 2017, Vol. 37
2017, Vol. 37文章信息
- 卫楠, 张弥, 王辉民, 张雷明, 温学发, 刘寿东
- WEI Nan, ZHANG Mi, WANG Huimin, ZHANG Leiming, WEN Xuefa, LIU Shoudong.
- 散射辐射对亚热带人工针叶林光能利用率的影响
- The impacts of changes in diffuse radiation on light use efficiency in a subtropical plantation coniferous forest
- 生态学报. 2017, 37(10): 3403-3414
- Acta Ecologica Sinica. 2017, 37(10): 3403-3414
- http://dx.doi.org/10.5846/stxb201603080411
-
文章历史
- 收稿日期: 2016-03-08
- 网络出版日期: 2017-02-17





2. 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室, 北京 100101
2. Key Laboratory of Ecosystem Network Observation and Modeling, Synthesis Research Center of Chinese Ecosystem Research Network, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
太阳辐射是植被光合作用的能量来源, 也是气候形成及变化的重要驱动因子[1]。然而, 当天空云量及大气气溶胶改变时, 会影响到达地表的总太阳辐射强度及散射辐射与直接辐射之间的比例, 从而影响陆地生态系统的光合生产力[2]。研究发现, 近55年来中国地面太阳辐射的变化特征与全球变化特征相一致, 即经历了先逐渐降低变暗, 而后逐渐升高变亮的变化过程[3-6]。太阳辐射的变化尤其是散射辐射比例的改变势必会影响陆地生态系统对碳的固定及其光能利用率。
光能利用率 (Light Use Efficiency, LUE) 是表征植物利用太阳能通过光合作用转化有机物的效率[7]。一个地区生态系统生产力的大小与该地区植被光能利用效率有直接关系[8-9]。理论上植被叶片中的叶绿体对光的转化率最高可达20%—25%, 但是自然条件下生长的植被对光能的利用率却不到1%[10-11]。由此可以看出, 找到提高光能利用率的途径成为提高植被生产力的关键问题。森林是重要的陆地生态系统, 其光能利用率的变化得到很多学者的关注。大量研究表明, 当天空云量及大气气溶胶含量增加导致地表接受的太阳辐射中散射辐射的比例增加时, 会使得森林生态系统的光合作用较晴朗天气条件下高[2, 12-14], 从而提高了森林生态系统的光能利用率及固碳能力。近年来, 我国中东部地区常出现严重的雾霾天气[15-17]。雾霾天气的出现会导致地表接受的太阳总辐射减少[6], 而散射辐射的比例增加。因此, 在当前雾霾天气经常出现的条件下研究散射辐射变化对森林生态系统光能利用率的影响变得尤为重要。
人工林是我国森林生态系统的重要组成部分, 面积比居世界首位, 因此其对我国陆地生态系统的碳收支会产生巨大影响[18-20]。我国南方林区以人工林为主, 面积占全国人工林的54.3%, 故在当前人为气溶胶排放增加及气候变化条件下, 研究散射辐射变化对南方人工林光能利用率的影响能够为更好地评价人工林生态系统在区域碳平衡中的作用提供科学依据。
地处江西省千烟洲的亚热带人工针叶林是我国南方亚热带地区典型的人工林生态系统, 大量研究表明, 该生态系统净CO2交换量 (Net Ecosystem Exchange of CO2, NEE) 在5—6月出现最高值, 而太阳辐射强烈少雨的7月份易受高温干旱影响[18, 20-21]。但是同期在散射辐射提高、气温及饱和水汽压差下降的条件下NEE会提高[22]。由此可见, 该生态系统生产力受光以及温度、水分等环境因子的影响。在不受干旱胁迫、温度适宜的条件下, 植被光能利用率决定着该生态系统的生产力, 但是目前该生态系统光能利用率对散射辐射变化的响应, 特别是植被在散射辐射及直接辐射条件下光合特性的差异还不明确。本研究以千烟洲亚热带人工针叶林为研究对象, 利用2003—2012年的30 minCO2通量及常规气象观测资料, 分析太阳辐射中散射辐射的变化对千烟洲亚热带人工针叶林光能利用率的影响, 以及光合有效辐射中散射部分和直接辐射部分对植被光合特性的影响, 从而为准确地评价同区域人工林碳汇能力对气候变化的响应特征提供科学依据。
1 研究地概况及研究方法 1.1 研究地概况千烟洲亚热带人工针叶林位于江西省泰和县, 其通量观测塔 (26°44′29.1″N, 115°03′29.2″E) 位于中国生态研究网络的千烟洲红壤丘陵农业综合开发实验站内。千烟洲地处亚热带大陆性季风气候区, 海拔高度为60—120 m, 年平均气温约为17.8 ℃, 年平均降水量为1360 mm, 年平均太阳总辐射量为4349 MJ/m2[23]。林内主要植被为马尾松 (Pinus Massoniana)、湿地松 (P. elliottii)、杉木 (Cunninghamia lanceolata) 等, 冠层平均高度为12 m, 覆盖率在90%以上。
1.2 研究方法 1.2.1 数据获取本研究获取2003—2012年千烟洲站实时的30 minCO2通量数据及常规气象数据, 包括太阳总辐射数据 (S0)、光合有效辐射数据 (PAR)、温度 (气温Ta及土壤5 cm处的温度Ts)、饱和水汽压差 (VPD)、降水量 (P) 及土壤表层5 cm处的含水量 (SWC)。CO2通量数据是由安装在观测塔39 m高度处的涡度相关系统观测得到, 该观测系统包括三维超声风速仪 (Model CSAT-3, Campbell Scientific) 和红外气体分析仪 (Model Li-7500, Licor Inc)。仪器原始采样频率为10 Hz, 数据采集器 (CR10XTD, CR23XTD, CR5000) 在采集实时数据的同时计算通量数据30 min的平均值并进行存储。辐射数据来自42 m, 气温、水汽压来自23.5 m的观测, 详细的站点及数据材料描述见宋霞等[21]的研究。
1.2.2 数据处理在CO2通量观测过程中, 由于仪器响应误差、下垫面起伏条件、天气状况等因素的影响, 会造成观测数据出现缺失或异常, 因此需要对获取的30 min的CO2通量数据进行处理。具体操作步骤包括:(1) CO2气体浓度的密度校正;(2) 倾斜校正, 即坐标轴旋转;(3) 储存项校正;(4) 数据筛选和插补, 剔除由于仪器故障、天气以及不符合通量观测条件等导致的无效通量数据, CO2通量数据采用查表法进行插补, 常规气象数据采用滑动平均法进行缺失数据插补[14, 18-19, 24], 最终得到连续半小时的净CO2交换量, 即NEE的数据。
1.2.3 NEE组分拆分由于涡度相关法只能直接获取净生态系统CO2交换量 (NEE, mgCO2 m-2 s-1), 该量值为正代表生态系统为碳源, 即释放CO2;该量值为负代表生态系统为碳汇, 即吸收CO2。生态系统水平上总初级生产力 (GPP, mgCO2 m-2 s-1) 无法直接观测, 因此, 需对NEE进行组分拆分[25], 计算得到GPP, 即使用下式:

