂蛛属Zelotes | 26.通道狂蛛* Zelotes tongdao Yin, Bao & Zhang | R | O | O | |||||
| 圆颚蛛科Corinnidae | |||||||||
| 奥塔蛛属Otacilia | 27.尖峰岭奥塔蛛* Otacilia jianfengling Fu, Zhang & Zhu | O | R | ||||||
| 盾球蛛属Orthobula | 28.福建盾球蛛* Orthobula fujian sp.nov. | R | O | O | |||||
| 刺蛛属Echinax | 29.拟猫刺蛛Echinax Oxyopoides Deeleman-Reinhold | R | R | O | |||||
| 长纺蛛科Hersiliidae | |||||||||
| 长纺蛛属Hersilia | 30.波纹长纺蛛Hersilia dtriata Wang & Yin | O | O | ||||||
| 皿蛛科Linyphiidae | |||||||||
| 贡奥蛛属Gongylidioides | 31.小野贡奥蛛*Gongylidioides onoi Tazoe | O | O | O | |||||
| 沟瘤蛛属Ummeliata | 32.食虫沟瘤蛛* Ummeliata insecticeps (Boesemberg & Strand) | A | O | A | O | A | |||
| 微蛛属Erigone | 33.隆背微蛛*Erigone prominens (Boesenberg & Strand) | F | R | R | O | O | R | ||
| 角微蛛属Ceratinella | 34.普氏角微蛛*Ceratinella plancyi (Simon) | O | R | O | R | O | O | ||
| 额毛蛛属Caviphantes | 35.三齿额毛蛛*Cariphantes samensis Oi | O | R | O | O | ||||
| 面蛛属Prosoponoides | 36.中华面蛛*Prosoponoides sinensis (Chen) | R | R | R | |||||
| 玲蛛属Parameionetat | 37.二叶玲蛛*Parameioneta bilobata Li & Zhu | R | R | R | |||||
| 盖蛛属Neriene | 38.卡氏盖蛛Neriene cavaleriei (Schenkel) | F | O | F | O | F | |||
| 须蛛属Paranasoona | 39.前须蛛*Paranasoona cirrifrons (Heimer) | O | O | O | |||||
| 跳蛛科Salticidae | |||||||||
| 金蝉蛛属Phintella | 40.多色金蝉蛛Phintella versicolor (C.L.Koch) | A | A | F | A | A | F | ||
| 兜跳蛛属Ptocasius | 41.毛垛兜跳蛛*Ptocasius strupifer Simon | A | F | A | F | F | F | ||
| 斑蛛属Euophrys | 42.前斑蛛*Euophrys frontalis (Walckenaer) | F | O | F | O | F | O | ||
| 西菱头蛛属Sibianor | 43.斜纹西菱头蛛*Sibianor aurocinctus (Ohlert) | A | F | A | F | F | O | ||
| 哈莫蛛属Harmochirus | 44.鳃哈莫蛛*Harmochirus brachiatus (Thorell) | O | R | R | |||||
| 45.巴莫方胸蛛*Thiania bhamoensis Thorell | F | F | F | F | O | O | |||
| 46.细齿方胸蛛*Thiania suboppressa Strand | A | F | F | F | O | O | |||
| 布氏蛛属Bristowia | 47.巨刺布氏蛛*Bristowia heterospinosa Reimoser | O | R | O | R | O | O | ||
