 2017, Vol. 37
2017, Vol. 37文章信息
- 黄荣珍, 朱丽琴, 王赫, 贾龙, 刘勇, 段洪浪, 吴建平, 刘文飞
- HUANG Rongzhen, ZHU Liqin, WANG He, JIA Long, LIU Yong, DUAN Honglang, WU Jianping, LIU Wenfei.
- 红壤退化地森林恢复后土壤有机碳对土壤水库库容的影响
- Effects of soil organic carbon on soil reservoir capacity after forest restoration in degraded red soil
- 生态学报[J]. 2017, 37(1): 238-248
- Acta Ecologica Sinica[J]. 2017, 37(1): 238-248
- http://dx.doi.org/10.5846/stxb201608041609
-
文章历史
- 收稿日期: 2016-08-04
- 修订日期: 2016-11-04

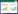




植被恢复过程是与土壤环境相适应的过程[1], 其显著特征表现在凋落物腐化后归还土壤、植被根系生长穿插及土壤微生物活动等, 不仅能够向土壤提供新的碳源, 而且能够改变土壤的孔隙状况, 增加土壤的通气性能及透水性能, 从而改善土壤环境, 提高土壤肥力[2];土壤结构和功能的改善反过来将促进生物多样性的形成、植被的加速生长。然而在我国南方红壤侵蚀区, 侵蚀退化恢复地虽然植被覆盖率和多样性有了很大的提升, 在暴雨季节依然面临山洪频发的威胁, 这在2016年夏季得到充分体现。究其原因, 很多侵蚀地虽然植被得以快速恢复, 但土壤包括土壤水库的生态功能未能得到同步的恢复。加上降雨量大而集中, 且暴雨频繁, 降雨无法及时入渗, 从而形成超渗地表径流、造成强烈土壤冲刷。与华北潮土、东北黑土、西北黄土相比, 红壤由于本身黏粒含量较高, 其防洪库容和有效库容大约仅为它们的一半, 使红壤更容易受到干旱威胁[3]。已有研究发现土壤有机碳含量的提高可改善土壤的持水性能[4], 土壤有机质的积累构成了土壤水分环境恢复和改善的主导因素[1]。通过植被恢复, 土壤有机碳增加, 土壤结构及其水力特性得到改善, 土壤水库蓄水供水调水及抗侵蚀功能得到提高。
目前, 国内外学者对退化地有机碳恢复、土壤水分物理性状的研究不少[5-6], 且有一些影响深远的研究成果, 但是, 对于退化地有机碳与土壤水库容的相关关系研究很少, 尤其是有机碳不同组分和土壤水库各种库容之间的实证定量研究更少。由于土壤水库容和退化地有机碳均表现为多个维度因素, 应用简单相关分析方法无法满足多维度变量之间相关分析的要求, 而典型相关分析模型却能分析两组变量之间的相关性。基于此点, 本文分别选取死库容、兴利库容和防洪库容作为土壤水库容的变量, 土壤总有机碳、微生物量碳、水溶性有机碳和易氧化有机碳作为土壤有机碳的变量, 应用SAS分析软件通过典型相关分析模型来分析退化地森林恢复后土壤有机碳和土壤水库容之间的相关性, 为从土壤有机碳-土壤结构-土壤恢复力角度理解土壤结构与功能相互作用提供新的认识, 从而有助于植被-土壤-水分环境互动关系及演变机理的深入研究和探讨, 并为南方严重红壤侵蚀退化区植被恢复树种选择、搭配和水土保持生态恢复实践提供科学依据。
1 材料与方法 1.1 试验地概况试验地设在中国科学院江西省千烟洲红壤丘陵综合开发试验站(115°04′13″E, 26°44′48″N), 属典型红壤丘陵地貌, 海拔高度在100 m左右, 相对高差在20-50m。土壤类型为红壤, 成土母质为红色砂岩。该区年均气温为17.9 ℃, 年均降水量为1489mm, 年均日照为1406h, 无霜期为323d, 属典型亚热带湿润季风气候。该站的原生植被以中亚热带常绿阔叶林为主, 由于人类长期开发和破坏, 原生植被遭到严重破坏, 在建站初期已退化为灌草丛, 且多处出现不同程度的水土流失, 现土地利用类型以人工林为主。各试验地基本情况见表 1。
| 森林恢复类型 Forest restoration type |
栽植时间 Planting time |
坡度 Slop/(°) |
坡向 Aspect |
保留密度 Reserved density/(株/hm2) |
平均树高/平均胸径 Average tree height/m /Average DBH/cm |
乔木郁闭度 Tree crown density/% |
灌木层盖度 Shrub coverage/% |
| 阔叶混交林(BF) | 1984 | 20 | 北向 | 889 | 18.9/15.8 | 90 | 45 |
| 木荷与马尾松混交林(SP) | 1984 | 15 | 西北向 | 1027(木荷) 753(马尾松) |
20.2/15.6(木荷) 22.0/19.8(马尾松) |
85 | 15 |
| 马尾松与阔叶复层林(PB) | 1984 | 15 | 南向 | 1025(马尾松) 850(阔叶乔木) |
18.9/18.0(马尾松) 10.4/8.0(阔叶乔木) |
75 | 60 |
| PB: Pinus massoniana and broad-leaved tree multiple layer forest, SP: Schima superba-Pinus massoniana mixed forest, BF: broad-leaved mixed forest | |||||||
