 2017, Vol. 37
2017, Vol. 37文章信息
- 王继丰, 韩大勇, 王建波, 付晓玲, 朱道光, 刘赢男, 曹宏杰, 黄庆阳, 谢立红, 钟海秀, 隋心, 倪红伟
- WANG Jifeng, HAN Dayong, WANG Jianbo, FU Xiaoling, ZHU Daoguang, LIU Yingnan, CAO Hongjie, HUANG Qingyang, XIE Lihong, ZHONG Haixiu, SUI Xin, NI Hongwei.
- 三江平原湿地小叶章群落沿土壤水分梯度物种组成及多样性变化
- Variations in plant species composition and diversity of Calamagrostis angustifolia community along soil water level gradient in the Sanjiang Plain
- 生态学报. 2017, 37(10): 3515-3524
- Acta Ecologica Sinica. 2017, 37(10): 3515-3524
- http://dx.doi.org/10.5846/stxb201602250328
-
文章历史
- 收稿日期: 2016-02-25
- 网络出版日期: 2017-02-17





2. 黑龙江省科学院自然与生态研究所, 湿地与生态保育国家地方联合工程实验室, 黑龙江省湿地与恢复生态学重点实验室, 哈尔滨 150040
2. Institute of Natural Resource and Ecology, Heilongjiang Academy of Sciences, National and Provincial Joint Engineering Laboratory of Wetlands and Ecological Conservation, Key Laboratory for Wetland and Restoration Ecology in Heilongjiang Province, Harbin 150040, China
群落多样性是生物多样性研究的核心内容之一[1]。物种多样性沿环境梯度的变化规律已经成为生物多样性研究的中心议题之一[2]。物种多样性是物种演化的空间范围和对特定环境的生态适应, 是最直接和最适合研究生物多样性的层次[3-4], 可反映出群落在组成、结构和功能和动态等方面的异质性[5]。水分作为湿地植物多样性变化的主要驱动因子, 亦是植物生长的重要因子, 在全球变化背景下, 研究水分环境梯度下湿地植物群落的物种多样性变化显得尤为重要[6-8]。
三江平原是中国最大的淡水湿地集中分布区, 亦是中国五大淡水和湿地生物多样性的关键区域之一, 蕴藏着丰富的生物多样性[9-10], 小叶章湿地是该区主要的湿地类型[11]。小叶章湿地主要有典型草甸小叶章湿地(地表无积水)、沼泽化草甸小叶章湿地(季节性积水)和沼泽小叶章湿地(常年积水)[12]。目前, 小叶章湿地的研究主要集中在小叶章种群数量特征[13]、生产力[14]、枯落物[15]、生理生态[16-17]等方面, 在土壤环境因子[18-19]、温室气体排放亦开展了大量[20-21]的研究, 有关研究生物多样性方面的研究主要集中在湿地斑块破碎化对植物多样性的影响[22]、氮沉降对土壤微生物群落多样性的影响[23-24]等方面, 但对三江平原小叶章湿地沿水分环境梯度上物种多样性变化研究尚未见报道。本文以三江平原典型湿地小叶章(Calamagrostis angustifolia)植物群落为研究对象, 分析了不同水分演替阶段小叶章群落的结构组成、α多样性和β多样性特征, 揭示了湿地植物群落在不同水分演替阶段的多样性变化规律, 这为全球变化条件下湿地物种多样性与生态系统功能的关系研究以及生物多样性的保护提供科学的理论依据。
1 材料与方法 1.1 研究区域概况研究区位于我国洪河国家级自然保护区, 其保留了我国面积最大的沼泽区——三江平原原始沼泽景观, 是三江平原湿地的“缩影”, 为未被破坏、保持完整的原始沼泽生态系统, 是我国目前湿地类型最全、保持最完好的原始湿地[25], 地理位置47°42′18″—47°52′07″N, 133°34′ 38″—133°46′29″E。保护区面积21835.73hm2, 地势较为平坦, 海拔51.5—54.5m, 地面坡降1/5000—1/10000;区内河流皆为沼泽性河流, 仅浓江河支流沃绿兰河贯穿整个保护区, 全长7km, 为保护区的主要水源之一;土壤类型主要以白浆土、沼泽土和泛滥地土壤为主;气候属明显的温带季风气候, 四季分明, 冬季漫长、严寒多雪, 春季多风少雨, 夏季炎热, 秋季短暂;年平均降水量为585mm, 降雨多集中于7—9月份, 年平均蒸发量为1166mm;年平均气温为1.9 ℃, 最冷月份平均气温-23.4 ℃, 最热平均气温为22.4 ℃;全年冻结期为7个月左右, 沼泽土壤冻层深80—160cm;霜降一般出现在9月下旬。
