 2017, Vol. 37
2017, Vol. 37文章信息
- 薛文艳, 杨斌, 张文辉, 于世川
- Xue W Y, Yang B, Zhang W H, Yu S C.
- 桥山林区麻栎种群不同发育阶段空间格局及关联性
- Spatial pattern and spatial association of Quercus acutissima at different developmental stages in the Qiaoshan Mountains
- 生态学报. 2017, 37(10): 3375-3384
- Acta Ecologica Sinica. 2017, 37(10): 3375-3384
- http://dx.doi.org/10.5846/stxb201603130447
-
文章历史
- 收稿日期: 2016-03-13
- 网络出版日期: 2017-02-17

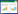




种群空间分布格局是指种群内个体在生存空间的分配方式及配置特点, 是种群与环境因素相互作用的结果[1]。分析种群空间分布格局有助于了解种群生长发育、生活史规律, 种子传播、种内种间关系, 以及种群与环境因素的互作效应[2-4]。对不同发育阶段种群格局类型及其关联性的研究可以更好的揭示种群发育过程中对环境资源的适应和利用状况[3]。
研究空间格局可有多种方法, 其中以点格局分析应用最为广泛[5-6]。该方法以个体在二维空间的坐标为基础, 构成点图, 并以此点图为基础进行分析。它基于点之间的距离统计, 最大限度的利用了点之间的距离信息, 可提供更全面的空间格局信息[7]。点格局分析可分析各尺度种群空间分布格局, 同时还可分析种群间关联性, 并给出空间分布格局的最大聚集强度及其对应的尺度[8]。由于空间格局具有很强的尺度依赖性, 即空间格局的分布特性会随尺度的不同而发生变化, 因此, 点格局分析的结果更符合生态学规律。
麻栎Quercus acutissima, 壳斗科栎属, 是黄土高原南部桥山地区阔叶落叶林地带性植被的建群种, 常以纯林或松栎混交林形式构成当地最重要森林景观, 在当地涵养水源、保持水土、保护生物多样性方面发挥着重要作用[9]。目前对栎属植物空间格局的研究主要集中于栓皮栎[10]、辽东栎[11]、锐齿栎[12]。袁志良[10]等研究发现栓皮栎种群空间格局受空间异质性的影响并影响种群的天然更新;常伟等[12]认为不同生境锐齿栎空间分布格局不同, 营造适宜生境可促进种群更新与稳定。迄今为止, 有关麻栎种群空间分布及空间关联鲜有报道。本研究以桥山林区麻栎天然次生纯林和油松麻栎天然次生混交林为研究对象, 用点格局分析法分析了不同发育阶段麻栎种群空间分布格局及空间关联, 阐明了种群分布格局与发育阶段的关系及不同发育阶段的关联性, 揭示了种群发育动态变化机制, 为种群合理经营及持续稳定更新提供依据。
1 研究区概况研究区选择在陕北黄土高原延安市桥山林业局双龙林场, 地理位置为108°31′—109°11′ E, 35°30′—35°50′ N, 海拔800—1700 m, 属暖温带气候区, 年平均气温9.4 ℃, 最高气温38 ℃, 最低气温-23 ℃, 无霜期190—225 d, 年均降水量677.4 mm, 主要土壤类型为森林褐土和灰褐土。主要乔木树种有辽东栎(Quercus wutaishanica)、麻栎(Quercus acutissima)、油松(Pinus tabuliformis)、山杨(Populus davidiana)和白桦(Betula platyphylla)等;主要灌木树种有胡枝子(Lespedeza bicolor)、狼牙刺(Sophora davidii)、刺五加(Eleutherococcus senticosus)、水栒子(Cotoneaster multiflorus)等;主要草本植物有白茅(Imperata cylindrica)、茜草(Rubia cordifolia)等[9]。
2 研究方法 2.1 样地设置与调查为了准确揭示麻栎种群不同发育阶段的空间格局及其关联性, 于2015年7—8月经过野外踏查后, 根据点格局研究的取样要求[7], 在双龙林场南沟, 选择一个较大的阴坡坡面由一条较大侵蚀沟分割, 具有典型特征的林地:林地A为麻栎纯林, 林地B为油松-麻栎混交林。根据林场保留的林相图和小班卡记录, 两块样地1976年前作为薪炭林, 林场和当地农户从中经常性从中采樵生活用柴, 形成比较残败的天然次生林(麻栎纯林, 油松—麻栎混交林), 1976年实行封育措施, 此后随着人口迁出, 林场职工减少, 两块林地未经受过较大的外界干扰。在A、B两块林地分别设置面积为50 m×50 m两块样地(A、B样地)。采用相邻网格法, 分别将两块样地划分为25个10 m×10 m的样方;以样地左下方为原点, 对样方内的麻栎个体进行坐标定位;每木调查包括植株的高度、胸径、冠幅;对于所有高度小于1 m的植株, 测定其基径、高度、冠幅;记录样地海拔、坡向、坡度等环境因子, 并用样点统计法调查林分郁闭度[13], 在每块样地内随机设置50个样点, 观察树冠是否遮盖样点, 统计被遮盖的样点数, 进而得到林分郁闭度, 样地基本情况见表 1。