阔叶混交林(BF, 对照):主要乔木树种为樟树(Cinnamomum camphora)、枫香(Liquidambar formosana)、鹅掌楸(Liriodendron chinense)、厚朴(Magnolia officinalis)、青冈栎(Cyclobalanopsis glauca)。林下植被主要有黄瑞木(Adinandra millettii)、石斑木(Rhaphiolepis indica)、香椿(Toona sinensis)、秤星树(Llex asprella)、三叶赤楠(Syzygium grijsii)、深山含笑(Michelia maudiae)、桂花(Osmanthus fragrans)、檵木(Loropetalun Chinense)、朱砂根(Ardisia crenata)、麦冬(Ophiopogon japonicus)、淡竹叶(Lophatherum gracile Brongn)、芒萁(Dicranopteris dichotoma)、狗脊蕨(Woodwardia japonica)、鳞毛蕨(Dryopteris)、铁线蕨(Adiantum capillus-veneris)等。
木荷与马尾松混交林(SP):主要乔木树种为木荷(Schima superba)和马尾松(Pinus massoniana)。林下植被主要有黄瑞木、青冈栎、白栎(Quercus fabri)、檵木、深山含笑、六月雪(Serissa japonica)、淡竹叶、麦冬、海金沙(Lygodium japonicum)、
马尾松与阔叶树复层林(PB):主要乔木树种为马尾松、黄瑞木、枫香、三角槭(Acer buergerianum)、白栎。林下植被主要有三叶赤楠、乌饭树(Vaccinium bracteatum)、檵木、秤星树、淡竹叶、芒萁、菝葜(Smilacis Chinae Rhizoma)、狗脊蕨等。
1.2 土壤采集与测定方法 1.2.1 土壤采集2014年4月, 在BF、SP和PB 3种林地分别设立3块20m×20m固定标准地, 共9块。在每个标准地内按“S”型布设5个取样点, 每个取样点开挖土壤剖面, 用环刀按0-10 cm、10-20 cm、20-40 cm、40-60 cm四个层次采集原状土样(分析时取0-10 cm与10-20 cm平均值作为0-20 cm的数值, 有机碳的数据类同), 每个层次分别用环刀取3个样, 用来测定土壤饱和持水量、毛管持水量和土壤容重。另用自封袋采集对应土壤层次土样, 用孔径2mm的土壤筛分离出直径大于2mm的石砾, 小于2mm的土样用四分法分成两份, 其中一份鲜土保存在4℃冰箱中, 用于微生物量碳、水溶性有机碳测定使用, 另一份风干研磨过筛, 作为凋萎持水量、总有机碳、易氧化有机碳测定使用。
1.2.2 测定方法土壤饱和持水量、毛管持水量和土壤容重采用环刀法测定:将装有湿土的环刀, 揭去上底盖, 保留一垫有滤纸的带网眼底盖, 放入平底盆中, 注入并保持盆中水层高度至环刀上沿不淹没为止, 使其吸水12h, 水平取出, 立即称重;接着放置在铺有干砂的平底盘中2h, 再次称重;然后将环刀土样放在105℃烘箱内烘干至恒重, 称量烘干土及环刀重量, 从而分别计算土壤饱和持水量、毛管持水量和土壤容重。土壤凋萎持水量采用饱和K2SO4法测定[7]。土壤总有机碳(SOC)采用K2Cr2O7氧化-外加热法测定。土壤微生物量碳(MBC)采用氯仿熏蒸-硫酸钾浸提[8], 并用碳氮分析仪(multi N/C 3100)测定浸提液有机碳浓度。土壤水溶性有机碳(WSOC)根据Chantigny[9]等方法, 稍作修改:首先称取鲜土10g (过2mm筛), 按土水比1:10加入100mL去离子水, 封口后在25℃恒温振荡器上以200 r/min振荡30min, 然后用中速定量滤纸过滤, 滤液以4000 r/min离心10min, 取上清液用0.45μm滤膜进行抽滤, 最后用碳氮分析仪(multi N/C 3100)测定滤液相应浓度。土壤易氧化有机碳(ROC)采用KMnO4氧化比色法测定[10]。
1.3 数据计算(1)土壤水库各库容计算[11]
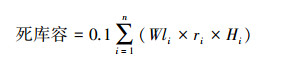
|
(1) |

|
(2) |
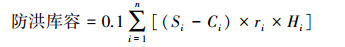
|
(3) |
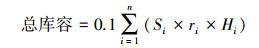
|
(4) |
式中,Wli为第i层土壤凋萎持水量(%);ri为第i层土壤容重(g/cm3);Hi为第i层土壤厚度(cm);Ci为第i层土壤毛管持水量(%);Si为第i层土壤饱和持水量(%);n为土壤层次。
(2)土壤有机碳密度计算[12]

|
(5) |
式中, SOCi为第i层土壤有机碳密度(kg/m2);Ti为第i层的土壤有机碳含量(g/kg);ri为第i层的土壤容重(g/cm3);Hi为第i层的土壤厚度(cm);Di为直径大于2mm的石砾所占的体积百分比(%);n为土壤层次。
1.4 数据处理所有数据为各重复实测值的平均值, 数据统计及采用Microsoft Excel 2003进行, 典型相关分析采用SAS 8.1软件进行, 制图采用PowerPoint 2003进行。
2 结果与分析 2.1 土壤水库各库容状况土壤水库死库容即土壤凋萎持水量所对应的库容, 其蓄持的水分无法被植物吸收利用, 也不能从土壤中释放, 保持相对稳定, 对径流调节不起作用。由表 2可见, 3种森林恢复类型土壤水库死库容均随土层深度的增加而增大, 其中BF、SP 20-40 cm、40-60 cm的死库容显著大于表层0-20 cm (P<0.05), 但两类型的20-40 cm与40-60 cm之间, 以及PB的各个土壤层次之间的死库容不存在显著性差异(P>0.05)。BF剖面0-60 cm总死库容显著大于PB (P<0.05)。