1.2 样地设计与取样方法依据小叶章湿地水分差异, 划分为小叶章典型草甸、小叶章沼泽化草甸和小叶章沼泽。小叶章典型草甸土壤含水量60%—80%, 群落盖度95%以上, 以小叶章为优势种, 伴生有垂梗繁缕(Stellaria radians)、二歧银莲花(Anemone dichotoma)、五脉山黧豆(Lathyrus quinquenervius)、箭头唐松草(Thalictrum simplex)等;小叶章沼泽化草甸土壤含水量80%—120%, 呈现季节性积水, 群落盖度达85%—95%, 以小叶章为优势种, 以灰脉苔草(Carex appendiculata)、五脉山黧豆、毛果苔草(C. miyabei var. maopengensis)等为主要伴生种;小叶章沼泽地表常年积水, 积水深度5—20cm, 以漂筏苔草(Carex pseudo-conica)和小叶章为共优势种, 伴生有毛果苔草、狭叶甜茅(Glyceria spiculosa)、忽略野青茅(Calamagrostis neglecta), 偶见细叶沼柳(Salix rosmarinifolia)、越桔柳(Salix myrtilloides)等。
在上述三种类型湿地中开展植被调查, 采用样方法, 随机选取样方, 样方面积1m×1m, 记录样方内植物种类、盖度、多度、高度、密度。于2010年和2011年的7月份开展调查, 每个群落类型调查样方数30个, 共计90个样方, 每个群落的土壤含水量采用烘干法测量, 计算公式为土壤含水量(重量%)=(原土重-烘干土重)/烘干土重×100%, 每个群落的取样样本数为15个, 取地表以下0—15cm的泥炭土, 经测定, 小叶章典型草甸土壤含水量为65%±12%, 小叶章沼泽化草甸为94%±10%, 小叶章沼泽为180%±35%。
1.3 计算方法(1) 重要值
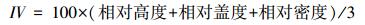
|
(2) 物种丰富度指数
Patrick指数:
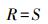
|
(3)α多样性指数
Simpson优势度指数:
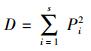
|
Shannon-Wiener指数:
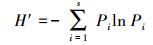
|
Pielou均匀度指数:

|
式中, Pi为第i种的重要值, S为样方中出现的物种数。
(4) 物种周转指数
Jaccard指数
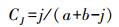
|
式中, j为两个群落共有种数, a和b分别为样地A和样地B的物种数。
Cody指数
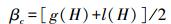
|
式中, g(H)是沿生境梯度H增加的物种数, l(H)是沿生境梯度H失去的物种数, 即在上一个梯度中存在而在下一个梯度中没有的物种数。
(5) β多样性指数
倍性β多样性指数
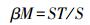
|
加性β多样性指数
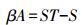
|
式中,ST为样地出现的总物种数,S为样方物种丰富度均值。
1.4 数据分析采用指示种分析检测小叶章群落在3种生境的指示性物种, 采用非度量多维尺度分析(non-metric multidimensional scaling, NMDS)检验群落分布和物种组成。用Excel 2010和Simplot 2000软件进行数据整理, 采用Pearson相关性分析方法进行统计分析。非度量多维尺度分析用R统计语言的Vegan程序包[26]实现, 指示种分析用labdsv程序包[27]实现。
2 结果与分析 2.1 物种组成特征变化表 1列出了沿土壤水分梯度小叶章群落优势种的重要值分布情况。典型草甸和沼泽化草甸以小叶章占据绝对优势地位, 重要值分别达到58.43和64.46, 沼泽群落以小叶章和漂筏苔草共优, 重要值分别为33.26和38.65。其中, 典型草甸具有显著指示意义的物种有二歧银莲花、垂梗繁缕、直穗苔草(Carex orthostachys)、北山莴苣(Mulgedium sibiricum)、林拉拉藤(Galium paradoxum)和莫石竹(Moehringia lateriflora)。沼泽化草甸指示种包括灰脉苔草、五脉山黧豆、球尾花(Lysimachia thyrsiflora)和小叶章(Calamagrostis angustifolia)。沼泽指示种包括漂筏苔草、毛果苔草、狭叶甜茅、燕子花(Iris laevigata)、东北沼委陵菜(Comarum palustre)和地耳草(Hypericum japonicum)(表 1)。
| 物种 Species |
缩写 Abbrev. |
典型草甸 Typical meadow |
沼泽化草甸 Wet meadow |