|
(1) |
式中, Re(Ecosystem Respiration) 表示生态系统呼吸 (mgCO2 m-2 s-1);Re是利用Lloyd-Taylor方程[26], 使用筛选后的全年夜间有效NEE数据与对应有效的土壤5 cm处温度数据对式2进行拟合, 并将拟合的方程外推至白天, 从而计算白天的生态系统呼吸, 最终利用式1计算出GPP。

|
(2) |
式中, Ts为土壤5 cm处温度 (K);Rref为生态系统在参考温度Tref(283.1 K) 下的呼吸值 (mgCO2 m-2 s-1);E0为活化能;T0为常数 (-46.02 ℃, 即227.13 K)。
1.2.4 光能利用率 (LUE)生态系统水平的光能利用率为生态系统总初级生产力与植被上方接受的光合有效辐射的比值[27-29]:
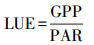
|
(3) |
式中, PAR (photosynthetic active radiation) 表示相应时间段生态系统接受的光合有效辐射量 (μmol m-2 s-1), 因此LUE的单位为 (mgCO2/μmol), 本文进行单位转换, 转换为 (gCO2/mol)。
1.2.5晴空指数 (kt)
晴空指数 (Clearness index, kt) 是指一定太阳高度角下地表接受的总太阳辐射S0 (W/m2) 与地球大气层上方平行于地表面上接受的总太阳辐射Se (W/m2) 的比值[12]:
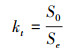
|
(4) |

|
(5) |
式中, Ssc为太阳常数 (1370W/m2), td为日序数, h为太阳高度角。晴空指数是天空晴朗程度的判定指标, 当kt接近于0时, 天空完全被云层覆盖, 到达地面的太阳辐射非常小;当kt=0.5左右时, 为多云天空中等辐射条件, 太阳总辐射下降而散射辐射增加;当kt接近于1时, 天空晴朗无云[14]。
1.2.6 散射辐射本文使用散射辐射分割模型简化后的形式对千烟洲人工针叶林接受的散射辐射进行估算[2, 30-31], 方法如下:

|
(6) |
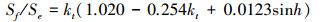
|
(7) |
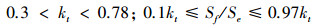
|
(8) |

|
(9) |
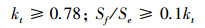
|
(10) |
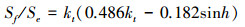
|
(11) |
式中, Sf为地表水平面上接受的散射辐射 (W/m2)。
光合有效辐射的散射辐射部分利用Spitters等[32-33]提出的经验公式计算。
1.2.7 NEE对光的响应本文用Michaelis-Menten方程 (式12) 对多云及晴朗天空条件下千烟洲人工林生态系统的光响应曲线进行拟合[13-14, 34-35]:

|
(12) |
式中, α为生态系统的初始量子效率, 表示弱光条件下植被对光的利用能力 (mgCO2/μmol), 本文进行单位转换, 转换为 (gCO2/mol);β表示生态系统潜在最大光合作用速率 (mgCO2 m-2 s-1);由于NEE为负值表示CO2的吸收, 故α和β都为负值, 所以α值的绝对值越大表明植被弱光下对光的利用能力越强, β值的绝对值越大表明光饱和时的潜在最大光合作用速率越大[2]。
为区分该森林生态系统光合作用对直接辐射、散射辐射的响应特征, 本文使用Gu等[2]提出的植被光响应模型, 分析千烟洲人工林生态系统NEE对散射辐射与直接辐射的光响应特征:

|
(13) |
式中, If和Ir分别表示光合有效辐射中散射辐射部分与直接辐射部分, 单位:μmol m-2 s-1;αf与αr分别表示生态系统在If和Ir下的光量子初始量子效率 (mgCO2/μmol), 本文进行单位转换, 转换为 (gCO2/mol);βf与βr分别表示生态系统在If和Ir下的潜在最大光合作用速率 (mgCO2 m-2 s-1), 由于本模型中采用NEE为正值表示CO2吸收, 故α、β均为正值。
本研究采用11天滑动法[2]对生长旺盛季节不同时段的光响应曲线进行拟合, 以得到生长旺季内不同时段的光合参数, 即从6月1日开始到8月30日, 每11天为一组数据进行拟合, 后一个11天的开头从前一个11天的中间开始, 共拟合17次得出17组不同参数。
2 结果与讨论 2.1 千烟洲亚热带人工针叶林环境因子的季节变化千烟洲亚热带人工针叶林气温和降水季节变化如图 1所示, 气温变化呈单峰型, 多年月平均气温在7月份达到最高值, 为29.1 ℃, 冬季平均最低气温出现在1月份, 为4.9 ℃。该生态系统总体夏季降水多, 冬季降水少, 总体可以看出4—6月气候湿润, 其降水量约占全年降水的33.6%, 而7—9月相对干旱, 这也与王萌萌等[22]对2003—2010年以及孙晓敏等[20]对2003、2004年千烟洲人工林气温和降水季节动态的研究相一致。饱和水汽压差 (VPD) 表征环境大气的水分亏缺状况, 从图 1可以看出:全年中VPD在7、8月份相对较高, 7月达到峰值, 说明在7、8月份, 空气中水分含量相对较少, 比较干旱;土壤表层5 cm处的含水量在1—6月份较多, 平均值为0.21 m3/m3, 7—12月份较少, 平均值为0.14 m3/m3, 这与7—12月份降水量较前半年降水量少相一致。

|
| 图 1 千烟洲亚热带人工针叶林2003—2012年气温 (Ta) 与降水 (P)、饱和水汽压差 (VPD) 与土壤表层5 cm处的含水量 (SWC)、太阳总辐射 (S0) 与散射辐射 (Sf) 的平均季节变化 Fig. 1 Averaged seasonal variation of air temperature (Ta) and precipitation (P), vapor pressure deficit (VPD) and soil water content at 5 cm (SWC), global solar radiation (S0) and diffuse radiation (Sf) in Qianyanzhou subtropical plantation coniferous forests from 2003 to 2012 |
2003—2012年太阳总辐射以及散射辐射季节变化特征如图 1所示:二者都是夏季大于冬季, 在1月份出现最低值, 在7月份达到最大值, 太阳总辐射最大值为633.8 MJ/m2, 散射辐射最大值为318.7 MJ/m2。6月份太阳总辐射相比5月份减少而散射辐射却上升, 这是由于当月降水增多, 多云天空会导致散射辐射增加。
综上所述, 该生态系统光、水、热资源比较丰富, 但是在生长旺盛季节, 即6—8月, 受副热带高压控制, 7月会出现高温少雨的现象, 易出现季节性干旱, 对植被生长会造成一定影响。
2.2 千烟洲亚热带人工针叶林LUE的季节变化从图 2可以看出, 2003—2012年生长旺盛季节内, GPP和LUE都有略微上升的趋势, 但是从P值得出二者都没有通过F检验, 这说明10年间植被本身生长变化对LUE的影响较小, 可以忽略。

|
| 图 2 千烟洲亚热带人工针叶林2003—2012年6—8月生态系统总初级生产力 (GPP) 与光能利用率 (LUE) 年动态 Fig. 2 Inter-annual variation of gross primary productivity (GPP) and light use efficiency (LUE) in Qianyanzhou subtropical plantation coniferous forests from 2003 to 2012 in June to August |
光能利用率的季节变化特征呈双峰型 (图 3), 生长较旺盛季节, 两个高值分别出现在6月和9月, 谷值出现在7月份, 这是由于6月份平均GPP为739.4 gCO2/m2, 平均PAR为874.3 mol/m2, 7月份平均GPP为855.4 gCO2/m2, 平均PAR为1257.8 mol/m2, 虽然7月份的GPP较高, 但7月份PAR相比6月份却猛增, 二者比值减小, 故光能利用率低。

|
| 图 3 千烟洲亚热带人工针叶林2003—2012年光能利用率 (LUE) 的平均季节变化 Fig. 3 Averaged seasonal variation of light use efficiency (LUE) in Qianyanzhou subtropical plantation coniferous forests from 2003 to 2012 |
由此可以看出, PAR最高的月份并非LUE最高的季节, 而在云雨相对较多的6月、9月, LUE较高, 因此, 太阳辐射变化尤其是散射辐射比例的改变会对LUE产生重要影响。
2.3 散射辐射变化对千烟洲亚热带人工针叶林LUE的影响为去除叶面积指数变化与太阳高度角的改变对林冠接受太阳辐射的影响, 本文选择在该区域生长旺盛季节 (6—8月) 的最大太阳高度角范围内 (76.7°—86.7°) 分析散射辐射的改变对LUE的影响, 该太阳高度角范围对应的时段也是一天中植被生理活动较为旺盛的时段。由于各年份变化趋势一致, 图 4—图 8皆以2011年、2012年为例。