| 华蛛属Chinattus | 48.胫节华蛛*Chinattus tibialis (Zabka) | O | O | O | O | ||||
| 哈沙蛛属Hasarius | 49.花哈沙蛛Hasarius adansoni (Audouin) | O | F | O | F | F | O | ||
| 丽跳蛛属Chrysilla | 50.华美丽跳蛛*Chrysilla lauta Thorell | F | F | F | F | O | O | ||
| 蝇象属Hyllus | 51.斑腹蝇象蛛*Hyllus diardi (Walckenaer) | O | R | O | R | R | |||
| 拟伊蛛属Pseudicius | 52.四川拟伊蛛*Pseudicius szechuanensis Logunov | O | R | ||||||
| 狼蛛科Lycosidae | |||||||||
| 豹蛛属Pardosa | 53.沟渠豹蛛Pardosa laura Karsch | O | D | O | D | O | A | ||
| 54.拟环纹豹蛛Pardosa pseudoannulata (Boesenberg et Strand) | O | D | O | D | A | ||||
| 55.幼豹蛛*Pardosa pusiola (Thorell) | O | F | F | O | F | ||||
| 涡蛛属Wadicosa | 56.忠涡蛛*Wadicosa fidelis (O. P. -Cambridge) | O | O | O | |||||
| 水狼蛛属Ocyale | 57.细毛水狼蛛*Pirata tenuisetacea (Chai) | O | F | F | O | ||||
| 58.拟水狼蛛Pirata subpiraticus (Boesenberg et strand) | O | D | O | Ds | O | F | |||
| 脉狼蛛属Venonia | 59.旋曩脉狼蛛Venonia spirocysta Chai | O | F | F | F | ||||
| 狼蛛属Lycosa | 60.格氏狼蛛*Lycosa grahami Fox | O | F | O | F | F | |||
| 熊蛛属Arctosa indicus | 61.印熊蛛*Arctosa indica Tikader & Malhotra | O | A | O | A | F | |||
| 獾蛛属Trochosa | 62.版纳獾蛛*Trochosa bannaensis Yin & Chen | O | A | O | A | O | |||
| 马蛛属Hippasa | 63.格里马蛛*Hippasa greenalliae (Blackwall) | O | O | O | |||||
| 卵形蛛科Oonopidae | |||||||||
| 巨膝蛛属Opopaea | 64.角巨膝蛛*Opopaea cornuta Yin & Wang | F | F | F | |||||
| 65.羽巨膝蛛*Opopaea plumula Yin & Wang | F | O | O | ||||||
| 奥蛛属Orchestina | 66.纹胸奥蛛*0rchestina thoracica Xu | R | R | ||||||
| 漏斗蛛科Agelenidae | |||||||||
| 异漏斗蛛属Allagelena | 67.机敏异漏斗蛛Allagelena difficilis (Fox) | F | F | F | |||||
| 漏斗蛛属Agelena | 68.森林漏斗蛛*Agelena silvatica Oliger | F | O | F | F | ||||
| 猫蛛科Oxyopidae | 69.斜纹猫蛛Oxyopes sertatus L.Koch | F | O | F | O | F | O | ||
| 猫蛛属Oxyopes | 70.爪哇猫蛛Oxyopes javanus Thorell | F | O | F | F | O | |||