沼泽 Swamp |
| 小叶章Calamagrostis angustifolia | Caan | 58.43±8.52 | 64.46±7.85(0.41**) | 33.26±3.65 |
| 灰脉苔草Carex appendiculata | Caap | 3.24±0.24 | 10.21±1.97(0.74**) | 1.34±0.20 |
| 五脉山黧豆Lathyrus quinquenervius | Laqu | 2.57±0.32 | 5.77±0.58(0.47**) | 2.02±0.22 |
| 球尾花Lysimachia thyrsiflora | Lyth | 0.18±0.03 | 3.23±0.38(0.43**) | 2.26±0.20 |
| 漂筏苔草Carex pseudo-conica | Caps | 2.26±0.10 | 1.83±0.26 | 38.65±4.01(0.91**) |
| 毛果苔草Carex miyabei var. maopengensis | Cami | 1.78±0.09 | 5.93±1.12 | 11.34±1.58(0.58**) |
| 狭叶甜茅Glyceria spiculosa | Glsp | 5.02±0.87 | 1.96±0.13 | 6.18±2.03(0.39**) |
| 燕子花Iris laevigata | Irla | 1.55±0.25(0.30*) | ||
| 东北沼委陵菜Comarum palustre | Copa | 0.09±0.01 | 0.96±0.17(0.23*) | |
| 地耳草Hypericum japonicum | Hyja | 0.14±0.02 | 0.59±0.15(0.21*) | |
| 二歧银莲花 Anemone dichotoma | Andi | 7.05±0.99(0.83**) | ||
| 垂梗繁缕Stellaria radians | Stra | 3.62±0.32(0.61**) | ||
| 直穗苔草Carex orthostachys | Caor | 3.17±0.45(0.39**) | ||
| 北山莴苣Mulgedium sibiricum | Musi | 2.65±0.27(0.35**) | ||
| 林拉拉藤Galium paradoxum | Gapa | 1.21±0.78(0.30**) | ||
| 莫石竹Moehringia lateriflora | Mola | 0.98±0.21(0.17*) | ||
| 绣线菊Spiraea salicifolia | Spsa | 0.98±0.22(0.13) | ||
| 箭头唐松草Thalictrum simplex | Thsi | 0.68±0.14(0.13) | ||
| 广布野豌豆Vicia cracca | Vicr | 0.77±0.15(0.13NS) | ||
| 芦苇Phragmites communis | Phco | 0.39±0.04(0.09NS) | 0.30±0.05 | |
| 地瓜苗Lycopus lucidus var. hirtus | Lylu | 0.16±0.02 | 0.45±0.09(0.09NS) | |
| 水问荆Equisetum fluviatile | Eqfl | 1.25±0.13(0.13NS) | ||
| 翻白蚊子草Filipendula intermedia | Fiin | 0.34±0.03 | 0.40±0.05(0.04NS) | |
| 臌囊苔草Carex physodes | Caph | 0.32±0.02 | 0.35±0.03(0.03NS) | |
| 忽略野青茅Calamagrostis neglecta | Cane | 0.50±0.06 | 0.96±0.18(0.10NS) | |
| ** P < 0.01, * P < 0.05, NS: no significance | ||||
非度量多维尺度排序结果揭示了小叶章群落物种组成在水分梯度上的变化, Stress值为0.148, 表明排序结果良好。图中第一轴从左至右的梯度基本代表样方从沼泽至草甸排列, 梯度明显, 第二轴从上至下的梯度代表从沼泽化草甸至沼泽的分布, 代表梯度比较短。从样方分布看, 沼泽和沼泽化草甸的样方分布集中, 而草甸的样方分布发散, 表明典型草甸的物种组成差异更大, 而沼泽和沼泽化草甸的物种组成差异较小。结合物种分布看, 沼泽和沼泽化草甸大部分物种亦表现出集中分布, 而典型草甸物种呈零散分布, 说明典型草甸物种大多局限分布于特定样方, 而沼泽和沼泽化草甸物种大多分布于多个样方, 反映出群落中常见种和稀有种数的变化(图 1)。

总共记录到55种维管植物, 隶属于24科39属, 其中, 沼泽总共记录到16种, 沼泽化草甸26种, 草甸39种。