|
| 图 4 千烟洲亚热带人工针叶林2011、2012年6—8月太阳高度角在76.7°—86.7°范围内LUE与kt的关系 Fig. 4 Relationship between LUE and ktfor solar elevation angle 76.7°—86.7° in Qianyanzhou subtropical plantation coniferous forests from June to August in 2011 and 2012 |

|
| 图 5 千烟洲人工针叶林2011、2012年6—8月太阳高度角在76.7°—86.7°范围内Sf /S0与kt的关系 Fig. 5 Relationship between Sf /S0 and kt for solar elevation angle 76.7°—86.7° in Qianyanzhou subtropical plantation coniferous forests from June to August in 2011 and 2012 |

|
| 图 6 千烟洲亚热带人工针叶林2011、2012年6—8月太阳高度角在76.7°—86.7°范围内LUE与Sf /S0的关系 Fig. 6 Relationship between LUE and Sf /S0 (solar elevation 76.7°—86.7°) in Qianyanzhou subtropical plantation coniferous forests from June to August in 2011 and 2012 |

|
| 图 7 千烟洲亚热带人工针叶林2011、2012年6—8月太阳高度角在76.7°—86.7°范围内GPP与Sf /S0的关系 Fig. 7 Relationship between GPP and Sf /S0 (solar elevation 76.7°—86.7°) in Qianyanzhou subtropical plantation coniferous forests from June to August in 2011 and 2012 |