| 71.类斜纹猫蛛*Oxyopes sertatoides Xie & Kim | F | F | F | ||||||
| 米图蛛科Miturgidae | |||||||||
| 红螯蛛属Cheiracanthium | 72.短距红螯蛛* Cheiracanthium Brevicalcaratum L. Koch | F | O | F | O | F | O | ||
| 73.钩红螯蛛Cheiracanthium unicum (Boesenberg et strand) | F | O | F | O | F | O | |||
| 管巢蛛科Clubionidae | |||||||||
| 管巢蛛属Clubiona | 74.羽斑管巢蛛*Clubiona Jucunda (Karsch) | F | O | F | O | ||||
| 75.谷川管巢蛛* Clubiona tanikawai Ono | F | O | F | O | O | ||||
| 76.盔黄树蛛* Xantharia galea Zhang & Fu | R | R | |||||||
| 园蛛科Araneidae | |||||||||
| 艾蛛属Cyclosa | 77.浊斑艾蛛* Cyclosa confusa (Boesenberg & Strand) | F | F | F | |||||
| 78.日本艾蛛* Cyclosa japonica (Boesenberg & Strand) | F | F | F | ||||||
| 八氏蛛属Yaginumia | 79.叶斑八氏蛛* Yaginumia sia (Strand) | F | F | O | |||||
| *表示广东茶区蜘蛛新纪录种;D:优势种Dominnant;A:丰盛种Abundant; F:常见种Frequent;O:偶见种Occational;R:稀有种Rare; OP:有机茶园Organic tea platation;PP:无公害茶园Non-Pollution tea platation;CP:普通茶园Commen tea platation | |||||||||
运用物种多样性方法, 对不同类型茶园蜘蛛群落数据处理, 见表 2所示。结果表明:3种不同类型茶园蜘蛛的个体数、物种数、多样性指数、均匀度指数、优势度指数和丰富度指数均表现为有机茶园高, 无公害茶园次之, 普通茶园低, 差异达到显著水平(P<0.05)。有机茶园蜘蛛个体数、物种数与普通茶园的相比差距较大, 与无公害茶园差距较小, 说明有机茶园蜘蛛的优势度和丰富度远高于普通茶园, 与优势性指数和丰富度指数相一致。3个茶园的优势性都很高, 说明茶园蜘蛛是茶园害虫的主要天敌。有机茶园Shannon-Wiener多样性指数高于无公害茶园1.5倍, 高于常规管理的普通茶园的2.40倍,表明有机茶园蜘蛛群落结构稳定性好, 普通茶园相对较差。
| 茶园类型 Tea plantation |
有机茶园 Organic tea plantation |
无公害茶园 Pollution-free tea plantation |
普通茶园 Common tea plantation |
| 个体数Individual number | 1615.20±95.04a | 982.20±80.03b | 601.00±54.90c |
| 物种数Species number | 57.80±3.90a | 48.20±4.49b | 34.00±2.74c |
| 多样性指数Shannon-Wiener diversity index | 3.51±0.20a | 2.30±0.13b | 1.46±0.09c |
| 均匀度指数Pielou evenness index | 0.86±0.04a | 0.60±0.034b | 0.42±0.03c |
| 优势性指数Simpson dominance index | 0.99±0.02a | 0.98±0.03b | 0.92±0.01c |
| 丰富度指数Magarlef richness index | 7.72±0.56a | 6.83±0.68b | 5.23±0.54c |
| 同一行不同小写字母表示有显著差异(P<0.05) | |||