| 样地 Plot | 海拔/m Elevation | 坡度/° Slope | 坡向 Aspect | 平均胸径/cm Mean DBH | 平均树高/m Mean tree height | 郁闭度 Canopy density |
| A | 1274 | 20 | ES10° | 14.8 | 13.8 | 0.7 |
| B | 1266 | 25 | ES25° | 18.8 | 7.5 | 0.65 |
| A:麻栎纯林, Q.acutissima natural secondary pure forest;B:麻栎-油松混交林, Q.acutissima-Pinus tabuliformis natural secondary mixed forest | ||||||
不同发育阶段种群的数量结构在一定的程度上可以反映种群的发展趋势[14]。栎属树木通常树体高大且木质坚硬, 不易用生长锥直接测定其年龄, 通常以径级结构和高度级结构代替年龄结构进行分析[15-16]。本次调查径级划分标准为[17]:Ⅰ级(高度H ≤ 50 cm)、Ⅱ级(50 cm<H ≤ 100 cm)、Ⅲ级(H>100 cm, DBH<4 cm)、Ⅳ级(4 cm ≤DBH<8 cm)、Ⅴ级(8 cm ≤ DBH<12 cm)、Ⅵ级(12 cm ≤ DBH<16 cm)、Ⅶ级(16 cm ≤ DBH<20 cm)、Ⅷ级(20 cm ≤ DBH<24 cm)、Ⅸ级(24 cm ≤ DBH<28 cm)和Ⅹ级(DBH ≥ 28 cm)。依据实际调查结果并结合麻栎生长特性, 将麻栎种群划分为4个生长阶段: s1幼苗(Ⅰ、Ⅱ级)、s2幼树(Ⅲ、Ⅳ级)、s3中龄树(Ⅴ—Ⅷ级)和s4成龄树(Ⅸ—Ⅹ级)。分别统计各阶段株树, 并以其为横坐标, 以各阶段株数为纵坐标, 做麻栎种群不同发育阶段数量结构图(图 1)。

|
| 图 1 桥山双龙林场两种样地麻栎种群不同发育阶段个体数量分布图 Fig. 1 Number of Quercus acutissimain different growth |
Morishita Index(MI)可用来表示种群聚集程度, 一个MI表示固定样方内种群的分布格局。将固定样方连续分割后得到不同的MI, 便可将空间聚集程度与尺度联系在一起。具体方法为将每一个大样方逐步分割为2×2, 3×3, …, n×n个小样方, 通过统计落入样方内的个体数计算一组连续的MI[18]。
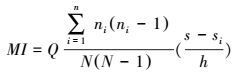
|
式中, ni代表落入第i个样方中的个体数, N代表所有点的计数和。完全随机分布的种群, MI接近于1, MI值大于1则代表种群呈聚集分布。以样方大小为横坐标, MI值为纵坐标绘制麻栎种群不同尺度下的聚集指数图(图 2)。
2.4 点格局分析点格局分析法(point pattern analysis)由Ripley最先提出[19], 其主要思路为通过划定一定面积的研究区域, 以该区域内植物个体的空间坐标构成的点图为基础数据进行格局分析。由于其可用于分析任一尺度下种群的空间分布格局, 因此在实际中得到了广泛的应用。Ripley′s K(r)函数公式为:
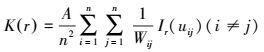
|
式中, A是研究区(样方)的面积;n为个体数;uij为两个点i和j之间的距离;Ir(uij)为指示函数, 当uij<r时, Ir(uij)=1, 当uij>r时, Ir(uij)=0;wij为权重值, 用于边缘校正。
通常情况下, 为保持方差稳定, 通常用L(r)代替K(r)函数, 其表达式为:
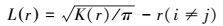
|
当L(r)=0时, 表示随机分布, 位于上下包迹线之间;当L(r)>0时, 为集群分布, 位于上下包迹线之上;当L(r)<0时, 为均匀分布, 位于上下包迹线之下。
不同生长阶段麻栎个体间空间关系可以用双变量函数分析, 具体计算公式为:
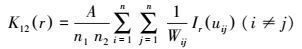
|
同理, 可用L12(r)代替K12(r), 表达式为:
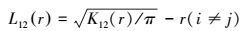
|
当L12(r)=0时, 表示两变量间相互独立, 位于上下包迹线之间;当L12(r)>0时, 表示两变量空间正相关, 位于上下包迹线之上;当L12(r)<0时, 表示两变量间呈空间负相关, 位于上下包迹线之下。