| 森林恢复类型 Forest restoration type |
土层深度/cm Soil depth |
死库容/mm Invalid |
兴利库容/mm Storage capacity |
防洪库容/mm Flood control capacity |
总库容/mm Total capacity |
| 阔叶混交林(BF) | 0-20 | 11.70±1.41Ba | 51.82±1.19Aab | 6.32±3.29Ab | 69.84±2.40Aa |
| 20-40 | 14.60±1.44Aa | 41.80±3.24Ba | 6.74±1.34Ab | 63.14±1.86Ba | |
| 40-60 | 16.83±0.41Aa | 39.27±2.78Ba | 6.32±2.35Ab | 62.42±1.12Ba | |
| 0-60 | 43.13±1.51a | 132.89±3.41a | 19.39±6.45b | 195.41a | |
| 木荷与马尾松 | 0-20 | 7.58±1.60Bb | 54.03±2.41Aa | 9.63±2.57Aab | 71.24±0.96Aa |
| 混交林(SP) | 20-40 | 13.00±1.15ABab | 45.22±2.05Ba | 6.96±1.47Ab | 65.18±0.60Ba |
| 40-60 | 15.29±4.33Aa | 40.72±2.77Ba | 7.27±2.56Ab | 63.28±0.92Ba | |
| 0-60 | 35.87±5.52ab | 139.97±5.26a | 23.86±6.39b | 199.70a | |
| 马尾松与阔叶树 | 0-20 | 8.79±1.75Aab | 43.82±7.43Ab | 12.72±2.26Aa | 65.33±2.01Ab |
| 复层林(PB) | 20-40 | 10.12±2.28Ab | 41.50±5.74Aa | 13.69±1.74Aa | 65.31±2.10Aa |
| 40-60 | 11.57±3.44Aa | 38.60±8.22Aa | 14.44±2.66Aa | 64.61±1.22Aa | |
| 0-60 | 30.48±7.46b | 123.93±21.19a | 40.86±1.04a | 195.27a | |
| 表中数据表示平均值±标准差, 不同大写字母表示土壤层次间差异达显著水平(P < 0.05), 不同小写字母表示森林恢复类型之间差异达显著水平(P < 0.05) | |||||
土壤水库兴利库容是对应于毛管持水量与凋萎持水量之间的贮水库容, 表征土壤较长时间贮存水分的能力, 其大小在中小尺度上对消减洪峰流量具有重要意义。3种森林恢复类型0-60 cm土层兴利库容大小顺序依次为SP>BF>PB, 三者之间不存在显著性差异(表 2), 表明SP土壤所能贮存的水量更多, 调节洪峰流量的潜力更大。3种类型土壤水库兴利库容均以0-20 cm土层最大, 随土层深度的增加而降低, 其中BF、SP表层0-20 cm兴利库容显著大于20-40 cm、40-60 cm (P<0.05)。
土壤水库防洪库容是指土壤剖面中非毛管孔隙的容量, 表征土壤暂时贮存水分的能力, 也是雨水进入土壤并补充地下水的主要通道[11]。3种恢复类型0-60 cm土层防洪库容以PB最大(表 1), 显著大于SP和BF (P<0.05), 防洪库容在土壤剖面不同层次中的变化规律不明显, 但每个层次PB的防洪库容显著大于SP、BF的(P<0.05)。
土壤水库总库容是指土壤剖面中所有孔隙的容量, 表征土壤所能容蓄水分的总量, 由死库容、兴利库容和防洪库容组成。0-60 cm土层总库容表现SP>BF>PB, 但相互之间没有显著差异(P>0.05)。从土壤剖面不同层次看, BF、SP表层0-20 cm总库容显著大于20-40 cm、40-60 cm (P<0.05), 但PB的不同层次之间不存在显著差异。
2.2 土壤有机碳和活性有机碳密度分布不同森林恢复类型下, 同一土层土壤总有机碳(SOC)密度均表现出PB>SP>BF (表 3);在0-20 cm土层, BF略小于SP且显著小于PB (P<0.05);在20-40 cm土层, BF和SP、PB土壤SOC密度相互之间存在显著性差异(P<0.05), 在40-60 cm土层, 表现为SP、PB显著大于BF (P<0.05)、SP与PB之间无显著差异(P>0.05)。在0-60 cm整个土壤厚度, 3种恢复类型SOC密度在5.794-8.717kg/m2之间, BF显著小于SP和PB (P<0.05)。随着土层深度的增加, 各类型土壤SOC密度均呈下降趋势, BF、SP和PB均呈现出0-20 cm土层土壤SOC密度显著大于20-40 cm、40-60 cm土层(P<0.05);同时, 除SP外, BF和PB 20-40 cm土壤SOC密度都显著大于40-60 cm土层, 显示出土壤SOC密度在>20 cm土壤深度急剧缩小, 且在20-60 cm深度变化幅度减小。
| 森林恢复类型 Forest restoration type |
土层深度 Soil depth/cm |
SOC密度 Density of SOC/ (kg/m2) |
MBC密度 Density of MBC/ (kg/m2) |
WSOC密度 Density of WSOC/ (kg/m2) |
ROC密度 Density of ROC/ (kg/m2) |