沿土壤水分梯度, 小叶章群落α多样性的变化如图 2。其中, 物种丰富度表现为逐渐减少的趋势, 典型草甸的物种丰富度最高, 为9.38, 沼泽的物种丰富度最低, 为7.38。以典型草甸群落物种丰富度为基准, 沿水分梯度从典型草甸到沼泽物种减少比率为16.31%和21.32%。

|
| 图 2 三江平原小叶章群落沿土壤水分梯度α多样性变化 Fig. 2 Changes of α diversity of Calamagrostis angustifolia community along a soil water-level gradient in the Sanjing plain |
群落的优势度表现为逐渐降低的趋势, 与物种丰富度的变化趋势基本一致。而群落的多样性、均匀度表现为逐渐上升的趋势, 与群落的物种丰富度和优势度正好相反。
同样以典型草甸群落为基准, 沿水分梯度从典型草甸到沼泽群落优势度减少比率为7.32%和40.32%, 物种多样性增加比率为7.67%和31.50%, 均匀度增加比率为3.24%和57.39%。由此可见, 随着水分的增加, 群落优势度、物种多样性和均匀度的变化趋势较为明显, 尤其是从沼泽化草甸至沼泽变化最为明显(图 2)。k-优势度曲线结果也显示出沼泽和沼泽化草甸优势度曲线相似, 均表现出短而陡峭的特征, 而典型草甸优势度曲线长且平缓, 表明沼泽和沼泽化草甸的物种多样性和均匀度接近, 且均低于典型草甸(图 3)。

|
| 图 3 三江平原湿地小叶章群落沿土壤水分梯度k-优势度曲线 Fig. 3 The k-dominance curves of Calamagrostis angustifolia community along a soil water-level gradient in the Sanjing plain |
土壤水分与Simpson优势度指数呈显著负相关关系, 与Shannon-Wienner多样性指数和Pielou均匀度指数呈显著的正相关关系, 说明从小叶章典型草甸至小叶章沼泽, 随着土壤水分的增加, 群落优势度呈现降低趋势, 群落多样性和均匀度则呈现上升趋势(表 2)。
| R | D | H′ | E | |
| SW | -0.325 | -0.557* | 0.861* | 0.805* |
| R:物种丰富度; D: Simpson优势度指数; H′: Shannon-Wienner指数; E: Pielou均匀度; SW, 土壤含水量; * P < 0.05 | ||||
由表 3可以看出, 沼泽与沼泽化草甸的相似性系数(CJ、CS)高于典型草甸的相似性系数, 而沼泽化草甸与典型草甸的相似性系数最低, 说明随着水分梯度的增加相似性系数呈现增加的趋势, 沼泽与沼泽化草甸有更高的相似性。
| 群落类型Community types | |||
| β多样性β diversity | M | W | S |
| 加性β多样性Additive β diversity (βA) | 30.3 | 20.1 | 10.4 |
| 倍性β多样性Multiplicative β diversity (βM) | 4.48 | 4.26 | 2.50 |
| 物种周转Species turnover | M-W | M-S | W-S |
| Jaccard指数Jaccard index (CJ) | 0.20 | 0.35 | 0.24 |
| Bray-Curtis指数Bray-Curtis similarity (CS) | 0.33 | 0.52 | 0.39 |
| 共有种数No. of shared species (RS) | 4 | 5 | 7 |
| 增加物种数No. of increasing species (RI) | 5 | 10 | 7 |
| 减少物种数No. of decreasing species (RD) | 6 | 12 | 6 |
| Cody指数Cody index (βc) | 5.5 | 11 | 6.5 |
沼泽化草甸和小叶章沼泽“增加的物种数”和“减少的物种数”少于典型草甸与沼泽化草甸, 其Cody指数表现出相反的趋势, 从沼泽至典型草甸, 随着生境水分的降低, 群落间共有物种数逐渐减少β多样性指数(βA和βM)逐渐加大, β多样性增加, 亦是植物群落在水分梯度上的基本演替趋势(表 3)。
3 讨论 3.1 群落物种组成本研究对三江平原小叶章湿地沿水分梯度上植物群落组成的探讨发现, 小叶章均为各群落的优势种, 但随着水分梯度的增加, 从典型草甸到沼泽, 小叶章的优势度逐渐降低, 在沼泽中小叶章甚至成为亚优势种, 同时, 各群落的主要伴生种存在一定差异。造成上述差异主要与各群落的生境差异有关。从典型草甸到沼泽, 水分汇集量增加, 适应这种生境, 群落的伴生种由垂梗繁缕、二歧银莲花、苦参等旱中生境生长的物种, 逐渐过渡到球尾花、灰脉苔草等湿润环境生长的物种, 再过渡到东北沼委陵菜、水问荆、睡菜等水生生境生长的物种。这与娄彦景等[28]的结论相同。当然, 除了生境的水分差异外, 造成上述变化还与土壤理化性质、微地形等环境因子密切相关。