本文通过Monte-Carlo拟合检验确定置信区间, 选取空间完全随机模型作为零模型[20], 重复拟合99次, 得到95%置信区间, 即图中上、下包迹线之间的范围。以r为横坐标, 上下包迹线为纵坐标作图。本文所有数据处理、格局分析、统计图制作均通过R软件spatstat包[21]实现。
3 结果与分析 3.1 种群不同发育阶段数量分布图由图 1可知, 样地A中, 各阶段个体数分别占个体总数的76.7%、5.4%、15.9%、2.0%。样地B中, 各阶段个体数分别占个体总数的89.4%、2.1%、6.6%、1.9%。两块样地中, 麻栎种群不同发育阶段数量结构均呈基部宽顶部窄的不规则倒“J”型。s1阶段的麻栎个体数最多, 说明种群更新状态良好。尽管幼苗数量储备丰富, 但幼树数量不足, 样地A中, 由s1阶段到s2阶段, 种群个体数减少92.9%, 样地B相同阶段种群个体数减少97.6%, 在此阶段竞争激烈导致只有少数幼苗能顺利进入s2, s2阶段成为制约种群发育的瓶颈期。在瓶颈期之后两块样地麻栎个体数分别增加192%, 209%。在s4阶段也出现了一个死亡高峰, 但死亡率低于s2阶段。
3.2 麻栎种群总体分布格局由图 2可知, 0—35 m范围内, 样地A和样地B麻栎种群MI始终大于1, 两麻栎种群均表现为聚集分布。样地A中, 种群MI值在0—10 m尺度内迅速减小, 10—35 m尺度上MI值稳定在1左右, 说明样地A中麻栎种群在10—35 m尺度上表现为低强度聚集分布;样地B中, 0—5 m尺度上种群MI值迅速降低, 在5—35 m尺度上表现为低强度聚集分布, 种群MI值稳定在1左右。

|
| 图 2 桥山双龙林场两种样地麻栎种群总体在不同尺度下的聚集指数MI Fig. 2 Morishita Index of increasing diameter of quadrat |
图 3、图 4表明, s1阶段麻栎种群在0—6 m的小尺度表现为聚集分布, 稍大尺度上表现为随机分布。不同的是, 样地A中, s2阶段种群在0—12 m尺度上均表现为聚集分布;随着种群发育至s3, s4阶段, 分布状态逐渐变为随机分布;样地B中, s2阶段, 种群基本表现为随机分布, 仅在1—2 m尺度上表现为聚集分布;s3阶段, 种群在3 m尺度表现为均匀分布, 12 m尺度上表现为聚集分布;s4阶段各尺度上种群均表现为随机分布。

|
| 图 3 桥山双龙林场样地A麻栎种群各龄级空间分布格局 Fig. 3 Spatial pattern of each stage of Quercus acutissimain plot A s1:幼苗, seedling stage; s2:幼树, sapling stage; s3:中龄树, mid-mature stages4:成龄树, mature stage |

|
| 图 4 桥山双龙林场样地B麻栎种群各龄级空间分布格局 Fig. 4 Spatial pattern of each stage of Quercus acutissimain plot B |
图 5、图 6结果显示, 样地A中, s1与s2阶段的个体仅在1—2 m小尺度上呈负相关, s2与s3阶段的个体在8 m尺度呈正相关, 其余各生长阶段之间在任意尺度上均无关联性。样地B中, s2—s3阶段个体在2—12 m尺度上表现为正相关, 其余各生长阶段之间在各尺度上均无关联。

|
| 图 5 桥山双龙林场样地A麻栎种群各生长阶段空间关系图 Fig. 5 Associations of different growth of Quercus acutissimain plot A |

|
| 图 6 桥山双龙林场样地B麻栎种群各生长阶段空间关系图 Fig. 6 Associations of different growth of Quercus acutissimain plot B |
植物种群的年龄结构是指种群个体的年龄组配状况, 是植物种群生殖特性与外界环境相互作用的结果, 它可以反映种群的动态变化和群落的发展趋势[22]。本研究中, 麻栎种群的龄级结构是幼苗比例最大, 中龄树、成龄树比例次之, 幼树比例最小。这说明该种群属进展性种群, 种群能进行自我更新, 且具有较强的持续发育能力。麻栎幼苗数量多主要是由其生殖特性决定, 麻栎种群可以产生大量高质量种子并萌发[23]。随着种群发育, 幼树阶段出现明显的瓶颈现象, 产生这种现象的原因是, 随着树龄的增大, 麻栎种群不同个体对光照、水分、养分等环境资源需求随之增大, 由于环境资源有限, 种内竞争加剧, 大量的幼苗在竞争中会死亡, 导致尽管麻栎种群幼苗充足, 却难以转化为中高龄个体[24]。林下经常出现只见幼苗不见幼树的现象。随着种群进一步发育, 中龄树对环境的适应能力增强且中龄树占据林冠中上层空间, 光照充足, 竞争力提高, 使得种群死亡率降低, 中龄树个体数增加[25]。成龄树死亡率又有所提高, 这主要是由于此阶段个体进入生理衰老年龄, 出现自然死亡。总的来说, 桥山地区麻栎种群表现出间歇型种群结构, 但由于麻栎种群可通过群落微结构-林窗完成更新[26], 因此这种间歇型结构可保持相对稳定, 从而促进群落的稳定。