茶园生态系统、周边生境条件及茶园管理方式对蜘蛛群落的个体数量和物种组成有很大影响。本次调查对茶丛和地表蜘蛛统计的数据显示, 有机茶园、无公害茶园和普通茶园的地表与茶丛蜘蛛物种比值分别为1:1.24, 1:1.27, 1:1.02, 个体数比值为1:1.74, 1:1.43, 1:1.29。由此看出, 蜘蛛的个体数、物种数及相对丰度均表现为茶丛高于地表, 有机茶园高, 无公害茶园次之, 普通茶园低;除园蛛科、肖蛸科、漏斗蛛科的蜘蛛在茶树上结网外, 跳蛛科、蟹蛛科、管巢蛛科、猫蛛科及皿蛛科的大多数蜘蛛游猎于茶树上狩猎昆虫, 少数在地面上捕猎, 所以茶丛蜘蛛的丰富度、多度均高于地表。由于游猎蜘蛛的活动范围大, 在茶丛和地表上都有游猎蜘蛛活动, 说明游猎蜘蛛在茶园捕食功能的重要性。而3类茶园茶丛与地表蜘蛛的差异, 却与茶园的周边生境和茶园自身的生态环境相关, 周边生境与自身生态环境良好的有机茶园适宜更多数量和种类的蜘蛛聚集和生存, 生态环境较差的普通茶园蜘蛛的数量和种类则减少。
从表 3可以看出, 3种类型茶园茶丛和地表蜘蛛的个体数、物种数均表现为有机茶园高, 无公害茶园次之, 普通茶园低, 且达到显著水平(P<0.05)。茶丛蜘蛛群落的物种多样性指数、均匀度指数、优势性指数和丰富度指数均表现有机茶园高, 无公害茶园次之, 普通茶园低, 且差异性均达到显著水平(P<0.05);3类茶园地表蜘蛛群落的个体数和多样性指数差异达到显著水平(P<0.05), 普通茶园的物种数和丰富度指数与无公害茶园间无显著差异, 有机茶园和无公害茶园的均匀度指数无显著差异, 但与普通茶园差异显著;3类茶园的优势性指数无显著差异。整体上, 茶丛蜘蛛群落结构和物种多样性高于地表蜘蛛群落, 有机茶园地表与茶丛蜘蛛群落结构的相差较大, 无公害茶园与普通茶园地表与茶丛蜘蛛相差较小。
| 指标Index | 茶丛Tea bush | 地表Ground surface | |||||
| 有机茶园 Organic tea plantation |
无公害茶园 Pollution-free tea plantation |
普通茶园 Common tea plantation |
有机茶园 Organic tea plantation |
无公害茶园 Pollution-free tea plantation |
普通茶园 Common tea plantation |
||
| 个体数Individual number | 1025.60±45.90a | 578.40±65.05b | 307.40±63.018c | 589.60±101.94a | 403.80±50.52b | 292.20±33.83c | |
| 物种数Species number | 41.80±2.39a | 33.00±2.45b | 23.40±1.14c | 33.80±3.27a | 26.00±2.83b | 22.80±3.49b | |
| 多样性指数Diversity index | 3.37±0.062a | 3.14±0.07b | 2.73±0.07c | 2.58±0.15a | 2.36±0.10b | 2.19±0.23c | |
| 均匀度指数Evenness index | 0.90±0.01a | 0.89±0.01a | 0.79±0.02b | 0.79±0.02a | 0.78±0.04a | 0.73±0.03b | |
| 优势性指数Dominance index | 0.96±0.04a | 0.94±0.04b | 0.92±0.01c | 0.88±0.02a | 0.88±0.02a | 0.86±0.04a | |
| 丰富度指数Richness index | 5.89±0.35a | 5.04±0.43b | 3.81±0.27c | 5.15±0.49a | 4.37±0.45b | 3.79±0.59b | |