3.2 α多样性物种丰富度是生态学的经典概念, 是物种多样性测度中较为简单且生物学意义明显的指数[29-30]。本研究中三江平原小叶章湿地从典型草甸到沼泽群落的物种丰富度呈现逐渐降低的格局。分析其成因, 典型草甸生境土壤含水量适中, 适宜旱、中生植物类群的生长, 沼泽化草甸呈现季节性积水, 湿生植物类群在该群落占主导, 沼泽群落中长期积水的生境只有少数能长期适应厌氧的植物物种生存, 并且旱生植物种类和数量要大于湿生和水生植物类群。因此, 在水分环境梯度上从典型草甸到沼泽群落的物种丰富度逐渐降低, 这与Heather[31]的研究结论基本一致。但物种丰富度与土壤水分的相关关系不显著, 可能是植物通过长期演化进而适应该生境, 形成了自身某些特殊的特征属性, 形成较宽的生态位, 故对土壤水分的变化响应相对不敏感[32]。物种丰富度与均匀度指数、多样性指数的变化格局表现出相反的趋势, 这与以往的多数研究中多样性指数与丰富度指数呈现密切的正相关的结论[33-34]不一致, 分析上述变化的成因, 在水分梯度上3个小叶章湿地积水状况、腐殖质层厚度、土壤类型各异, 造成生境异质性差异较大, 典型草甸是3个群落类型中生境多样性最高的一个类型, 造就了部分物种的最适生境, 导致群落偶见种和稀有种出现的几率加大。有关小叶章群落土壤种子库的研究表发现, 群落植物种类和优势种数量为小叶章典型草甸最多, 小叶章沼泽化草甸其次, 小叶章沼泽最少[35], 亦为本文研究中物种丰富度和多样性指数变化格局与多数研究相反的结论提供了良好的佐证。
本研究中Simpson优势度指数、Shannon-Wienner多样性指数和Pielou均匀度指数与土壤水分表现出显著的相关关系, 受水分的影响显著, 其变化变化格局与多数研究结论一致[36-38]。从典型草甸至沼泽, 随着土壤水分的增加, 生境异质性逐渐降低, 种间竞争呈现降低趋势, 植物分布更加均匀。作为3个群落优势种的小叶章, 具有较大的生态位宽度, 但典型草甸无疑成为其最适的生态位生境, 其种群生长迅速, 在群落中占绝对优势, 此外从土壤营养因子状况分析, 从典型草甸至沼泽土壤中的速效氮、磷亦呈现逐渐降低趋势, 优势种小叶章的环境资源发生改变, 进而改变自身种群的生长策略。
3.3 β多样性和物种周转鉴于β多样性概念和测度方法的混用, 研究采用Vellend[39]的提法将β多样性指数区分为β多样性和物种周转两大类, 前者以Whittaker的β多样性指数及由其衍生出来指数为测度, 后者以样方对之间的诸多相似性系数为测度, 代表了物种沿环境梯度的替代过程[40-41]。Jaccard指数、Sorenson指数Cody指数可反映物种多样性沿环境梯度的物种组成分化程度。本研究中沼泽化草甸与沼泽群落间的相似性最大, 典型草甸与沼泽化草甸群落间的相似性最小, 沼泽化草甸与沼泽群落处于相对稳定的阶段, 群落结构和组成相对较为成熟。在水分梯度上, 从典型草甸到沼泽, 随着水分的增加, 群落间共有种数增加, Cody指数减小, 物种替代速率逐渐降低, 有研究表明湿地植被分布格局及演替与水分条件密切相关[42]。
Whittaker的β多样性由于建立了α多样性和γ多样性之间的数学联系, 常用于探讨不同尺度物种丰富度的差异及相应的生态过程。目前主要有加性分配[43]和倍性分配[44]两种计算方法。本研究中, 两种分配方法得到的βA和βM具有相似的变化格局, 均以典型草甸最高, 沼泽最低。根据加性分配方法得到的βA与总物种数的比例可用来估计β多样性对γ多样性的贡献, 据此计算, 典型草甸、沼泽化草甸和沼泽的β多样性贡献率分别为77.7%、77.3%和65%, 表现出与β多样性类似的梯度变化, 相应地α多样性贡献率表现为增加趋势, 这个结果说明在典型草甸中稀有种更多, 而在沼泽化草甸和沼泽中, 常见种更多。这种格局的深层次机制可能来自于生境多样性效应, 沼泽化草甸和沼泽中过多的地表水分限制了大部分物种的定居, 两种生境相似性程度高, 在NMDS分析中也表明了二者间的环境梯度较短, 物种周转速率低, 因此,物种多样性与局域α多样性关系更密切,而典型草甸地表水分较少,环境异质性较高, 可接纳更多的物种, 由此增加β多样性表明典型草甸的物种多样性更多来自于较高的β多样性,而较少依赖于局域α多样性。这个结果初步显示小叶章群落在典型草甸和沼泽生境中具有不同的多样性维持机制, 前者以扩散过程为主, 后者以生态位分化过程为主。
4 结论三江平原湿地小叶章群落随着土壤水分的增加, 从小叶章典型草甸至小叶章沼泽, 指示物种发生明显改变, 物种组成差异逐渐减小;物种丰富度和Simpson优势度指数呈现逐渐降低趋势, Shannon-Wienner多样性指数和Pielou均匀度指数表现为逐渐上升趋势, 其中Simpson优势度指数、Shannon-Wienner指数和Pielou均匀度指数受土壤水分影响显著;物种周转指数表现为, 小叶章沼泽与小叶章沼泽化草甸有更高的相似性(CJ、CS), 群落间共有物种数逐渐增加β多样性指数(βA和βM)逐渐降低。水分是影响湿地植物群落物种组成和多样性变化的主导因子, 此外, 还受到土壤营养因子、微地形等环境因子以及自身特性的影响, 尚需进一步探讨。