麻栎种群分布格局一方面受生殖特性影响, 另一方面也与水分、气候、光照、土壤等可利用生态资源有关[27]。本研究中, 麻栎种群总体上呈聚集分布。这是因为聚集分布可以提高种群竞争力及对外界不良环境的抵抗能力[28]。不同的是, 混交林样地乔木层有油松、白桦、茶条槭, 灌木层有胡枝子、狼牙刺等。在白桦枯死木周围有小面积林窗, 麻栎便在这些林窗聚集。随取样尺度增大, 林窗效应减弱, 气候, 土壤水分与养分状况, 群落郁闭度等影响增强, 导致混交林样地种群聚集程度随尺度迅速变化。而纯林样地乔木层以麻栎为主, 在一定尺度内生境条件相对均匀, 因此样地内种群聚集程度变化缓慢。
对种群在不同发育阶段的空间格局变化的研究可以了解种群幼龄个体的产生及老龄个体的死亡等发育过程并推断种群是否遭受人为干扰。本研究选取的两块样地中, 幼苗阶段个体呈聚集分布, 这主要是由麻栎的生殖特性及种子散布方式决定。同时, 聚集分布使个体间能相互庇护, 进而提高了种群的抗性并降低了死亡率。中龄、成龄阶段个体大致呈随机分布, 这一方面是由于麻栎幼苗耐荫而小树, 大树喜光, 另一方面是由于随着树龄增大, 种内及种间竞争加剧, 自疏及他疏作用加强, 种群密度降低是造成聚集强度降低的主要因素。幼树阶段混交林样地内个体呈随机分布而纯林样地内个体呈聚集分布, 是由于混交林样地内有枯树形成的林窗, 个体在林窗内环境资源充足, 竞争力提高, 因此趋于随机分布。本研究与其他天然群落格局分析结果类似[2, 29]。这是因为自然状态下, 不同发育阶段的个体占据不同的空间位置, 可以使个体对有限环境资源的利用达到最大化, 同时还可以提高种群对环境的适应能力, 有利于其生存发展。由聚集分布到随机分布的变化, 是种群发育的一种生存策略和适应机制[30]。
植物种群的空间分布格局有很强的尺度依赖性。同一物种相同发育阶段在不同尺度上表现出的分布状态均有差异[31]。种群在某一尺度上的空间格局是由多种生态过程共同形成的。尺度较大时, 格局由物种分布、斑块异质性以及环境因素共同决定;尺度较小时, 格局与种内种间竞争、种子落地及扩散方式有关[32]。本研究中, 两块样地中, 幼龄个体在小尺度上呈聚集分布, 这主要是由落种及散布方式决定。随着尺度增大则趋于随机分布, 这种不同发育阶段空间格局的镶嵌分布有利于麻栎种群的稳定发育[33]。
同一物种不同发育阶段在空间分布上的相互关联性可用双变量种间关联来表示, 其指示了某一种群不同发育时期个体间的相互关系, 这种关系既指个体空间分布关系, 也指个体间的功能关系, 可以据此了解种群的现状并预测种群发展趋势[6]。由于树木个体之间的相互影响仅出现在一定范围内[34], 因此对物种的空间分布关联的研究多集中在较小尺度。本研究调查的两块样地内, 幼苗与其它阶段个体在所有尺度上均为负关联或无关联, 这是由于其他个体营造的郁闭环境不利于幼苗生长;幼树与中龄树均在一定尺度上呈正相关, 而与其它阶段个体呈负相关或无关联, 这种现象可能是由于种内竞争具有非对称性[35], 即当个体大小相近时, 二者间关系较协调;当个体间差异较大时, 由于大株个体对小株个体强烈的排斥作用及大株个体自身较强的竞争力, 使其周围小株个体很少[36]。因此中龄树与成龄树在所有尺度上均呈现无关联或负相关。
鉴于麻栎种群现状, 建议有针对性地该地区麻栎天然林进行抚育。林下灌木和草本会影响种子萌发及幼苗存活, 导致幼树阶段成为制约种群发育的瓶颈期。因此应加强下层抚育, 清除部分杂草及灌木, 为幼苗生长创造适宜环境。幼树阶段后对光照要求增加, 此时应对林冠层进行透光伐, 加快个体生长发育, 促进种群良性发育。
| [1] | 薛建辉. 森林生态学. 北京: 中国林业出版社, 2006: 75. |
| [2] | Druckenbrod D L, Shugart H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont. Journal of Vegetation Science, 2005, 16(1): 37–48. DOI:10.1111/j.1654-1103.2005.tb02336.x |
| [3] | Nathan R. Long-distance dispersal of plants. Science, 2006, 313(5788): 786–788. DOI:10.1126/science.1124975 |
| [4] | 胡尔查, 王晓江, 张文军, 海龙, 张雷, 张胜利, 徐鹏雁. 乌拉山自然保护区白桦种群的年龄结构和点格局分析. 生态学报, 2013, 33(9): 2867–2876. |
