 2017, Vol. 37
2017, Vol. 37文章信息
- 刘文亭, 卫智军, 吕世杰, 王天乐, 张爽
- LIU Wenting, WEI Zhijun, LÜ Shijie, WANG Tianle, ZHANG Shuang.
- 放牧对短花针茅荒漠草原植物多样性的影响
- The impacts of grazing on plant diversity in Stipa breviflora desert grassland
- 生态学报. 2017, 37(10): 3394-3402
- Acta Ecologica Sinica. 2017, 37(10): 3394-3402
- http://dx.doi.org/10.5846/stxb201603120435
-
文章历史
- 收稿日期: 2016-03-12
- 网络出版日期: 2017-02-17





2. 内蒙古农业大学理学院, 呼和浩特 010018
2. Inner Mongolia Agricultural University, College of Science, Huhhot 010018, China
生物多样性是当前生态学领域的重大科学问题[1-2]。以往的生物多样性、生态系统功能及维持机制的研究中, 草地生态系统以其巨大的土地面积、过频的人类干扰、较强的实验操控性赢得了研究人员的重视[3-4]。家畜通过采食等 (下行作用, Top-down) 作用植株个体生存来操控天然草地植物多样性[5], 因此, 揭示放牧对草地生态系统植物多样性的影响过程, 对于草地生态系统的多样性保育和可持续利用具有重要的意义。
生态学现象均不同程度地表现出尺度依赖性, 即在相应的空间尺度、时间尺度或组织尺度背景下来开展相应研究[6]。物种多样性是基于低组织尺度的数量特征来获得高组织尺度研究范畴的运算方法, 表征的是低组织尺度的植物多样性, 但却是高组织尺度的概念, 即通常研究的物种多样性体现的是群落这一尺度的概念。物种多样性与功能群成员多样性尽管是基于物种的数量特征计算, 但前者体现的是群落的概念, 后者是功能群的概念[7]。这亦提供了一个思路, 在研究植物多样性时, 可通过扩张研究尺度来俯瞰植物多样性, 亦可缩小研究尺度将局部细节展漏无疑, 这将更有利于理解和简化生态系统过程[8]。
放牧家畜是草地群落组成的直接管理者, 研究认为, 高植物多样性条件下, 家畜拥有更多的机会去选择其喜食的物种或物种组合[9], 频繁地在不同物种间进行尝试, 而家畜个体的数量增加会增大其选择的积累效应, 导致草地多样性差异。模拟实验显示, 物种或功能群去除促进了保留种的生长, 功能群间的竞争关系通过保留种或其相应功能群的补偿生长进而稳定[10]。而群落结构简单, 层次分明, 物种较少并以强旱生多年生丛生小禾草为主导的短花针茅荒漠草原, 植物多样性对放牧干扰如何响应?本研究依托于内蒙古锡林郭勒盟苏尼特右旗的短花针茅荒漠草原建立的长期放牧平台作为模式生态系统, 从植物组织尺度入手, 以期回答以下问题:1) 不同放牧强度的草地群落的物种组成如何变化, 是否存在积极响应的物种或功能群?2) 在群落上, 植物物种多样性对放牧强度的响应有何不同?进一步其每个功能群成员 (多年生禾草、灌木、半灌木、多年生杂类草和一年生草本) 对放牧强度的响应又有何差异, 是否与群落尺度的多样性变化相一致?揭示荒漠植物组成、多样性及稳定性对放牧的响应, 对制定干旱区荒漠草原的放牧管理制度提供数据支撑与理论基础。
1 材料与方法 1.1 研究区概况本研究在内蒙古锡林郭勒盟苏尼特右旗朱日和镇 (112°47′11.2″E, 42°15′48.7″N) 的牧区进行。该地区地形平坦, 有明显的钙积层, 主要分布在10—30cm。土壤为淡栗钙土, 腐殖质层厚5—10cm。2012—2014年平均降水量230.9mm, 而60%—80%的降水集中在牧草生长旺季的7月到9月, 蒸发量2500mm, 大多数年受到不同程度的干旱威胁。2012—2014年月平均温度较为接近, 年平均气温5.9℃, 月平均最高温度24.45℃, 最低温度-16.28℃。优势种为短花针茅 (Stipa breviflora)、无芒隐子草 (Cleistogenes songorica)、碱韭 (Allium polyrhizum), 伴生寸草苔 (Carex duriuscula)、狭叶锦鸡儿 (Caragana stenophylla)、戈壁天门冬 (Asparagus gobicus)、木地肤 (Kochia prostrata)、银灰旋花 (Convolvulus ammannii)、茵陈蒿 (Artemisia capillaries)、阿尔泰狗娃花 (Heteropappus altaicus)、乳白花黄芪 (Astragalus gulactites) 等。