小叶章湿地是三江平原湿地的典型代表, 其植物群落组成及物种多样性的变化格局是三江平原湿地植物群落沿环境梯度变化的典型缩影, 尤其在三江湿地面积日益缩小, 地下水位不断降低的趋势下, 针对典型草甸和沼泽物种多样性维持机制的差异, 制定相应的科学管理制度和恢复措施具有紧迫的现实意义。
| [1] | 刘旻霞, 王刚. 高寒草甸植物群落多样性及土壤因子对坡向的响应. 生态学杂志, 2013, 32(2): 259–265. |
| [2] | Kratochwil A. Biodiversity in ecosystems: some principles//Kratochwil A, ed. Biodiversity in Ecosystems: Principles and Case Studies of Different Complexity Levels. Dordrecht: Kluwer Academic Publishers, 1999: 5-38. |
| [3] | Bråthen K A, Ravolainen V T. Niche construction by growth forms is as strong a predictor of species diversity as environmental gradients. Journal of Ecology, 2015, 103(3): 701–713. DOI:10.1111/jec.2015.103.issue-3 |
| [4] | 王永健, 陶建平, 彭月. 陆地植物群落物种多样性研究进展. 广西植物, 2006, 26(4): 406–411. |
| [5] | 菇文明, 张金屯, 张峰, 张桂萍, 刘瑞祥. 历山森林群落物种多样性与群落结构研究. 应用生态学报, 2006, 17(4): 561–566. |
| [6] | Liu W X, Zhang Z, Wan S Q. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Global Change Biology, 2009, 15(1): 184–195. DOI:10.1111/gcb.2009.15.issue-1 |
| [7] | Niu S L, Yang H J, Zhang Z, Wu M Y, Lu Q, Li L H, Han X G, Wan S Q. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe. Ecosystems, 2009, 12(6): 915–926. DOI:10.1007/s10021-009-9265-1 |
| [8] | Bai Y F, Wu J G, Clark C M, Naeem S, Pan Q M, Huang J H, Zhang L X, Han X G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands. Global Change Biology, 2010, 16(1): 358–372. DOI:10.1111/(ISSN)1365-2486 |
| [9] | 倪红伟, 郭亚胜, 吴海一. 小叶章种群分布格局的分形特征Ⅱ信息维数与关联维数. 植物研究, 2000, 20(3): 351–354. |
| [10] | 韩永利, 葛东宁, 张永喜. 三江平原湿地鸟类资源的现状及保护. 国土与自然资源研究, 2003(1): 76–76. |
| [11] | 刘吉平, 杨清, 吕宪国, 佟守正. 三江平原典型环型湿地生物多样性. 农村生态环境, 2005, 21(3): 1-5, 42-42. |
| [12] | 倪红伟, 张兴, 贾利, 高玉慧, 吴海一. 三江平原典型草甸小叶章种群地上生物量动态. 植物研究, 1998, 18(3): 328–335. |
| [13] | 倪红伟, 王宏韬, 李君霞, 宋玉玲, 马志军, 程岭, 王永吉. 三江平原小叶章种群生长速率的季节动态. 国土与自然资源研究, 1996(3): 67–71. |
| [14] | 孙志高, 刘景双, 王金达. 三江平原典型小叶章种群地上生物量的空间结构分形特征. 生态环境, 2006, 15(4): 775–780. |
| [15] | 杨继松, 刘景双, 于君宝, 王金达, 李新华, 孙志高. 三江平原沼泽湿地枯落物分解及其营养动态. 生态学报, 2006, 26(5): 1297–1302. |
| [16] | 王建波, 钟海秀, 付小玲, 王继丰, 倪红伟. 氮沉降对小叶章光合生理特性的影响. 中国农学通报, 2013, 29(7): 45–49. DOI:10.11924/j.issn.1000-6850.2012-3340 |