| 阔叶混交林(BF) | 0-20 | 3.189±0.324Ab | 0.060±0.020Aa | 0.195±0.022Ab | 1.853±0.343Aa |
| 20-40 | 1.504±0.227Bc | 0.024±0.008Ba | 0.186±0.003Ab | 0.717±0.254Ba | |
| 40-60 | 1.102±0.301Cb | 0.018±0.006Ba | 0.176±0.014Ab | 0.374±0.141Ca | |
| 0-60 | 5.794±0.349c | 0.103±0.032a | 0.557±0.037b | 2.944±0.720a | |
| 木荷与马尾松 | 0-20 | 3.697±0.266Ab | 0.031±0.008Ab | 0.242±0.016Aa | 1.010±0.066Ab |
| 混交林(SP) | 20-40 | 2.105±0.255Bb | 0.014±0.003Bb | 0.245±0.005Aa | 0.470±0.087Bb |
| 40-60 | 1.753±0.204Ba | 0.012±0.05Bb | 0.244±0.020Aa | 0.352±0.060Ca | |
| 0-60 | 7.554±0.575b | 0.058±0.016b | 0.731±0.037a | 1.832±0.200b | |
| 马尾松与阔叶 | 0-20 | 4.278±0.366Aa | 0.033±0.004Ab | 0.178±0.014Ab | 0.852±0.068Ab |
| 树复层林(PB) | 20-40 | 2.584±0.063Ba | 0.016±0.003Bb | 0.172±0.029Ab | 0.423±0.105Bb |
| 40-60 | 1.856±0.115Ca | 0.017±0.003Bab | 0.173±0.037Ab | 0.293±0.065Ca | |
| 0-60 | 8.717±0.539a | 0.066±0.009ab | 0.523±0.074b | 1.569±0.215b | |
| 表中数据表示平均值±标准差, 不同大写字母表示土壤层次间差异达显著水平(P < 0.05), 不同小写字母表示森林恢复类型间差异达显著水平(P < 0.05) | |||||
土壤微生物量碳(MBC)是反映土壤微生物活性强弱和有机质分解过程的重要生物学指标, 表征土壤质量的总体状况。由表 3可以看出, 3种类型同一土层土壤MBC密度大小顺序均表现为BF>PB>SP, BF在0-20 cm、20-40 cm土层均显著大于SP和PB (P<0.05)。随着土层深度的增大, 各类型土壤MBC密度总体成下降趋势, 下降的幅度在上层土壤较为明显;BF、SP和PB 20-40 cm土层MBC含量分别比0-20 cm土层下降了59.57%、54.70%和49.98%。
土壤水溶性有机碳(WSOC)是微生物自身生长及生物分解过程中的主要物质和能量来源[13], 3种森林恢复类型同一土层土壤WSOC密度大小顺序均表现为SP>BF>PB (表 3), SP显著高于BF、PB (P<0.05), BF与PB之间无显著性差异(P>0.05)。随着土层深度的增加, 土壤WSOC密度变化趋势不明显, 各层之间变化幅度不大。BF 0-60 cm土层土壤WSOC密度显著小于SP (P<0.05), 而略微大于PB。
易氧化有机碳(ROC)可反映土壤有机碳库的微小变化, 其与土壤有机碳呈显著正相关[14]。在同一土层深度下, BF土壤ROC密度最高(表 2), 在0-20 cm、20-40 cm土层显著大于SP、PB (P<0.05)。随着土层深度的增加, 各类型土壤ROC密度均呈逐渐降低的趋势, 相邻土层均存在显著差异(P<0.05)。BF 0-60 cm土层ROC密度显著大于SP、PB (P<0.05)。
2.3 土壤水库库容与土壤有机碳的典型相关分析本文将土壤SOC (x1)、MBC (x2)、WSOC (x3)和ROC密度(x4)作为原因组, 将土壤水库的死库容(y1)、兴利库容(y2)和防洪库容(y3)作为4个有机碳密度综合作用的结果组, 然后将两组指标做典型相关分析。土壤区分0-10 cm、10-20 cm、20-40 cm、40-60 cm四个层次对上述7个指标进行原因组和结果组的对应分析。
在0-60 cm土层, 第一对典型变量解释了86.61%的信息量, 第二对典型变量解释了11.55%的信息量, 根据对该两对典型变量进行Wilks′ Lambda、Pillai′s Trace、Hotelling-Lawler和Trace Roy′s Greatest Root等四种典型相关分析的总体检验, 显著性都小于0.001, 因此, 有机碳组与库容组之间存在着典型相关关系, 能够用有机碳数据解释库容数据。
2.3.1 相关系数矩阵原因组的x1、x2、x3和x4 4个指标两两之间的相关系数在-0.2131-0.4693之间(表 4), 结果组的y1、y2和y3 3个变量间的相关系数在0.1885-0.6233之间, 说明原因组4个变量之间、结果组的3个指标之间的相关程度均不高, 组内变量相互之间的信息重叠性不高, 因此, 典型相关分析可以将其一起分别作为原因组、结果组。
| 因子Factor | x1 | x2 | x3 | x4 | y1 | y2 | y3 |
| x1 | 1.0000 | 0.1524 | 0.0294 | 0.2565 | -0.1864 | -0.0125 | 0.2516 |
| x2 | 0.1524 | 1.0000 | -0.2131 | 0.4693 | -0.0585 | -0.1845 | -0.0768 |