|
| 图 8 千烟洲亚热带人工针叶林2011、2012年不同天空条件下的光响应曲线 Fig. 8 Light response curves in Qianyanzhou subtropical plantation coniferous forests under clear sky condition and cloudy sky condition in 2011 and 2012 |
如图 4所示, 光能利用率随晴空指数的变化呈递减趋势, kt值越高, 天空越晴朗, 太阳辐射越强, LUE越小。2011、2012年kt在0.6—0.7范围内的晴朗天气条件下平均光能利用率比kt在0.4—0.5范围内的多云条件下光能利用率减小了46.55%、42.95%, 其它年份如表 1所示, 10年中晴朗天空下的LUE较多云天空下平均减小了44.66%。以往研究发现, 长白山阔叶红松林、鼎湖山亚热带常绿阔叶林、西双版纳热带雨林森林生态系统中, kt在0.4—0.6范围内的中等辐射条件下, 各生态系统的光能利用率都会相应的提高[36-37], 本文研究结果与其一致。
| 年份Year | ||||||||||
| 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | |
| a | 46.27% | 45.98% | 48.48% | 46.68% | 30.24% | 46.28% | 46.60% | 46.57% | 46.55% | 42.95% |
| b | 21.49% | 23.56% | 21.61% | 19.11% | 23.27% | 38.02% | 25.71% | 16.80% | 22.10% | 10.76% |
| a: kt在0.6—0.7范围内的LUE比kt在0.4—0.5范围内的LUE平均减少的百分比; b: Sf /S0在70%—85%之间的LUE比Sf /S0在55%—70%之间的LUE平均提高的百分比 | ||||||||||
千烟洲亚热带人工针叶林在生长旺季太阳高度角76.7°—86.7°范围内, 到达地面的散射辐射 (Sf) 在太阳总辐射中 (S0) 所占的比例Sf/S0随kt的增加而减小 (图 5)。该结果表明天空越晴朗, 散射辐射占总辐射的比例越小。
而千烟洲亚热带人工针叶林, LUE随Sf /S0的增大而增大 (图 6)。10年间, Sf /S0在70%—85%之间的LUE比Sf /S0在55%—70%之间的平均提高了22.24%(表 1)。由此可以看出, 在多云及气溶胶一定程度增多导致总太阳辐射减小而散射辐射增加的天气条件下, LUE增加。
光能利用率的增加与散射辐射促使植被光合作用的增加有关。从图 7中可以看出, 2003—2012年GPP随Sf /S0的增加先增加后减小, 最大值出现在60%—70%附近, 这是由于散射辐射在林冠中具有更强的透射性[38-39], 易入射至林冠下方, 因此, 散射辐射占太阳总辐射增加时, 更多的辐射入射到冠层底部的叶片上, 使林冠中占比例较大只能接受散射辐射进行光合作用的阴生叶的光合作用增加, 从而使整个冠层的光合作用提高[40]。因此, 多云天空或气溶胶一定程度的增加导致散射辐射增加时, 会使植被的光合作用增强。
2.4 千烟洲亚热带人工针叶林光合作用对光的响应 2.4.1 不同天空条件下光合作用的光响应特征利用Michaelis-Menten方程对2003—2012年生长旺季 (6—8月) 多云天气和晴朗天气下该生态系统的光响应曲线进行拟合发现 (图 8):10年中除2008、2009、2010年之外其他年份多云天气条件下的初始量子效率α比晴朗天气条件下平均提高了29.3%(表 2)。这说明在弱光下多云天气条件更有利于千烟洲人工针叶林对光的利用。对2003—2012年多云天气条件下和晴朗天气条件下拟合潜在光合速率参数进行比较发现:除了2008、2009年, 其他年份潜在最大光合速率在多云天气条件下低于晴朗天气条件下的拟合值 (表 2)。
| 年份 Year |
α /(gCO2/mol) | β/(mgCO2 m-2 s-1) | Re /(mgCO2 m-2 s-1) | 相关性系数R2 | |||||||||
| 晴空条件 Clear sky condition |
多云条件 Cloudy sky condition |
c/% | 晴空条件 Clear sky condition |
多云条件 Cloudy sky condition |
d/% | 晴空条件 Clear sky condition |
多云条件 Cloudy sky condition |
晴空条件 Clear sky condition |
多云条件 Cloudy sky condition |
||||
| 2003 | -1.07 | -1.31 | 18.32 | -1.00 | -0.85 | -17.83 | 0.18 | 0.14 | 0.66 | 0.63 | |||
| 2004 | -0.95 | -1.36 | 30.30 | -1.74 | -1.20 | -44.57 | 0.15 | 0.15 | 0.61 | 0.7 | |||
| 2005 | -1.04 | -1.20 | 13.33 | -1.88 | -1.18 | -59.59 | 0.25 | 0.12 | 0.7 | 0.74 | |||
| 2006 | -0.77 | -1.47 | 47.70 | -1.48 | -1.33 | -11.03 | 0.10 | 0.17 | 0.67 | 0.78 | |||
| 2007 | -0.89 | -1.31 | 31.94 | -1.31 | -1.22 | -7.22 | 0.17 | 0.17 | 0.77 | 0.68 | |||
| 2008 | -1.59 | -1.54 | -3.25 | -0.88 | -1.14 | 23.00 | 0.11 | 0.17 | 0.63 | 0.73 | |||
| 2009 | -1.47 | -1.35 | -8.89 | -1.13 | -1.16 | 2.85 | 0.11 | 0.14 | 0.64 | 0.69 | |||
| 2010 | -1.37 | -1.30 | -5.38 | -1.30 | -1.18 | -9.88 | 0.21 | 0.15 | 0.72 | 0.72 | |||
| 2011 | -1.16 | -1.44 | 19.44 | -1.45 | -1.23 | -17.69 | 0.24 | 0.19 | 0.72 | 0.76 | |||
| 2012 | -0.74 | -1.34 | 44.42 | -2.55 | -1.37 | -86.19 | 0.19 | 0.10 | 0.66 | 0.71 | |||
| α:生态系统的初始量子效率The initial quantum efficiency of ecosystem; β:生态系统潜在最大光合作用速率The capacity of a canopy to resist photosynthetic saturation at high levels of PAR; Re:生态系统呼吸Ecosystem respiration; c表示多云天空条件α比晴朗条件下的α增加的百分比; d表示多云天空条件β比晴朗条件下的β增加的百分比 | |||||||||||||
张弥等[14]及Law等[13]的研究发现, 光饱和时潜在最大光合作用速率在多云天空条件下大于晴朗天空条件下的值, 与本研究的结果不一致。这与千烟洲亚热带人工针叶林的植被类型对环境的适应性有关。千烟洲人工林植被类型为马尾松、湿地松、杉木等, 梁春等[41]对马尾松光响应曲线研究表明:1500 μmol m-2 s-1光合有效辐射条件下, 马尾松的光合速率达到最高值, 比光合有效辐射在2000 μmol m-2 s-1条件下的光合速率高, 即强光下, 该树种会产生光抑制现象。本文对光响应曲线比较发现, 无论是多云天气条件还是晴朗天气条件下, 其PAR都会超过1500 μmol m-2 s-1, 即都会对植被的光合作用产生抑制作用, 故与晴朗天空相比较, 千烟洲亚热带人工林光饱和时的潜在光合速率在多云天空条件下并没有明显的增加。
虽然将多云天气条件与晴朗天气条件进行区分, 可以得出两种辐射条件下植被的光合特性, 但是该方法并不能完全将散射辐射与直接辐射分开, 为了进一步探索散射辐射与直接辐射对植被光合的影响, 将光合有效辐射分为散射有效辐射和直接有效辐射进行研究。
2.4.2 植被的光合作用对散射辐射与直接辐射变化的响应特征利用公式 (13) 给出的模型拟合分析了2003—2012年间生长旺盛季节光合作用对散射辐射与直接辐射的响应特征。从散射辐射与直接辐射条件下的初始量子效率αf、αr的变化看出 (图 9), 2005、2006、2008、2009、2010、2012年相类似, 以2012年为例说明, 这些年份中70%以上的数据点αf >αr, 6年中生长旺盛季节αf比αr平均高0.63 gCO2/mol, 由此说明, 气温降水适宜时, 弱光条件下散射辐射比直接辐射更有利于植被对光的利用。而2003、2004、2007、2011年相类似, 以2003年为例可以看出6月份以及8月份αf >αr, 而在7月份αf < αr, 该现象表明, 在降水较少的7月, 即易发生高温干旱的时期, 散射辐射条件下的初始量子效率并未表现出显著的增加。