| [5] | Zhang J C, Zhang Y Q, Fan D Q, Qin S G, Jia X, Wu B, Chen D, Gao H, Zhu L F. Effects of sand dune stabilization on the spatial pattern of Artemisia ordosicapopulation in Mu Us desert, Northwest China. PLoS One, 2015, 10(6): e0129728. DOI:10.1371/journal.pone.0129728 |
| [6] | Shields J M, Jenkins M A, Saunders M R, Zhang H, Jenkins L H, Parks A M. Age distribution and spatial patterning of an invasive shrub in secondary hardwood forests. Forest Science, 2014, 60(5): 830–840. DOI:10.5849/forsci.13-079 |
| [7] | 张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344–349. |
| [8] | Kaito C, Dieckmann U, Sasaki A, Takasu F. Beyond pairs: definition and interpretation of third-order structure in spatial point patterns. Journal of Theoretical Biology, 2015, 372(6): 22–38. |
| [9] | 曹旭平, 张文辉. 陕西桥山省级自然保护区综合科学考察报告. 杨凌:西北农林科技大学出版社, 2015: 1–5. |
| [10] | 袁志良, 王婷, 朱学灵, 沙迎迎, 叶永忠. 宝天曼落叶阔叶林样地栓皮栎种群空间格局. 生物多样性, 2011, 19(2): 224–231. |
| [11] | 王道亮, 李卫忠, 曹铸, 王庆, 王广儒. 黄龙山天然次生林辽东栎种群空间格局. 生态学报, 2016, 36(9): 1–10. |
| [12] | 常伟, 党坤良, 武朋辉, 李明雨. 秦岭林区锐齿栎次生林种群空间分布格局. 生态学报, 2016, 36(4): 1021–1029. |
| [13] | RautiainenM, StenbergP, NilsonT. Estimating canopy cover in Scots pine stands. Silva Fennica, 2005, 39(1): 137–142. |
| [14] | Lan G Y, Getzin S, Wiegand T, Hu Y H, Xie G S, Zhu H, Cao M. Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China. PLoS One, 2010, 7(11): e46074. |
| [15] | 蔡飞. 杭州西湖山区青冈种群结构和动态的研究. 林业科学, 2000, 36(3): 67–72. |
| [16] | Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed treeQuercusrobur in two spruce-dominated forests. Écoscience, 2000, 7(1): 38–44. DOI:10.1080/11956860.2000.11682569 |
| [17] | 周建云, 李荣, 张文辉, 何景峰. 不同间伐强度下辽东栎种群结构特征与空间分布格局. 林业科学, 2012, 48(4): 149–155. |
| [18] | 王晓雨, 于大炮, 周莉, 周旺明, 吴志军, 郭焱, 包也, 孟莹莹, 代力民. 长白山北坡林线岳桦种群空间分布格局. 生态学报, 2015, 35(1): 116–124. |
| [19] | Ripley B D. Modelling spatial patterns. Journal of the Royal statistical Society.Series B, 1977, 39(2): 172–212. |
| [20] | Wang X T, Liang C Z, Wang W. Balance between facilitation and competition determines spatial patterns in a plant population. Chinese Science Bulletin, 2014, 59(13): 1405–1415. DOI:10.1007/s11434-014-0142-8 |