| x3 | 0.0294 | -0.2131 | 1.0000 | -0.1910 | 0.6917 | 0.7867 | 0.2301 |
| x4 | 0.2565 | 0.4693 | -0.1910 | 1.0000 | -0.1419 | -0.1665 | -0.3198 |
| y1 | -0.1864 | -0.0585 | 0.6917 | -0.1419 | 1.0000 | 0.6233 | 0.1885 |
| y2 | -0.0125 | -0.1845 | 0.7867 | -0.1665 | 0.6233 | 1.0000 | 0.3234 |
| y3 | 0.2516 | -0.0768 | 0.2301 | -0.3198 | 0.1885 | 0.3234 | 1.0000 |
从有机碳组和库容组两两之间的相关系数来看, x3与y1、y2间的相关系数分别为0.6917和0.7867, 说明土壤WSOC密度的增加对土壤水库的死库容和兴利库容的增长具有明显的影响。对于土壤水库防洪库容来说, 土壤SOC密度影响最大, 相关系数为0.2516。
2.3.2 典型相关系数及检验由表 5可知, 第一对典型变量的典型相关系数为0.840, 通过显著性检验(P=0.01);第二对典型变量的典型相关系数为0.492, 通过显著性检验(P=0.01), 可利用其进行补充分析;第三对典型变量的典型相关系数为0.220, 没有通过显著性检验(P=0.01)。因此, 有机碳组与库容组之间存在典型相关关系, 能够用有机碳数据来解释库容数据, 可以用第一对和第二对典型变量来进行典型相关关系分析。
| 序号 Serail number |
典型关系系数 Canonical correlation coefficient |
相关系数的卡方统计值 Chi-SQ |
自由度 DF |
显著性 Sig. |
| 1 | 0.840 | 458.010 | 12.000 | 0.000 |
| 2 | 0.492 | 348.000 | 6.000 | 0.000 |
| 3 | 0.220 | 175.000 | 2.000 | 0.013 |
为了分析两组因子形成典型变量时相对作用的大小, 来自有机碳的原因组第一对典型变量为:

|
对于有机碳原因组的第一对典型变量, x3的系数最大, 为1.000, 因此可近似代表土壤有机碳水平。来自库容的结果组第一典型变量为:
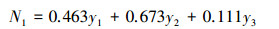
|
对于库容结果组的第一对典型变量, 兴利库容和死库容的系数相对较大, 两者联合可表示土壤水库库容状况。
第二对典型变量的典型相关系数为0.4918, 可利用其进行补充分析, 来自有机碳的原因组第二对典型变量为:

|
对于有机碳原因组的第二对典型变量, x1的系数最大, 为0.833, 表明其对土壤有机碳水平具有显著性影响;x4的系数的绝对值为0.848, 显示其反向即不易氧化的有机碳对土壤有机碳水平具有重要的影响。
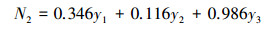
|
对于库容结果组的第二对典型变量, 防洪库容的系数高达0.986, 表明其对土壤水库库容状况具有极显著的影响。
2.3.4 典型结构分析结构分析基于因子在典型变量上的负载系数展开, 在第一对典型变量中(图 1), y1、y2、y3在库容结果组N1上的负载系数都是正数, 且y1和y2的负载系数较高, 分别为0.861和0.926, 因此, 3个库容变量与结果组第一对典型变量N1是正相关的, 且与y1、y2的相关程度很高。x3在原因组的第一对典型变量M1上的负载系数很高, 达0.981, 而x1、x2和x4的负载系数不高, 说明原因组第一对典型变量M1反映土壤的有机碳水平。

|
| 图 1 第一对典型变量的结构关系图 Fig. 1 Structure relation of the first pair of canonical variables |
利用第二对典型变量进行补充分析发现(图 2), y2、y3在库容结果组N2上的负载系数都是正数, 且y3的负载系数高达0.958, 因此, 这2个库容变量与结果组第二对典型变量N2是正相关的, 且与y3相关程度高。x1在原因组的第二对典型变量M2上的负载系数高达0.633, 而在x2、x3和x4上的负载系数不高, 说明原因组第二对典型变量M2也能一定程度上反映土壤的有机碳水平。

|
| 图 2 第二对典型变量的结构关系图 Fig. 2 Structure relation of the second pair of canonical variables |
本研究3种森林恢复类型0-20 cm、0-60 cm土层的总库容分别为65-71 mm和195-199 mm, 远小于东北黑土的120 mm和330 mm、华北潮土的108 mm和300 mm[15]、红壤区中龄林地的103-106 mm和307-347 mm、亚热带常绿阔叶天然林的126-130 mm和345-351 mm[16]。本研究的3种恢复林地前身水土流失严重, 表层原生疏松土壤遭受冲刷, 土壤有机碳损耗大, 土壤结构遭到严重破坏, 导致其土壤水库库容低下, 森林恢复后, 虽然水土流失和有机碳的矿化得到有效控制, 土壤有机碳状况有了大幅度的改善, 但恢复的时间相对较短, 要达到亚热带地带性常绿森林下的土壤结构和功能还需要漫长的时间。