|
| 图 9 千烟洲亚热带人工针叶林2003、2012年6—8月散射辐射与直接辐射条件下对应的初始量子效率 (αf与αr) 的对比 Fig. 9 The initial quantum efficiency under diffuse radiation condition (αf) and under direct radiation condition (αr) in Qianyanzhou subtropical plantation coniferous forests from June to August in 2003 and 2012 |
2003—2012年间 (图 10, 以2011、2012年为例) 拟合所得散射辐射与直接辐射条件下最大潜在光合速率βf和βr相比, 10年间生长旺季散射辐射下的βf比直接辐射下的βr平均提高了0.81 mgCO2 m-2 s-1, 这说明散射辐射条件下该人工林生态系统光合潜力比直接辐射条件下大, 即散射辐射条件下, 植被的光合潜力增加, 这将有利于提高植被的光能利用率。

|
| 图 10 千烟洲亚热带人工针叶林2011、2012年6—8月散射辐射与直接辐射条件下对应的最大潜在光合速率 (βf与βr) 的对比 Fig. 10 The capacity of a canopy to resist photosynthetic saturation at high levels of PAR under diffuse radiation condition (βf) and under direct radiation condition (βr) in Qianyanzhou subtropical plantation coniferous forests from June to August in 2011 and 2012 |
本研究中αf和αr、βf和βr的结果与Gu等在2002年对欧洲赤松林、白杨林、混合落叶林、草地和冬小麦作物的研究结果基本一致。但本研究中初始量子效率在一些年份中出现散射辐射条件下小于直接辐射条件下的现象, 而出现较多αf < αr的年份在生长旺季都出现不同程度的干旱天气。以2003年为例, 如图 11所示, 与2012年相比, 2003年的7月份土壤含水量偏少, 且气温高, 导致较严重的季节性伏旱, 植被在受到干旱胁迫时, 散射辐射条件下, 初始量子效率并未表现出显著的提高。