1.2 试验地设置与取样方法放牧试验从2010年开始, 每年5月开始放牧, 10月底放牧终止, 期间采用连续放牧方式, 晚上羊群在样地不归牧, 不进行补饲。试验平台设有放牧季节调控、放牧强度调控与放牧强度下季节调控3种实验方案, 而本研究试图从放牧强度这一角度来解释植物多样性对放牧的响应, 故抽取适度放牧 (MG)、重度放牧 (HG)、不放牧 (NG) 这3种处理方式 (图 1)。每个处理均设有3次重复, 共9个试验小区。每个实验小区面积约2.60hm2。适度放牧区和重度放牧区分别放牧健康的大小、体重、性别基本一致的苏尼特羊5和8只, 载畜率为0.96只羊hm-2 a-1和1.54只羊hm-2 a-1。每年植物生长高峰期 (8月) 进行取样, 每个试验小区随机调查10个1m×1m样方。每个样方主要调查指标为物种数、密度 (丛生型植物为分株数, 非丛生型禾草植物为个体数)。每年共调查90个样方, 本研究的数据取自2012—2014年, 共270个样方。

|
| 图 1 放牧试验处理设计图 Fig. 1 Design diagram of grazing treatment |
以生活型为依据把群落划分为4类[8]:1) 多年生禾草 (PG);2) 灌木、半灌木 (SS);3) 多年生杂类草 (PF);4) 一年生草本 (AB)。
1.4 数据处理 1.4.1 多样性指标的计算科学测定区域生境内的多样性时, 通常以群落内物种数及其相对多度为基础, 来反映生境内物种间长期竞争而形成的多物种共存关系。丰富度指数体现了群落内物种数量, 但忽略了优势种和稀有种对多样性贡献大小的差异, 且易受野外取样面积的影响, 在应用时需与Simpson生态优势度指数、Shannon-Wiener多样性指数、Pielous均匀度指数等结合起来才更为准确。Shannon-Wiener多样性指数是群落内物种多样性和异质性程度的参数, 综合反映群落物种丰富度和均匀度, 但却忽略了物种组成因素。Simpson生态优势度指数所表达的信息与多样性指数和均匀度指数相反, 反映了各物种数量的变化情况, 其数值越大, 说明群落内物种数量分布越不均匀, 优势种的地位越突出。Pielous均匀度指数则体现了各群落内物种个体数分布的均匀程度。具体公式如下:
物种丰富度
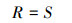
|
Simpson生态优势度指数

|
Shannon-Wiener多样性指数
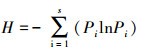
|
Pielous均匀度指数
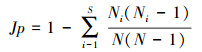
|
式中, S代表物种数, N代表所有物种的多度, Ni代表第i个物种的多度, Pi=Ni/N。
1.4.2 群落稳定性指数采用Godron稳定性测定方法评价群落的稳定性[11]。以试验小区所有样方为单位, 将群落中不同植物种的频度由大到小排序, 把植物的频度换算成相对频度, 按相对频度由大到小的顺序逐步累积相加, 与此同时, 逐一对应累积植物总物种数倒数, 采用模拟散点所得平滑曲线的方程与坐标轴第一象限的曲线y=1-x连接的对角直线来判断累积植物总物种数倒数与累积相对频度比值的交点坐标, 0.2/0.8是群落的稳定点, 交点坐标距离越接近0.2/0.8, 群落就越稳定。
1.5 统计分析数据统计分析过程在SAS 9.0和Excel 2007中完成。对植物群落、功能群密度、频度进行数据整理, 并计算最大值、上四分位数、中位数、下四分位数、最小值来表征数据位置和分散情况的信息, 并分析群落中各功能群所的比例, 用变异系数 (CV) 表征相同放牧处理内功能群多样性的变异特征, 计算方法为标准差除以平均值。对不同放牧强度下群落内物种与群落内各功能群的丰富度指数、Simpson生态优势度指数、Shannon-Wiener多样性指数、Pielous均匀度指数进行单因素方差分析, 并进行Duncun多重比较, 数据以平均值±标准误差表示。