东北黑土死库容、兴利库容和防洪库容占总库容的比例范围为30%-42%、40%-42%、17%-28%, 华北潮土的分别为30%-37%、33%-41%、22%-37%, 亚热带常绿阔叶天然林的分别为18%-25%、50%-51%、24%-32%[15, 16]。本研究中3种恢复森林土壤死库容、兴利库容和防洪库容占总库容的比例范围为13%-27%、61%-75%、7%-21%, 表明与黑土、潮土和天然林地红壤相比, 红壤恢复林地土壤水库不仅死库容、兴利库容、防洪库容与总库容小, 而且死库容和防洪库容占比小、兴利库容占比大。死库容占比较小可能与成土母质为红色砂岩有关, 其发育形成的土壤微细黏粒含量较第四纪红土发育的红壤低;防洪库容占比小、兴利库容占比大可能与红壤黏粒、Fe/Al氧化物含量高, 而有机质含量低有关, 导致由Fe、Al氧化物形成的微团聚体多、有机质形成的微团聚体及大团聚体相对较少。侵蚀退化恢复地土壤水库防洪库容占比较小的特性对于其透水理水、土壤结构与功能的恢复造成巨大的负面影响, 一方面不利于雨水的渗透和地表径流的削弱, 在雨强较大时无法及时渗透, 从而形成较大的地表径流和较强的土壤冲刷;另一方面土壤水分未能得到充分的补充, 导致土壤含水量经常处于亏缺状态, 影响林木根系水分吸收和土壤微生物活动, 不能快速有效培育土壤肥力, 同时也延滞恢复地森林生态系统的物质和能量循环, 使得土壤结构和功能恢复缓慢。
3.2 森林恢复对不同土壤有机碳的影响本研究森林恢复30a后3种森林0-20 cm、0-60 cm土壤总有机碳密度分别达31.89-42.78 t/hm2与57.94-87.17 t/hm2, 亚热带红壤严重退化地恢复24a马尾松林地(福建长汀)的为21.10 t/hm2、30.90 t/hm2[5], 亚热带不同演替阶段林地(湖南鹰嘴界自然保护区)的分别可达31.81-38.84 t/hm2、60.31-71.15 t/hm2[12], 亚热带常绿阔叶林分别可达61.69-77.27 t/hm2、143.66-180.30 t/hm2[17], 本研究结果大于严重侵蚀退化的马尾松恢复林地, 接近于不同演替阶段的马尾松林、马尾松阔叶树混交林, 远小于亚热带的地带性常绿阔叶天然林。土壤有机碳增加的幅度和速率取决于气候、土壤类型、植物类型和管理等因素[18]。本研究所在亚热带区域降雨与光照充沛、夏季高温多雨、雨热基本同季, 具备优越的植物生长和凋落物分解的气候条件, 有利于土壤有机碳的积累。3种森林类型选用适用于该区退化土地的水土保持先锋树种马尾松、木荷和枫香等品种, 具有成活率高、生长迅速、生产力高、凋落物丰富的特点。在造林过程中实施穴状整地、保留原有的植被, 抚育过程中将砍伐的灌木和草本用于林地覆盖, 而非传统的全垦、火烧整地和移除采伐剩余物, 减少了土地的干扰[19]、增加了凋落物对土壤有机碳的补充、降低了原有土壤有机碳的矿化[20]。
阔叶混交林MBC、WSOC、ROC占SOC的比例分别为1.78%、9.61%和50.81%, 木荷与马尾松混交林的分别为0.77%、9.68%和24.25%, 马尾松与阔叶树复层林的分别为0.76%、6.00%和18.00%;除了木荷与马尾松混交林的WSOC, 阔叶混交林其它类型活性有机碳密度及其占总有机碳的比例均大于马尾松与阔叶树复层林、木荷与马尾松混交林, 这与不同林分凋落物和细根的数量、性质及空间分布密切相关。土壤SOC和WSOC的主要来源是凋落物和细根, 不同植物类型凋落物、根系数量和质量不同, 导致其水溶性有机物、根际分泌物与微生物数量和质量相异[21], 而此3个因素对于土壤团聚体形成与稳定性提高具有显著影响[6, 22], 从而进一步对土壤水库的库容和功能发挥产生作用。
本研究中, 阔叶混交林的凋落物总量和凋落叶潜在有机碳归还量均显著低于木荷与马尾松混交林、马尾松与阔叶树复层林, 且木荷与马尾松混交林低于马尾松与阔叶树复层林, 与SOC的变化一致。凋落物通过微生物分解和淋溶等方式归还SOC和WSOC, 特别是新近凋落物可将大量WSOC释放到土壤中[23], 构成土壤WSOC最重要的“源”[24], 在相同气候条件和林龄下, 树种组成是影响森林凋落物产量与其有机碳的主要因素[25], 不同树种组成引起的凋落物组分差异即凋落物质量也会造成C归还差异, 其中落叶是凋落物C归还的主要组分, 这也使得亚热带森林中阔叶树的SOC和WSOC含量均比针叶树高[26]。本研究中阔叶混交林林分密度和生物量低于木荷与马尾松混交林、马尾松与阔叶树复层林, 但其落叶中阔叶量大、易于分解, 使得其SOC只略小于后两者。阔叶易于分解和快速归返的特性可为土壤微生物创造更有利的繁殖条件, 促进微生物活性和数量的增加[27], 微生物活动的加强又促进根系的代谢和分泌, 从而直接影响WSOC的增加[28]。相比较于SOC, WSOC活跃、矿化快, 可更快与土壤发生相互作用[28, 29], WSOC除了依赖于SOC含量外, 还取决于林地自身的微环境, 包括凋落物的数量和质量、根系状况及土壤理化性质等, 目前关于侵蚀地森林恢复后WSOC规模和动态还缺乏深入了解。
项目组前期研究发现细根生物量与SOC、WSOC显著相关, 且与WSOC相关系数大于SOC (未发表的数据), Ussiri等的研究也表明在矿山草地上恢复的森林细根生物量碳可导致SOC 62%的差异[30], 显示细根残渣和根际沉积提供的有机碳是SOC和WSOC的重要来源[31]。细根不仅影响着WSOC和SOC的数量, 还影响着两者的空间垂直分布。细根碳对SOC形成的贡献依赖于其生物量、周转速率、分泌物、菌根分布范围和土壤特征[32, 33]。因前期严重土壤侵蚀造成养分和SOC损失, 导致侵蚀退化地恢复后植被生产力低下, 凋落叶归还少[19], 细根就超过凋落物成为SOC、WSOC输入的重要渠道和主要来源[20], 特别是在土壤动物活动难以将源自凋落物叶的有机物质搬运到达的下层土壤中[21, 34], 根系不仅通过将土壤颗粒束绑在一起而增加团聚体, 同时也增加充当束绑介质作用的微生物[27]。因此, 选择细根生产力丰富的树木品种, 对于快速增加侵蚀退化地的土壤团聚体, 改善土壤水库库容状况具有至关重要的意义。