3种茶园蜘蛛群落NMDS排序分析结果表明, 排序轴1和2存在明显的正交性(84.3%, r=-0.40), 这两个排序轴累计解释了95.7%的数据变异(排序轴1:R2 = 0.888;排序轴2:R2=0.069), 说明前两个排序轴较好地反映了茶丛蜘蛛群落结构(图 1), 表明茶丛蜘蛛群落的个体数、物种数和多样性指数、丰富度指数高于地表。3种茶园蜘蛛群落NMDS排序分析计算出的Stress值为4.81, 表明NMDS排序分析结果是较好的。从排序图中可以看出, 3种茶园蜘蛛群落组成明显不同, 有机茶园蜘蛛群落组成与普通茶园相差较大, 而无公害茶园与普通茶园蜘蛛群落组成相差较小。3种茶园蜘蛛群落的方差(PerMANOVA)分析结果与NMDS排序结果一致, 3种茶园蜘蛛群落的个体数、物种数和生态指数均存在显著差异(F2, 14=12.46, P < 0.001)。多重比较的结果表明, 3种茶园间蜘蛛群落的个体数、物种数和蜘蛛群落生态学指数均存在显著差异(t=2.95, P=0.011;t =4.91, P=0.010;t=2.53, P=0.008)。

|
| 图 1 茶园蜘蛛群落NMDS排序图 Fig. 1 NMDS plots indicating 2-dimensional distances of the spider communities among three habitats |
生物多样性作为生物群落组成结构的重要指标, 能够反映群落内物种多少和生态系统的复杂程度, 从而反映生境间的相似性及差异性[27-28]。生物群落多样性是群落生态学研究的热点之一, 越来越多的研究表明, 复杂多样的植被环境蜘蛛群落多样性也增加[29-30]。蜘蛛群落多样性与茶园生态系统周边自然环境条件和外界人为干扰相互适应, 蜘蛛各类群在时空序列上产生与周围环境相适应的周期性节律动态, 并且协同进化[31]。茶园周边的生境条件、地理位置、温度、湿度、管理方式的差异, 以及蜘蛛与害虫间的食性关系, 是导致茶园蜘蛛群落结构产生差异的重要因素[32]。蜘蛛种类在茶园生态系统的构成受周边生境条件等影响较大, 对不同生境具有依赖性, 随着生境的改善群落密度变大[3, 32]。本研究结果有机茶园、无公害茶园和普通茶园蜘蛛的个体数之比为2.69:1.64:1, 物种数量比为1.59:1.14:1, 有机茶园蜘蛛的多样性高、稳定性强, 无公害茶园次之, 而普通茶园蜘蛛多样性低, 稳定性差。主要有两个因素, 其一是不同茶园生态环境及周边生境存在很大的差异。有机茶园周边植为阔叶混交林和竹林, 其交界处具有小灌木和草丛, 丰富的植被形成茶园周边复杂的生态环境, 构成茶园与植被缓冲带间的边缘效应[33], 对维持该环境的小气候稳定性起到关键作用。茶园周边丰富而稳定的植被生境改善了节肢动物的生态位和空间分布格局, 不仅为蜘蛛提供了良好的庇护场所, 小气候适宜的温度和湿度也为蜘蛛繁殖和生存提供了食物和栖息的条件[34], 因此该茶园蜘蛛种类和个体数都会增加, 物种多样性也有所提高, 茶园生态结构更加稳定。而普通茶园是过度开发茶园, 周边生态环境遭到严重的破坏, 茶园连成片, 形成单一茶园, 蜘蛛由于缺乏庇护场所, 夏季(7月份和8月份)形成高温, 秋季(9月下旬至11月上旬)因湿度降低形成干燥气候, 以及冬季低温对蜘蛛的捕食和繁殖造成很大的影响, 导致蜘蛛群落数量减少, 多样性降低。其二是不同茶园的管理方式不同。有机茶园人为干扰少, 有机茶园不使用化学农药和除草剂, 每年春季4月上旬至5月下旬采茶1次, 全年整枝1次, 普通茶园人为干扰多, 尤其是在4月、5月和6月使用农药, 不仅会杀死成熟蜘蛛, 而且对会杀死大量的幼蛛, 大大减少蜘蛛的种群数量, 这也是普通茶园蜘蛛数量和种类少于有机茶园的主要原因。其三, 茶园的地理位置、海拔高度等自然条件, 气温和湿度、降雨量等气候条件也是导致茶园蜘蛛群落多样性差异的重要因素。有机茶园海拔高, 生态环境结构复杂, 降雨量增多, 雾气增加, 茶园形成适宜的气温和湿度, 避免高温气候, 有利于蜘蛛的繁殖和生存, 提高茶园蜘蛛的物种数量及多样性, 这对控制茶园虫害、调控茶园生态系统的稳定起着重要的作用。而无公害茶园海拔较低, 尤其是普通茶园海拔只有350m, 茶园周边生境严重破坏, 夏季的高温、秋季的干燥以及冬季低温都会降低茶园蜘蛛种群数量和多样性。无公害茶园周边也具有良好的生境条件, 阔叶混交林、灌木丛和杂草等丰富的植被分布, 形成茶园的小气候条件, 会聚集更多种类和数量的蜘蛛, 提高蜘蛛群落的物种多样性, 但由于该茶园使用化学农药和除草剂, 会杀死或驱赶部分蜘蛛逃离茶园, 导致茶园蜘蛛种类和个体数都会减少, 因此, 无公害茶园蜘蛛群落的多样性及蜘蛛群落结构的稳定程度会低于有机茶园, 高于普通茶园。