2 结果和分析 2.1 物种组成与功能群对放牧的响应野外调查数据显示 (表 1), 短花针茅荒漠草原植物共有10科24种, 禾草科 (16.67%)、百合科 (20.83%)、菊科 (20.84%) 所占比例大于其他科植物的总和, 之后依次为豆科和藜科。对不同放牧强度的比较发现, 未放牧的草地物种组成 (23种) 最丰富, 适度放牧的草地次之 (16种), 而适度放牧和重度放牧的草地物种组成数基本一致。进一步分析, 物种组成差异主要由兔唇花、棘豆、叉枝鸦葱、冷蒿、二裂委陵菜、牻牛儿苗与画眉草等引起, 且集中于多年生杂类草和一年生草本这2个功能群中。
| 种Species | 未放牧 No grazing |
适度放牧 Moderate grazing |
重度放牧 Heavy grazing |
| 短花针茅Stipa breviflora | 1 | 1 | 1 |
| 寸草苔Carex duriuscula | 1 | 1 | 1 |
| 狭叶锦鸡儿Caragana stenophylla | 1 | 1 | 1 |
| 茵陈蒿Artemisia capillaries | 1 | 1 | 1 |
| 碱韭Allium polyrhizum | 1 | 1 | 1 |
| 银灰旋花Convolvulus ammannii | 1 | 1 | 1 |
| 乳白花黄芪Astragalus gulactites | 1 | 1 | 1 |
| 冷蒿Artemisia frigida | 1 | 0 | 0 |
| 细叶葱Allium tuberosum | 1 | 1 | 1 |
| 二裂委陵菜Potentilla bifurca | 1 | 0 | 0 |
| 猪毛菜Salsola collina | 1 | 1 | 1 |
| 狗尾草Setaria viridis | 1 | 0 | 1 |
| 无芒隐子草Cleistogenes songorica | 1 | 1 | 1 |
| 木地肤Kochia prostrata | 1 | 1 | 1 |
| 天门冬Asparagus gobicus | 1 | 1 | 1 |
| 阿尔泰狗娃花Heteropappus altaicus | 1 | 1 | 1 |
| 兔唇花Lagochilus diacanthophyllus | 1 | 0 | 0 |
| 棘豆Oxytropis | 1 | 0 | 0 |
| 叉枝鸦葱Scorzonera divaricata | 1 | 0 | 0 |
| 蒙古葱Allium mongolicum | 1 | 1 | 0 |
| 细叶韭Allium tenuissimum | 1 | 1 | 0 |
| 栉叶蒿Neopallasia pectinata | 1 | 1 | 1 |
| 牻牛儿苗Erodium stephanianum | 1 | 0 | 0 |
| 画眉草Eragrostis pilosa | 1 | 0 | 0 |
| 1代表有相应物种, 0代表无相应物种 | |||
放牧处理植物功能群统计结果 (表 2) 显示, 仅有未放牧草地的灌木、半灌木的比例显著大于适度放牧和重度放牧的草地 (P < 0.05), 未放牧处理草地的多年生禾草、多年生杂类草和一年生草本的比例均与放牧处理草地无显著性差异;变异系数重度放牧处理最高, 未放牧次之, 适度放牧最低。
| 项目 Item |
多年生禾草 Perennial grasses |
灌木、半灌木 Shrubs and semi-shrubs |
多年生杂类草 Perennial forbs |
一年生草本 Annual forbs |
|||||||
| 比例 Percentage |
CV | 比例 Percentage |
CV | 比例 Percentage |
CV | 比例 Percentage |
CV | ||||
| 未放牧No grazing | 0.34±0.01a | 24.65 | 0.13±0.01a | 78.58 | 0.47±0.02a | 28.33 | 0.06±0.01a | 139.67 | |||