3.3 森林恢复后土壤有机碳对土壤水库库容的影响由图 1可知, 第一对典型变量间的相关系数达0.840, 显示每当土壤有机碳水平提高1%时, 土壤水库库容就会增加0.84%。这种作用体现了土壤有机碳水平对土壤水库库容的增加有显著的因果影响关系, 其中对有机碳水平起到主导性贡献作用的是水溶性有机碳密度。同时, 土壤的有机碳水平对土壤水库防洪库容的影响程度明显小于死库容和兴利库容。根据第一对典型变量结构关系, 若土壤有机碳水平提高1%, 死库容和兴利库容可分别增加0.72%和0.78%, 而防洪库容仅增加0.16%。本研究中第二对典型变量间的相关系数为0.492, 说明每当土壤有机碳水平提高1%时, 土壤水库库容就会增加0.49%(图 2)。这种作用体现了土壤有机碳水平对土壤水库库容的增加有较好的因果影响关系, 其中总有机碳对有机碳水平增加起到主要作用。其中土壤有机碳水平对土壤水库防洪库容的影响程度明显高于死库容和兴利库容。根据第二对典型变量结构关系, 若土壤有机碳水平提高1%, 兴利库容和防洪库容可分别增加0.11%和0.47%。
Wick等人的研究表明, 随着植被恢复有机质增加, 大团聚体增加、微团聚体下降, 即土壤大孔隙增加、小孔隙减少[6, 35]。而Dexter等人则认为随着有机质的增加, 微团聚体(小孔隙)也增加[36]。这可能与有机质的质量有关, 但目前对于有机质与团聚体关系的研究中只关注其数量, 对于其质量的影响研究不多[6, 36-37]。同时, 研究团聚体时, 大团聚体得到更多的关注和研究, 缺乏对微团聚体比较深入的了解[6, 36]。构成土壤水库库容的孔隙存在于不同大小团聚体之间和内部, 依据自然特性, 其等级的形成和团聚体等级相对应[38], 要达到促进植被恢复和提高森林理水调水功能的统一目的, 需要大、小团聚体都得到增加, 方可令防洪库容和兴利库容同时得以改善和提升, 达到防洪和蓄水、供水的多重目的。
4 结论本研究得出水溶性有机碳对于兴利库容、死库容具有显著的促进作用, 而总有机碳对于防洪库容具有明显的改善作用。马尾松与阔叶树复层林因林木保留密度大、凋落物量大、土壤有机碳密度大, 显现出较大的由大孔隙形成的防洪库容;而阔叶混交林、木荷与马尾松混交林中的阔叶树占比高、凋落物中阔叶比例大、土壤活性有机碳密度高, 表现出更高的由小孔隙形成的兴利库容、死库容。因此, 对于红壤退化地森林或植被恢复初期, 可通过适当密植和立体种植, 栽植凋落物量和细根生物量丰富的植物品种, 以提高生态系统生物量和土壤碳密度, 扩大土壤水库的防洪库容。同时, 可在马尾松等先锋树种针叶林分中补植阔叶乔灌木, 以增加土壤活性有机碳含量, 增大土壤水库兴利库容, 从而增强土壤的透水、蓄水、供水性能, 这对有效防治侵蚀区水土流失、提高土壤的水源涵养能力和减缓季节性干旱具有重要的实践意义。
| [1] | 赵世伟.黄土高原子午岭植被恢复下土壤有机碳-结构-水分环境演变特征[D].杨凌:西北农林科技大学, 2012. |
| [2] | Horn R, Smucker A. Structure formation and its consequences for gas and water transport in unsaturated arable and forest soils. Soil and Tillage Research , 2005, 82 (1) : 5–14. DOI:10.1016/j.still.2005.01.002 |
| [3] | 赵其国. 红壤物质循环及其调控. 北京: 科学出版社, 2002203–209. |
| [4] | 梁向锋.子午岭次生林区土壤物理特征对植被恢复的响应[D].陕西:中国科学院研究生院, 2008. |
| [5] | Xie J S, Guo J F, Yang Z J, Huang Z Q, Chen G S, Yang Y S. Rapid accumulation of carbon on severely eroded red soils through afforestation in subtropical China. Forest Ecology and Management , 2013, 300 : 53–59. DOI:10.1016/j.foreco.2012.06.038 |
| [6] | Wick A F, Ingram L J, Stahl P D. Aggregate and organic matter dynamics in reclaimed soils as indicated by stable carbon isotopes. Soil Biology and Biochemistry , 2009, 41 (2) : 201–209. DOI:10.1016/j.soilbio.2008.09.012 |
| [7] | 国家林业局. LY/T 1217-1999森林土壤稳定凋萎含水量的测定. 19990715 |
| [8] | 李振高, 骆永明, 滕应. 土壤与环境微生物研究法. 北京: 科学出版社, 2008. |