| [17] | Wang J B, Zhu T C, Ni H W, Zhong H X, Fu X L, Wang J F. Effects of elevated CO2 and nitrogen deposition on ecosystem carbon fluxes on the Sanjiang Plain wetland in Northeast China. PLoS One, 2013, 8(6): e66563. DOI:10.1371/journal.pone.0066563 |
| [18] | 秦胜金, 刘景双, 王国平, 王金达. 三江平原湿地土壤磷形态转化动态. 生态学报, 2007, 27(9): 3844–3851. |
| [19] | 李新华, 刘景双, 孙志高, 杨继松. 三江平原小叶章湿地生态系统硫的生物地球化学循环. 生态学报, 2007, 27(6): 2199–2207. |
| [20] | 李新华, 刘景双, 杨继松. 不同小叶章湿地H2S和COS的排放通量研究. 环境化学, 2007, 26(3): 284–289. |
| [21] | 李新华, 刘景双, 杨继松. 三江平原小叶章湿地H2S和COS排放动态. 环境科学, 2006, 27(11): 2145–2149. DOI:10.3321/j.issn:0250-3301.2006.11.001 |
| [22] | 施建敏, 马克明, 赵景柱, 王继丰. 三江平原残存湿地斑块特征及其对物种多样性的影响. 生态学报, 2010, 30(24): 6683–6690. |
| [23] | Sui X, Zhang R T, Yang L B, Zhong H X, Wang J F, Ni H W. Effects of long-term elevated CO2 fumigation on microbial communities in a wetland soil. Journal of Residuals Science & Technology, 2015, 12(S1): s93–s96. |
| [24] | 隋心, 张荣涛, 杨立宾, 许楠, 柴春荣, 王继丰, 付晓玲, 钟海秀, 邢军会, 张玉, 倪红伟. 模拟氮沉降对三江平原小叶章湿地土壤细菌多样性的影响. 草业科学, 2016, 33(4): 589–598. DOI:10.11829/j.issn.1001-0629.2015-0444 |
| [25] | 倪红伟, 李君. 洪河自然保护区生物多样性. 哈尔滨:黑龙江科学技术出版社, 1999: 1–2. |
| [26] | Oksanen J, Blanchet F G, Kindt R, Legendre P, O, Hara R B, Simpson G L, Solymos P, Stevens M H R, Wagner H. Vegan:community ecology package version 1.17-9. 2011, http://vegan.r-forge.r-project.org/. |
| [27] | Roberts D W. labdsv: Laboratory for Dynamic Synthetic Vegephenomenology. R Package Version 1.2-2. 2006, http://cran_r-project.org/. |
| [28] | 娄彦景, 赵魁义, 马克平. 洪河自然保护区典型湿地植物群落组成及物种多样性梯度变化. 生态学报, 2007, 27(9): 3883–3891. |
| [29] | 钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科技出版社, 1994: 141–160. |
| [30] | Barbault R. Biodiversity dynamics: from population and community ecology approaches to a landscape ecology point of view. Landscape and Urban Planning, 1995, 31(1/3): 89–98. |
| [31] | Whitehouse H E, Bayley S E. Vegetation patterns and biodiversity of peatland plant communities surrounding mid-boreal wetland ponds in Alberta, Canada. Canadian Journal of Botany, 2005, 83(6): 621–637. DOI:10.1139/b05-034 |
| [32] | 陈文业, 赵明, 张继强, 袁海峰, 窦英杰, 朱丽, 陈旭. 甘肃敦煌西湖荒漠-湿地生态系统土壤水分含量对植被特征的影响. 林业科学, 2015, 51(11): 8–16. |