| 适度放牧Moderate grazing | 0.34±0.02a | 30.00 | 0.11±0.01b | 65.21 | 0.47±0.01a | 16.28 | 0.08±0.01a | 103.75 | |||
| 重度放牧Heavy grazing | 0.36±0.02a | 36.57 | 0.08±0.02b | 113.97 | 0.47±0.03a | 36.07 | 0.09±0.02a | 145.17 | |||
在群落上, 如图 2所示, 放牧强度没有显著影响丰富度指数 (以平均值为基础来进行统计), 而未放牧小区丰富度指数最大值与最小值 (11, 4) 均大于重度放牧小区 (9, 3)。Simpson生态优势度指数、Shannon-Wiener物种多样性指数、Pielou均匀度指数的结果规律一致, 未放牧处理显著大于重度放牧处理 (P < 0.05);且同样适用于各多样性指数最大值。而极差方面, 适度放牧小区数值最小。

|
| 图 2 短花针茅荒漠草原植物物种多样性指数 (平均值±标准误差) Fig. 2 Plant species diversity index in Stipa breviflora desert grassland (mean ± SE) 不同大写字母表示存在显著性差异; NG:不放牧No grazing; MG:适度放牧Moderate grazing; HG:重度放牧Heavy grazing |
在功能群上 (图 3), 多年生禾草与一年生草本的各多样性指数对放牧无显著性响应。在丰富度指数方面, 未放牧 (0.93±0.09) 与适度放牧处理 (0.79±0.08) 灌木、半灌木显著大于重度放牧的小区 (0.52±0.08), 其他功能群则对不同的放牧强度无显著性差异。在Simpson生态优势度指数上, 未放牧处理的多年生杂类草显著高于重度放牧小区 (P < 0.05), 而这一现象同样适用于Shannon-Wiener物种多样性指数与Pielou均匀度指数。

|
| 图 3 短花针茅荒漠草原植物功能群多样性指数 (平均值±标准误差) Fig. 3 Plant functional groups diversity index in Stipa breviflora desert grassland (mean ± SE) 图中3种处理从左向右依次未放牧、适度放牧、重度放牧, 每组处理分为4个功能群, 从左向右依次为多年生禾草、灌木半灌木、多年生杂类草、一年生草本; 不同小写字母表示存在显著性差异 |
将短花针茅荒漠草原群落累积相对频度与总物种数的倒数一一对应, 绘制种总数倒数累积-相对频度累积的散点图 (图 4), 之后完成平滑曲线的模拟。适度放牧、重度放牧和未放牧草地群落相应的交点坐标分别为 (0.16, 0.83)、(0.08, 0.91)、(0.12, 0.87)。离群落稳定点 (0.2, 0.8) 距离最近的是适度放牧的草地群落, 其次是未放牧的草地群落, 离群落稳定点最远的是重度放牧群落。

|
| 图 4 短花针茅荒漠草原植物群落Godron散点图 Fig. 4 Godron scatter graphs of plant community in Stipa breviflora desert grassland |
短花针茅荒漠草原不同放牧强度小区植物共有10科24种, 其中禾草科、百合科、菊科植物种类最多。短花针茅、无芒隐子草、寸草苔、木地肤、锦鸡儿、天门冬等均稳定出现于不同放牧强度的草地中 (表 1), 这是因为尽管绵羊个体间存在差异 (如食性选择、行走路线等), 但长期的采食等行为干扰诱使荒漠草原优势种或优势功能群组成与结构发生演变[12], 但亦有部分物种对环境有着极强的适应力, 如多年生丛生禾草的短花针茅、无芒隐子草虽多是矮小的禾草, 组成了荒漠草原独特的以强旱生多年生丛生小禾草为主导的植物群落[13]。说明草原植物群落中, 优势种保持了其较强的竞争优势, 拥有绝对的统治地位。