3.2 茶园周边良好的生境条件及科学的管理方式, 能增加茶丛和地表蜘蛛的种类和数量, 有效促进蜘蛛在茶丛和地表上的合理分布, 提高茶园蜘蛛优势类群蜘蛛的捕食和生殖方式是自然选择的结果, 也是蜘蛛在获取这种本能中竞争生存空间环境的过程。生境条件的优劣, 对蜘蛛获取食物和繁殖后代有着重大影响[35]。茶园周边生境条件、茶园小生态系统植被组成、植被生物量及植被生态系统的相对稳定性等直接影响蜘蛛群落的组成和分布[36-38]。本次调查的3种类型茶园茶丛蜘蛛的种类和个体数均高于地表的1.5倍以上, 而且茶丛和地表蜘蛛群落的丰富度和物种多样性均为有机茶园高于无公害茶园, 普通茶园最低。生境条件和管理方式是茶园栖息的蜘蛛群落空间分布的重要因素, 周边生境植被丰富稳定性好、有植被缓冲带的有机茶园, 栖息大量的节肢动物群落, 丰富了茶树地上部分的食物链结构, 大量的游猎型蜘蛛, 如多色金蝉蛛、鳃哈莫蛛、花哈沙蛛、兜垛跳珠、栓栅蛛、食虫沟瘤蛛、卡氏盖蛛等与狩猎型蜘蛛, 如角红蟹蛛、千岛花蟹蛛、羽斑管巢蛛、斜纹猫蛛等聚集在茶树干和树冠上, 导致茶丛蜘蛛群落的物种丰富, 数量增加, 多样性升高。而有机茶园茶行间不仅凋落物多, 地表覆盖度高, 而且茶园植被也较其他两类茶园丰富, 为沟渠豹蛛、幼豹蛛、版纳熊蛛、胡氏逍遥蛛等地表游猎型蜘蛛提供了栖息和捕食场所, 同样导致地表游猎型蜘蛛种类和数量的增加, 多样性升高。另外, 有机茶园为无害管理, 不使用农药和除草剂, 对茶园干扰少, 以及适宜的气温, 使结网型蜘蛛种群数量增多, 整体提升了有机茶园蜘蛛多样性, 为茶园害虫的生态调控起到了关键性的作用。无公害茶园虽然也具有良好的周边环境, 但农药和除草剂的使用, 很大程度上削弱了蜘蛛种群数量, 降低了蜘蛛群落物种多样性。周边裸露、没有植被分布的普通茶园, 其自身生态环境也较差, 茶丛蜘蛛种类和数量会大量减少, 由于地表覆盖层少, 大部分裸露, 栖息环境和食物来源都不能满足地表蜘蛛的生存需求, 因而导致地表蜘蛛物种与数量减少, 物种多样性降低。此外, 普通茶园频繁使用化学农药和除草剂, 采茶次数等人为干扰活动多, 茶树地上部分的结网蜘蛛, 如八斑鞘腹蛛、滑鞘腹蛛、花圆腹蛛、浊斑艾蛛、日本艾蛛、羽斑肖蛸、前齿肖蛸、森林漏斗蛛和机敏漏斗蛛数量会大量减少, 同时导致游猎或狩猎型的跳蛛科、蟹蛛科和个体较小的皿蛛科的种群数量大量减少, 因而很大程度上降低了蜘蛛的多样性。
4 结论本研究是对广东凤凰单丛茶区的首次调查, 记录了茶园蜘蛛79种, 补充广东茶园蜘蛛新纪录61种[11], 其中有机茶园占全部物种的98.53%, 其茶丛蜘蛛和地表蜘蛛优势种、个体数及生物多样性各项生态指标均大于无公害茶园和普通茶园[39-40]。可见, 周边具有良好的生境条件、生境复杂而结构稳定的有机茶园, 能提高蜘蛛群落结构的稳定性和多样性, 合理构建茶园周边生境并对其进行科学管理, 不使用化学农药, 构建结构复杂植物相对丰富的生境缓冲带, 能增加天敌蜘蛛群落多样性, 发挥蜘蛛对害虫叶蝉自然控制作用, 从而提高茶叶质量和经济效益, 科学管理方式是保证茶树干和树冠游猎蜘蛛群落多样性与稳定性的重要因素。因此保护茶园周边生境, 减少茶园的人为干扰, 能够提高茶园蜘蛛种群密度, 丰富蜘蛛种类, 优化和稳定茶园蜘蛛群落结构, 这是实现茶园害虫生态控制的有效途径。
| [1] | 王沅江, 谢振伦, 庞雄飞. 假眼小绿叶蝉及天敌蜘蛛生态位的研究. 茶叶科学, 2008, 28(6): 401–406. |
| [2] | 戴轩. 假眼小绿叶蝉的天敌蜘蛛种类及其物种多样性研究. 昆虫天敌, 1999, 21(4): 164–169. |
| [3] | 黎秀娣, 冯平万, 黎健龙, 唐颢, 唐劲驰. 茶树上游猎型蜘蛛功能团对景观低碳管理模式的反应. 生态环境学报, 2014, 23(1): 64–72. |
| [4] | 张汉鹄, 谭济才. 中国茶树害虫及其无公害治理. 合肥: 安徽科学技术出版社, 2004. |
| [5] | 石春华. 茶园病虫害防治彩色图说. 北京: 中国农业出版社, 2006. |
| [6] | 靳永, 谢玉, 张波涛, 邵永, 杨列芳. 茶黑刺粉虱的生物学特性及防治. 昆虫知识, 2007, 44(5): 734–736. DOI:10.7679/j.issn.2095-1353.2007.167 |