|
| 图 11 千烟洲亚热带人工针叶林2003、2012年6—8月气温与土壤表层5 cm处的含水量配置图 Fig. 11 The dynamic of air temperature and soil water content at 5 cm in Qianyanzhou subtropical plantation coniferous forests from June to August in 2003 and 2012 |
10年间生长旺季βf的平均值比βr的平均值均有所提高 (图 10), 这是由于散射辐射在林冠中具有更强的透射性[37-38], 易入射至林冠下方。因此, 在散射辐射增加时, 冠层中占比例较大的阴生叶的光合作用增加, 同时也可避免直射辐射过多导致林冠上部叶片出现光饱和的现象, 因此, 散射辐射的增加会使林冠整体上具有更高的光合潜力[42]。
3 结论通过对千烟洲亚热带人工针叶林光能利用率随散射辐射的变化, 以及散射辐射与直接辐射条件下植被光合作用光响应参数的特征进行分析, 并与同类型研究的比较讨论发现:
散射辐射更有利于提高该生态系统的光能利用率。与多云及气溶胶一定程度增加的中等辐射条件下 (0.4 < kt < 0.5) 相比, 天空较晴朗的强太阳辐射条件下 (0.6 < kt < 0.7) 的LUE平均减少了44.66%;散射辐射与直接辐射的比例Sf /S0在70%—85%之间的LUE比Sf /S0在55%—70%之间的LUE平均提高了22.24%。即散射辐射占太阳总辐射比例增加的条件下, 光能利用率增加。
不受干旱胁迫影响的条件下, 植被光合作用对光的响应表现为散射辐射条件下的初始量子效率αf比直接辐射条件下的αr平均提高0.63 gCO2/mol, 即弱光条件下散射辐射比直接辐射更有利于植被对光的利用。在强光下, 散射辐射增加更有利于植被光合潜力的增加, 从而提高植被的光能利用率。
太阳散射辐射比例改变时, 温度、水分等环境因子也会随之改变, 因此要量化太阳辐射改变对植被光能利用率的影响还需借助过程模型来加以分析。这也将有助于分析环境因子胁迫发生时直接辐射与散射辐射对植被光能利用率的影响及其机制。
| [1] | 杨胜朋, 王可丽, 吕世华. 近40年来中国大陆总辐射的演变特征. 太阳能学报, 2007, 28(3): 227–232. |
| [2] | Gu L H, Baldocchi D, Verma S B, Black T A, Vesala T, Falge E M, Dowty P R. Advantages of diffuse radiation for terrestrial ecosystem productivity. Journal of Geophysical Research, 2002, 107(D6): 4050. |
| [3] | 马金玉, 梁宏, 罗勇, 李世奎. 中国近50年太阳直接辐射和散射辐射变化趋势特征. 物理学报, 2011, 60(6): 853–866. |
| [4] | Wild M, Gilgen H, Roesch A, Ohmura A, Long C N, Dutton E G, Forgan B, Kallis A, Russak V, Tsvetkov A. From dimming to brightening: decadal changes in solar radiation at earth's surface. Science, 2005, 308(5723): 847–850. DOI:10.1126/science.1103215 |
| [5] | Mercado L M, Bellouin N, Sitch S, Boucher O, Huntingford C, Wild M, Cox P M. Impact of changes in diffuse radiation on the global land carbon sink. Nature, 2009, 458(7241): 1014–1071. DOI:10.1038/nature07949 |
| [6] | 齐月, 房世波, 周文佐. 近50年来中国东、西部地面太阳辐射变化及其与大气环境变化的关系. 物理学报, 2015, 64(8): 089201–1. |
| [7] | 卫亚星, 王莉雯. 青海省植被光能利用率模拟研究. 生态学报, 2010, 30(19): 5209–5216. |
| [8] | 李泽晖, 王云龙, 魏远, 马悦, 陈润芝. 湖南岳阳杨树人工林光能利用率动态特征分析. 资源科学, 2012, 34(10): 1832–1838. |
| [9] | 穆少杰, 李建龙, 周伟, 杨红飞, 章超斌, 居为民. 2001-2010年内蒙古植被净初级生产力的时空格局及其与气候的关系. 生态学报, 2013, 33(12): 3752–3764. |
| [10] | 解锋. 论提高光能利用率的途径. 陕西农业科学, 2008, 54(1): 144–146. |
| [11] | 徐恒永, 赵君实, 徐茂臻. 高产冬小麦光能利用率的研究. 莱阳农学院学报, 1994, 11(1): 6–10. |
| [12] | Gu L H, Fuentes J D, Shugart H H, Staebler R M, Black T A. Responses of net ecosystem exchanges of carbon dioxide to changes in cloudiness: results from two North American deciduous forests. Journal of Geophysical Research: Atmospheres, 1999, 104(D24): 31421–31434. DOI:10.1029/1999JD901068 |
| [13] | Law B E, Falge E, Gu L, Baldocchi D D, Bakwin P, Berbigier P, Davis K, Dolman A J, Falk M, Fuentes J D, Goldstein A, Granier A, Grelle A, Hollinger D, Janssens I A, Jarvis P, Jensen N O, Katul G, Mahli Y, Matteucci G, Meyers T, Monson R, Munger W, Oechel W, Olson R, Pilegaard K, Paw U K T, Thorgeirsson H, Valentini R, Verma S, Vesala T, Wilson K, Wofsy S. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agricultural and Forest Meteorology, 2002, 113(1/4): 97–120. |
| [14] | 张弥, 于贵瑞, 张雷明, 孙晓敏, 温学发, 韩士杰. 太阳辐射对长白山阔叶红松林净生态系统碳交换的影响. 植物生态学报, 2009, 33(2): 270–282. |
| [15] | 丁一汇, 柳艳菊. 近50年我国雾和霾的长期变化特征及其与大气湿度的关系. 中国科学:地球科学, 2014, 44(1): 37–48. |
| [16] | 张人禾, 李强, 张若楠. 2013年1月中国东部持续性强雾霾天气产生的气象条件分析. 中国科学:地球科学, 2014, 44(1): 27–36. |
| [17] | 吴兑. 近十年中国灰霾天气研究综述. 环境科学学报, 2012, 32(2): 257–269. |
| [18] | 刘允芬, 于贵瑞, 温学发, 王迎红, 宋霞, 李菊, 孙晓敏, 杨风亭, 陈永瑞, 刘琪珠. 千烟洲中亚热带人工林生态系统CO2通量的季节变异特征. 中国科学D辑地球科学, 2006, 34(S1): 91–102. |
| [19] | 刘允芬, 宋霞, 孙晓敏, 温学发, 陈永瑞. 千烟洲人工针叶林CO2通量季节变化及其环境因子的影响. 中国科学D辑地球科学, 2004, 34(S2): 109–117. |
| [20] | 孙晓敏, 温学发, 于贵瑞, 刘允芬, 刘琪璟. 中亚热带季节性干旱对千烟洲人工林生态系统碳吸收的影响. 中国科学D辑地球科学, 2006, 36(S1): 103–110. |
| [21] | 宋霞, 刘允芬, 徐小锋, 于贵瑞, 温学发. 红壤丘陵区人工林冬春时段碳、水、热通量的观测与分析. 资源科学, 2004, 26(3): 96–104. |
| [22] | 王萌萌, 张弥, 王辉民, 张雷明, 杨沈斌. 太阳辐射变化对千烟洲亚热带人工针叶林净CO2交换量的影响. 生态学杂志, 2015, 34(2): 303–311. |
| [23] | 韩佳音, 李胜功, 张雷明, 温学发, 李庆康, 王辉民. 江西千烟洲地区散射辐射的模拟与验证. 应用生态学报, 2015, 26(10): 2991–2999. |