此外, 木地肤、狭叶锦鸡儿、天门冬、茵陈蒿、阿尔泰狗娃花、碱韭、银灰旋花、乳白花黄芪、细叶葱、栉叶蒿、猪毛菜也出现在各放牧强度处理中 (表 1), 说明这些物种具有耐牧属性。研究认为, 这是家畜在采食时遗留的排泄物或口液等过程诱使植物形成补偿效应[14], 使其仍保持较强的竞争优势, 导致群落中部分物种或由其所组成的功能群 (未放牧处理的多年生禾草、多年生杂类草和一年生草本的权重均与放牧草地无显著性差异) 受放牧影响的结果表现维持不变 (表 2)。一般研究认为, 判断生物多样性高低的重要指标即群落中是否拥有大量的稀有种[15], 这些物种遭受干扰或破坏的过程中, 其相应的生物多样性和生态功能都异常脆弱。相反, 群落中优势种占有大量资源, 直接或间接的控制着其他物种的存活, 拥有绝对的统治地位, 对生态系统功能起主导作用[13]。根据以上论述, 本研究认为尽管稀有种和优势种对植物多样性的研究极其重要, 但在计算现存植物群落的多样性过程中, 草地本身耐牧种的数量特征才是判断其植物多样性高低的决定性因素。
3.2 不同放牧强度对短花针茅荒漠草原植物多样性的影响草原放牧系统是人类生存、生产的重要基础之一, 其可持续生产和有效的生态服务功能关键在于维持该系统的动、植物之间质量方面的平衡, 而生物多样性的关系就是其中的主要反映[15]。在群落尺度上, 未放牧处理多样性指数显著大于重度放牧小区 (P < 0.05), 适度放牧小区大于重度放牧小区 (图 2)。这可能是因为放牧家畜通过改变局部植物种的定植与灭绝速率, 动态调控植物群落的多样性, 当局部物种的灭绝速率高于定植速率, 引起局部物种丰富度的降低[16];也有研究认为, 放牧条件会改变草地物种间的竞争强度, 致使一些在竞争上占劣势的植物种因竞争而被排除, 导致群落中物种多样性降低[17]。但丰富度指数却无显著性差别 (图 2)。这是因为放牧家畜使草地出现较为可观的空余生态位, 增加种库植物可植入的利用位点, 提高了草地植物的繁殖与分布范围, 加之短花针茅荒漠草原本身结构简单, 物种数较少[13], 同时丰富度指数仅考虑调查样方内的物种数量, 并未体现优势种或稀有种对多样性贡献量。
Simpson生态优势度指数、Shannon-Wiener物种多样性指数、Pielou均匀度指数的结果显示, 未放牧草地具有的物种多样性高、均匀程度高和优势种地位高的特点, 适度放牧草地次之, 重度放牧草低最低。这似乎与中度干扰假说不符, 其假说认为, 适度干扰使得草地生境异质性增加, 抑制了原优势种群的竞争能力, 这就为新个体、特别是原有竞争力弱的物种提供了更大生存空间, 新个体还未完全发育便会受到干扰, 优势种群始终不能建立, 致使群落保持较高的多样性[18]。事实上, 家畜采食是一个非常复杂的生态过程, 家畜不仅影响植物多样性, 植物也会形成相应的对策来限制家畜的行为[19], 当某一植物个体与适口性较高的物种共存形成邻居关系时, 动物被更适口的植物吸引, 从而减少了对该物种的采食[20]。而在物种相对较少短花针茅荒漠草原中, 短花针茅与无芒隐子草在群落中占有绝对优势地位, 在其之上, 狭叶锦鸡儿等适口性较差的物种稳定生长, 在其之下, 生长着棘豆、二裂委陵菜、画眉草等低矮植物[13], 绵羊仔觅食过程中优先选择适口性较高的物种, 从而打破原有草地的平衡, 而随着放牧强度增加, 优势种的地位在群落中逐渐降低;此外, 调查群落内适口性差的物种数量的差异也会随着放牧强度的增加被放大, 从而使得Shannon-Wiener物种多样性指数、Pielou均匀度指数降低。
植物功能群是对局地环境有相似表现的一类物种组合[21], 它是基于植物生理生化、形态表现、行为模式、生活史或与其生态系统过程相关的一些生物学特性来划分的。已有研究指出, 家畜的选择性采食仅发生于群落的物种尺度上[22-23], 且选择的强度直接影响群落的植被组合[23]。但对于本研究所在地的群落物种较少、结构简单、功能群成员易区分的短花针茅荒漠草原而言, 家畜采食行为并不需要较高的学习成本, 即随着可利用物种组合数减少, 动物于群落尺度去选择功能群的机会减少, 取而代之的是集中精力去选择每个功能群下的物种。如果某一功能群下的某一物种家畜个体均喜食, 则家畜对功能群下植物多样性所产生累加效应将进一步增大, 这对植物多样性的影响会比群落尺度下物种多样性的作用更强;此外, 本研究将每一个功能群的地位人为的置于所研究的顶端水平, 来试图说明在群落尺度下的每个相对独立的功能群所对应成员的物种多样性, 是群落中物种对不同放牧强度的响应的再次简化过程。这从家畜的角度进一步解释了每个功能群内植物多样性对不同放牧强度的响应机制。