| [33] | Hector A, Schmid B, Beierkuhnlein C, Caldeira M C, Diemer M, Dimitrakopoulos P G, Finn J A, Freitas H, Giller P S, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen A, Körner C, Leadley P W, Loreau M, Minns A, Mulder C P H, O'Donovan G, Otway S J, Pereira J S, Prinz A, Read D J, Scherer-Lorenzen M, Schulze E D, Siamantziouras A S D, Spehn E M, Terry A C, Troumbis A Y, Woodward F I, Yachi S, Lawton J H. Plant diversity and productivity experiments in European grasslands. Science, 1999, 286(5442): 1123–1127. DOI:10.1126/science.286.5442.1123 |
| [34] | 杜茜, 沈海亮, 王季槐. 宁夏荒漠草原植物群落结构和物种多样性研究. 生态学杂志, 2006, 25(2): 222–224. |
| [35] | Zhu D G, Ni H W. Compositional features and their relationships between soil seed bank and vegetation of Deyeuxia angustifolia community in Sanjiang Plain//Green Energy and Environmental Sustainable Development. 2011 International Conference on Green Energy and Environmental Sustainable Development. Jilin: GEESD, 2011: 1606-1610. |
| [36] | 江明喜, 邓红兵, 唐涛, 蔡庆华. 香溪河流域河岸带植物群落物种丰富度格局. 生态学报, 2002, 22(5): 629–635. |
| [37] | 张继强, 陈文业, 康建军, 吴三雄, 袁海峰, 王志广, 陈徵尼. 敦煌西湖盐化草甸芦苇植物群落特征及多样性沿水分梯度的分布格局. 水土保持通报, 2013, 33(2): 173–176. |
| [38] | Moeslund J E, Arge L, Bøcher P K, Dalgaard T, Ejrnæs R, Odgaard M V, Svenning J C. Topographically controlled soil moisture drives plant diversity patterns within grasslands. Biodiversity Conservation, 2013, 22(10): 2151–2166. DOI:10.1007/s10531-013-0442-3 |
| [39] | Vellend M. Do commonly used indices of β-diversity measure species turnover?. Journal of Vegetation Science, 2001, 12(4): 545–552. DOI:10.2307/3237006 |
| [40] | 邢福, 吕宪国, 倪红伟, 高玉慧, 许坤, 于丽丽, 张亮. 三江平原沼泽湿地群落演替系列β多样性. 应用生态学报, 2008, 19(11): 2455–2459. |
| [41] | 卢涛, 马克明, 倪红伟, 傅伯杰, 张洁瑜, 陆琦. 三江平原不同强度干扰下湿地植物群落的物种组成和多样性变化. 生态学报, 2008, 28(5): 1893–1900. |
| [42] | Miura A, Urabe J. Spatial and seasonal changes in species diversity of epilithic fungi along environmental gradients of a river. Freshwater Biology, 2015, 60(4): 673–685. DOI:10.1111/fwb.2015.60.issue-4 |
| [43] | Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213–251. DOI:10.2307/1218190 |
| [44] | Lande R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos, 1996, 76(1): 5–13. DOI:10.2307/3545743 |