| [21] | BaddeleyA, Turner B. Spatstat: an R package for analyzing spatial point patterns. Journal of Statistical Software, 2005, 12(6): 1–42. |
| [22] | Benson E J, Hartnett D C. The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie. Plant Ecology, 2006, 187(2): 163–178. DOI:10.1007/s11258-005-0975-y |
| [23] | Wang XG, Ye J, Li BH, Zhang J, Lin F, Hao ZQ. Spatial distributions of species in an old-growth temperate forest, northeastern China. Canadian Journal of Forest Research, 2010, 40(6): 1011–1019. DOI:10.1139/X10-056 |
| [24] | 蔡新斌, 吴俊侠. 甘家湖自然保护区白梭梭种群特征与动态分析. 干旱区资源与环境, 2016, 30(7): 90–94. |
| [25] | 闫兴富, 杜茜, 石淳, 周立彪, 张靠稳. 六盘山区辽东栎的实生苗更新及其影响因子. 植物生态学报, 2011, 35(9): 914–925. |
| [26] | 赵学农, 刘伦辉, 高圣义, 和爱军. 版纳青梅种群结构动态与分布格局. 植物学报, 1993, 35(7): 552–560. |
| [27] | Li X M, Zhou W Q, Ouyang Z Y. Relationship between land surface temperature and spatial pattern of greenspace: what are the effects of spatial resolution?. Landscape and Urban Planning, 2013, 114(8): 1–8. |
| [28] | 龙成, 杨小波, 龙文兴, 李东海. 铜鼓岭热带常绿季雨矮林5种蒲桃属植物的种群结构及空间格局. 林业科学, 2015, 51(2): 18–27. |
| [29] | 刘振, 李红丽, 董智, 李钢铁, 万玲玲, 岳永杰. 浑善达克沙地2种生境下榆树种群空间点格局. 林业科学, 2012, 48(1): 29–34. |
| [30] | Hubbell S P. Neutral theory and the evolution of ecological equivalence. Ecology, 2006, 87(6): 1387–1398. DOI:10.1890/0012-9658(2006)87[1387:NTATEO]2.0.CO;2 |
| [31] | Greig-Smith P. Quantitative Plant Ecology.3rd ed. Oxford: Blackwell Scientific Publications, 1983. |
| [32] | Wiegand T, Gunatilleke S, Gunatilleke N, Okuda T. Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering. Ecology, 2007, 88(12): 3088–3102. DOI:10.1890/06-1350.1 |
| [33] | 张希彪, 郭小强, 上官周平, 刘富顺. 黄土丘陵子午岭油松天然林群落特征研究. 植物研究, 2006, 26(2): 169–175. |
| [34] | 侯继华, 黄建辉, 马克平. 东灵山辽东栎林主要树种种群11年动态变化. 植物生态学报, 2004, 28(5): 609–615. |
| [35] | 尤海舟, 刘兴良, 缪宁, 何飞, 马钦彦. 川滇高山栎种群不同海拔空间格局的尺度效应及个体间空间关联. 生态学报, 2010, 30(15): 4004–4011. |
| [36] | Kenkel N C. Pattern of self-thinning in Jack Pine: testing the random mortality hypothesis. Ecology, 1988, 69(4): 1017–1024. DOI:10.2307/1941257 |