3.3 不同组织尺度植物多样性的意义在生态学研究过程中, 生态学现象或问题都不同程度地表现出尺度依赖性, 即在相应的空间尺度、时间尺度或组织尺度这一大背景下来开展相应的生态学研究。本研究以植物组织尺度为基础, 试图从等级系统来解释放牧强度对短花针茅荒漠草原植物多样性的影响。从群落尺度入手, 分析比较不同放牧强度草地的物种组成与功能群内成员, 在现存物种的基础上, 通过分析不同放牧强度草地多样性指数, 来探讨放牧家畜如何影响天然草地的植物多样性过程, 但事实上, 仅从这一尺度还无法确实局部放大植物多样性与家畜行为的影响过程。接着将研究尺度缩小到功能群上, 意味着将群落内植物多样性的细节放大, 因为每个功能群内物种数的减少或增多与相对多度的变化对多样性指数的变化将更为敏感, 这进一步揭示了放牧家畜对植物多样性造成的影响。
尽管本研究结果显示放牧强度不同程度的影响了短花针茅荒漠草原植物的多样性, 但群落稳定性结果显示出适度放牧草地表现出更高的稳定性。放牧是草地经营管理的重要方式, 因此, 植物多样性与稳定性的权衡将是合理制定科学放牧强度的重要途径。
| [1] | Tilman D, Isbell F. Biodiversity: recovery as nitrogen declines. Nature, 2015, 528(7582): 336–337. DOI:10.1038/nature16320 |
| [2] | Storkey J, MacDonald A J, Poulton P R, Scott T, Köhler I H, Schnyder H, Goulding K W T, Crawley M J. Grassland biodiversity bounces back from long-term nitrogen addition. Nature, 2015, 528(7582): 401–404. DOI:10.1038/nature16444 |
| [3] | Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300–1302. DOI:10.1126/science.277.5330.1300 |
| [4] | Yachi S, Loreau M. Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(4): 1463–1468. DOI:10.1073/pnas.96.4.1463 |
| [5] | Stephens D W, Krebs J R. Foraging Theory. Princeton, New Jersey: Princeton University Press, 1986. |
| [6] | 丁曼, 温仲明, 郑颖. 黄土丘陵区植物功能性状的尺度变化与依赖. 生态学报, 2014, 34(9): 2308–2315. |
| [7] | 刘文亭, 卫智军, 吕世杰, 孙世贤, 贾利娟, 张爽, 王天乐, 代景忠, 卢志宏. 荒漠草地植物多样性对草食动物采食的响应机制. 植物生态学报, 2016, 40(6): 564–573. DOI:10.17521/cjpe.2015.0421 |
| [8] | 刘文亭, 卫智军, 吕世杰, 孙世贤, 代景忠, 闫宝龙. 短花针茅荒漠草原不同组织尺度地上生物量. 应用与环境生物学报, 2015, 21(5): 912–918. |
| [9] | Provenza F D. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. Journal of Range Management, 1995, 48(1): 2–17. DOI:10.2307/4002498 |
| [10] | Wardle D A, Bonner K I, Barker G M, Yeates G W, Nicholson K S, Bardgett R D, Watson R N, Ghani A. Plant removals in perennial grassland: vegetation dynamics, decomposers, soil biodiversity, and ecosystem properties. Ecological Monographs, 1999, 69(4): 535–568. DOI:10.1890/0012-9615(1999)069[0535:PRIPGV]2.0.CO;2 |