| [24] | 张雷明, 于贵瑞, 孙晓敏, 温学发, 任传友, 宋霞, 刘允芬, 关德新, 闰俊华, 张一平. 中国东部森林样带典型生态系统碳收支的季节变化. 中国科学D辑地球科学, 2006, 36(S1): 45–59. |
| [25] | 黄昆, 王绍强, 王辉民, 仪垂祥, 周蕾, 刘允芬, 石浩. 中亚热带人工针叶林生态系统碳通量拆分差异分析. 生态学报, 2013, 33(17): 5252–5265. |
| [26] | Lloyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315–323. DOI:10.2307/2389824 |
| [27] | Gilmanov T G, Tieszen L L, Wylie B K, Flanagan L B, Frank A B, Haferkamp M R, Meyers T P, Morgan J A. Integration of CO2 flux and remotely-sensed data for primary production and ecosystem respiration analyses in the Northern Great Plains: potential for quantitative spatial extrapolation. Global Ecology and Biogeography, 2005, 14(3): 271–292. DOI:10.1111/j.1466-822X.2005.00151.x |
| [28] | 赵育民, 牛树奎, 王军邦, 李海涛, 李贵才. 植被光能利用率研究进展. 生态学杂志, 2007, 26(9): 1471–1477. |
| [29] | Gilmanov T G, Soussana J F, Aires L, Allard V, Ammann C, Balzarolo M, Barcza Z, Bernhofer C, Campbell C L, Cernusca A, Cescatti A, Clifton-Brown J, Dirks B O M, Dore S, Eugster W, Fuhrer J, Gimeno C, Gruenwald T, Haszpra L, Hensen A, Ibrom A, Jacobs A F G, Jones M B, Lanigan G, Laurila T, Lohila A, Manca G, Marcolla B, Nagy Z, Pilegaard K, Pinter K, Pio C, Raschi A, Rogiers N, Sanz M J, Stefani P, Sutton M, Tuba Z, Valentini R, Williams M L, Wohlfahrt G. Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agriculture, Ecosystems & Environment, 2007, 121(1/2): 93–120. |
| [30] | 孙敬松, 周广胜. 散射辐射测量及其对陆地生态系统生产力影响的研究进展. 植物生态学报, 2010, 34(4): 452–461. |
| [31] | Reindl D T, Beckman W A, Duffie J A. Diffuse fraction correlations. Solar Energy, 1990, 45(1): 1–7. DOI:10.1016/0038-092X(90)90060-P |
| [32] | Spitters C J T. Separating the diffuse and direct component of global radiation and its implications for modeling canopy photosynthesis part Ⅱ. Calculation of canopy photosynthesis. Agricultural and Forest Meteorology, 1986, 38(1/3): 231–242. |
| [33] | Spitters C J T, Toussaint H A J M, Goudriaan J. Separating the diffuse and direct component of global radiation and its implications for modeling canopy photosynthesis part Ⅰ. Components of incoming radiation. Agricultural and Forest Meteorology, 1986, 38(1/3): 217–229. |
| [34] | Goulden M L, Daube B C, Fan S M, Sutton D J, Bazzaz A, Munger J W, Wofsy S C. Physiological responses of a black spruce forest to weather. Journal of Geophysical Research: Atmospheres, 1997, 102(D24): 28987–28996. DOI:10.1029/97JD01111 |
| [35] | Aubinet M, Chermanne B, Vandenhaute M, Longdoz B, Yernaux M, Laitat E. Long term carbon dioxide exchange above a mixed forest in the Belgian Ardennes. Agricultural and Forest Meteorology, 2001, 108(4): 293–315. DOI:10.1016/S0168-1923(01)00244-1 |
| [36] | 张弥, 吴家兵, 关德新, 施婷婷, 陈鹏狮, 纪瑞鹏. 长白山阔叶红松林主要树种光合作用的光响应曲线. 应用生态学报, 2006, 17(9): 1575–1578. |
| [37] | Zhang M, Yu G R, Zhuang J, Gentry R, Fu Y L, Sun X M, Zhang L M, Wen X F, Wang Q F, Han S J, Yan J H, Zhang Y P, Wang Y F, Li Y N. Effects of cloudiness change on net ecosystem exchange, light use efficiency, and water use efficiency in typical ecosystems of China. Agricultural and Forest Meteorology, 2011, 151(7): 803–816. DOI:10.1016/j.agrformet.2011.01.011 |
| [38] | Roderick M L, Farquhar G D, Berry S L, Noble I R. On the direct effect of clouds and atmospheric particles on the productivity and structure of vegetation. Oecologia, 2001, 129(1): 21–30. DOI:10.1007/s004420100760 |
| [39] | Yamasoe M A, von Randow C, Manzi A O, Schafer J S, Eck T F, Holben B N. Effect of smoke and clouds on the transmissivity of photosynthetically active radiation inside the canopy. Atmospheric Chemistry and Physics, 2006(6): 1645–1656. |
| [40] | Williams M, Rastetter E B, var der Pol L, Shaver G R. Arctic canopy photosynthetic efficiency enhanced under diffuse light, linked to a reduction in the fraction of the canopy in deep shade. New Phytologist, 2014, 202(4): 1267–1276. DOI:10.1111/nph.12750 |
| [41] | 梁春, 林植芳, 孔国辉. 不同光强下生长的亚热带树苗的光合-光响应特性的比较. 应用生态学报, 1997, 8(1): 7–11. |
| [42] | 何学兆, 周涛, 贾根锁, 张自银, 李秀娟, 赵超, 冯胜辉. 光合有效辐射总量及其散射辐射比例变化对森林GPP影响的模拟. 自然资源学报, 2011, 26(4): 619–634. DOI:10.11849/zrzyxb.2011.04.008 |