| [7] | 陈亦根, 黄明度. 茶尺蠖生物防治进展. 昆虫天敌, 2001, 23(4): 181–184. |
| [8] | 庞冬辉, 肖润林, 侯柏华, 单武雄, 陈佩. 生态管理对茶园节肢动物群落结构和多样性的影响. 中国生态农业学报, 2010, 18(6): 1272–1276. |
| [9] | 黎健龙, 黎华寿, 黎秀娣, 刘杰, 李家贤, 唐颢, 唐劲驰. 广东英德茶区蜘蛛群落结构及多样性研究. 茶叶科学, 2014, 34(3): 253–260. |
| [10] | 洪海林, 刘恒贵, 沈毓芬, 饶辉福, 施仕胜. 茶园蜘蛛的种类、消长、控害作用与保护利用. 中国茶叶, 2010, 32(2): 8–9. |
| [11] | 邓欣, 谭济才. 生态控制茶园内害虫、天敌种类及数量的季节变化规律. 生态学报, 2002, 22(7): 1166–1172. |
| [12] | 朱璧然, 黎健龙, 刘杰, 唐劲驰, 李家贤, 唐颢, 黎秀娣. 广东茶区蜘蛛新纪录及其地理分布. 广东农业科学, 2011(13): 63–64. DOI:10.3969/j.issn.1004-874X.2011.13.023 |
| [13] | 邢树文, 林瑞芬, 詹珍旋. 广东潮州凤凰山茶区蜘蛛名录及其地理分布. 蛛形学报, 2014(2): 116–122. |
| [14] | 曾明森, 吴光远, 王庆森, 余素红. 福建茶园蜘蛛种类与种群动态的初步研究. 贵州科学, 2008, 26(2): 34–38. |
| [15] | 王定锋, 刘丰静, 李慧玲, 周维, 曾明森, 王庆森, 吴光远. 宁德市低海拔茶园蜘蛛群落结构及发生动态. 茶叶科学, 2013, 33(5): 457–464. |
| [16] | 戴轩, 韩宝瑜. 贵州省茶园蜘蛛区系分布特征. 生态学报, 2009, 29(5): 2356–2367. |
| [17] | 韩宝瑜. 有机、无公害和普通茶园蜘蛛群落组成及动态差异. 蛛形学报, 2005, 14(2): 104–107. |
| [18] | 谭济才, 邓欣, 袁哲明. 不同类型茶园昆虫、蜘蛛群落结构分析. 生态学报, 1998, 18(3): 289–294. |
| [19] | 姚琳, 钟金, 毛建辉, 陈宇, 王胜华. 不同类型茶园昆虫和蜘蛛的总群落排序及分类. 贵州农业科学, 2014, 42(8): 99-101–105-105. |
| [20] | 刘文惠, 洪波, 胡懿君, 贺达汉. 不同景观结构下麦田地面甲虫和蜘蛛物种多样性及优势种分布的时空动态. 应用昆虫学报, 2014, 51(5): 1299–1309. DOI:10.7679/j.issn.2095-1353.2014.154 |
| [21] | Berger W H, Parker F L. Diversity of planktonic foraminifera in deep-sea sediments. Science, 1970, 168(3937): 1345–1347. DOI:10.1126/science.168.3937.1345 |
| [22] | 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ:α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231–239. |
| [23] | Tsang W, Liang G W, Yan H M. Comparison of structure and dynamics of spider community in organic, abandoned and conventional litchi orchards. Journal of Environmental Entomology, 2010, 32(2): 173–179. |
| [24] | Bray J R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 1957, 27(4): 325–349. DOI:10.2307/1942268 |