| [11] | 郑元润. 森林群落稳定性研究方法初探. 林业科学, 2000, 36(5): 28–32. DOI:10.11707/j.1001-7488.20000513 |
| [12] | 孙世贤, 卫智军, 吕世杰, 卢志宏, 陈立波, 李夏子, 吴艳玲, 李建茹. 放牧强度季节调控下荒漠草原植物群落与功能群特征. 生态学杂志, 2013, 32(10): 2703–2710. |
| [13] | 卫智军, 韩国栋, 赵钢, 李德新. 中国荒漠草原生态系统研究. 北京: 科学出版社, 2013. |
| [14] | 王德利, 王岭. 草食动物与草地植物多样性的互作关系研究进展. 草地学报, 2011, 19(4): 699–704. DOI:10.11733/j.issn.1007-0435.2011.04.028 |
| [15] | Mouillot D, Bellwood D R, Baraloto C, Chave J, Galzin R, Harmelin-Vivien M, Kulbicki M, Lavergne S, Lavorel S, Mouquet N, Mouquet N, Paine C E T, Renaud J, Thuiller W. Rare species support vulnerable functions in high-diversity ecosystems. Plos Biology, 2013, 11(5): 1–15. |
| [16] | Olff H, Ritchie M E. Effects of herbivores on grassland plant diversity. Trends in Ecology & Evolution, 1998, 13(7): 261–265. |
| [17] | 刘晓媛. 放牧方式对草地植被多样性与稳定性关系的影响[D]. 长春: 东北师范大学, 2013. |
| [18] | Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302–1310. DOI:10.1126/science.199.4335.1302 |
| [19] | Wang L, Wang D L, Yu G B, Huang Y, Fan M, Liu J S, Li Y X. Spatially complex neighboring relationships among grassland plant species as an effective mechanism of defense against herbivory. Oecologia, 2010, 164(1): 193–200. DOI:10.1007/s00442-010-1676-3 |
| [20] | Hjältén J, Danell K, Lundberg P. Herbivore avoidance by association: vole and hare utilization of woody plants. Oikos, 1993, 68(1): 125–131. DOI:10.2307/3545317 |
| [21] | 张国钧, 张荣, 周立. 植物功能多样性与功能群研究进展. 生态学报, 2003, 23(7): 1430–1435. |
| [22] | De Vries M F W, Daleboudt C. Foraging strategy of cattle in patchy grassland. Oecologia, 1994, 100(1): 98–106. |
| [23] | Laca E A, Demmen M W. Foraging strategies of grazing animals//Hodgson J, Illius A, eds. The Ecology and Management of Grazing Systems. Wallingford: CAB International, 1996: 37-158. |