| [25] | Clarke K R. Non-parametric multivariate analyses of changes in community structure. Austral Ecology, 1993, 18(1): 117–143. DOI:10.1111/aec.1993.18.issue-1 |
| [26] | McCune B, Grace J B. Analysis of Ecological Communities. Oregon:MJM Software Design, 2002. |
| [27] | Churchill T B. Spiders as ecological indicators:an overview for Australia. Memoirs of the Museum of Victoria, 1997, 56(2): 331–337. |
| [28] | Wheater C P, Cullen W R, Bell J R. Spider communities as tools in monitoring reclaimed limestone quarry landforms. Landscape Ecology, 2000, 15(5): 401–406. DOI:10.1023/A:1008171023039 |
| [29] | 张欣颖. 北京农林复合系统蜘蛛群落结构及其与环境关系研究[D]. 北京: 北京林业大学, 2012: 1-73. |
| [30] | 黎健龙, 唐劲驰, 赵超艺, 唐颢, 黎秀娣, 黎华寿. 不同景观斑块结构对茶园节肢动物多样性的影响. 应用生态学报, 2013, 24(5): 1305–1312. |
| [31] | 马艳滟, 李巧, 冯萍, 杨自忠. 云南苍山火烧迹地不同恢复期地表蜘蛛群落多样性. 生态学报, 2013, 33(3): 964–974. |
| [32] | 蒋杰贤, 万年峰, 季香云, 淡家贵. 桃园生草对桃树节肢动物群落多样性与稳定性的影响. 应用生态学报, 2011, 22(9): 2303–2308. |
| [33] | 何昌彤, 郑国, 李学军. 岫岩地区大豆田地表蜘蛛边缘效应及优势种群动态分析. 四川动物, 2015, 34(3): 364–369. |
| [34] | 黎健龙, 唐劲驰, 黎秀娣, 唐颢, 黎华寿. 周边不同生境条件对茶园蜘蛛群落及叶蝉种群时空结构的影响. 生态学报, 2014, 34(9): 2216–2227. |
| [35] | 刘晓博. 黄河下游典型农区景观结构及生境特征对表生蜘蛛群落的影响[D]. 郑州: 河南大学, 2015: 1-35. |
| [36] | 钟金. 不同类型茶园害虫天敌及其总群落研究[D]. 成都: 四川农业大学, 2012. |
| [37] | Uetz G W. Habitat structure and spider foraging//Bell S S, McCoy E D, Mushinsky H R, eds. Habitat Structure:The Physical Arrangement of Objects in Space. Netherlands:Springer, 1991:325-348. |
| [38] | Buchholz S. Ground spider assemblages as indicators for habitat structure in inland sand ecosystems. Biodiversity and Conservation, 2010, 19(9): 2565–2595. DOI:10.1007/s10531-010-9860-7 |
| [39] | 邢树文, 朱慧. 潮州凤凰山茶园蜘蛛优势度与多样性分析. 广东农业科学, 2015, 42(21): 81–90. DOI:10.3969/j.issn.1004-874X.2015.21.016 |
| [40] | 邢树文, 朱慧, 马瑞君, 吴清韩, 查广才. 有机茶园蜘蛛群落动态分析. 韩山师范学院学报, 2016, 37(3): 59–67. |