| [9] | Chantigny M H, Curtin D, Beare M H, Greenfield L G. Influence of temperature on water-extractable organic matter and ammonium production in mineral soils. Soil Science Society of America Journal , 2008, 74 (2) : 517–524. |
| [10] | Lefroy R D B, Blair G J, Strong W M. Changes in soil organic matter with cropping as measured by organic carbon fractions and 13C natural isotope abundance. Plant and Soil , 1993, 155-156 (1) : 399–402. DOI:10.1007/BF00025067 |
| [11] | 黄荣珍, 杨玉盛, 谢锦升, 王维明, 张金池. 福建闽江上游不同林地类型土壤水库"库容"的特性. 中国水土保持科学 , 2005, 3 (2) : 92–101. |
| [12] | 梁启鹏, 余新晓, 庞卓, 王琛, 吕锡芝. 不同林分土壤有机碳密度研究. 生态环境学报 , 2010, 19 (4) : 889–893. |
| [13] | 陈晓琳.中亚热带红壤丘陵区土地利用方式和景观位置对活性有机碳的影响研究[D].长沙:湖南大学, 2012. http://www.shangxueba.com/lunwen/v1514541.html |
| [14] | 林晓东, 漆智平, 唐树梅, 孟磊. 海南人工林地、人工草地土壤易氧化有机碳和轻组碳含量初探. 热带作物学报 , 2012, 33 (1) : 171–177. |
| [15] | 姚贤良. 红壤水问题及其管理. 土壤学报 , 1996, 33 (1) : 13–20. |
| [16] | 岳永杰.福建省主要森林水库特性与动态[D].福州:福建农林大学, 2003 http://www.docin.com/p-912869212.html |
| [17] | 周玉荣, 于振良, 赵士洞. 我国主要林林生态系统碳贮量和碳平衡. 植物生态学报 , 2000, 24 (5) : 518–522. |
| [18] | Lal R. Soil Carbon sequestration impacts on global climate change and food security. Science , 2004, 304 (5677) : 1623–1627. DOI:10.1126/science.1097396 |
| [19] | Yang Y S, Guo J F, Chen G S, Xie J S, Gao R, Li Z, Jin Z. Carbon and nitrogen pools in Chinese fir and evergreen broadleaved forests and changes associated with felling and burning in mid-subtropical China. Forest Ecology and Management , 2005, 216 (1/3) : 216–226. |
| [20] | Sheng H, Yang Y S, Yang Z J, Chen G S, Xie J S, Guo J F, Zou S G. The dynamic response of soil respiration to land-use changes in subtropical China. Global Change Biology , 2010, 16 (3) : 1107–1121. DOI:10.1111/gcb.2010.16.issue-3 |
| [21] | Malmer N, Svensson G, Wallén B. Carbon and mass balance in a south Swedish ombrotrophic bog:processes and variation during recent centuries. Mires and Peat , 2011, 8 (1) : 1–16. |
| [22] | Moreno-de las Heras M. Development of soil physical structure and biological functionality in mining spoils affected by soil erosion in a Mediterranean-Continental environment. Geoderma , 2009, 149 (3/4) : 249–256. |
| [23] | 王春阳, 周建斌, 夏志敏, 陈兴丽. 黄土高原区不同植物凋落物可溶性有机碳含量及其降解. 应用生态学报 , 2010, 21 (12) : 3001–3006. |
| [24] | Leff J W, Wieder W R, Taylor P G, Townsend A R, Nemergut D R, Grandy A S, Cleveland C C. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest. Global Change Biology , 2012, 18 (9) : 2969–2979. DOI:10.1111/j.1365-2486.2012.02749.x |
| [25] | 杨智杰, 陈光水, 谢锦升, 杨玉盛. 杉木、木荷纯林及其混交林凋落物量和碳归还量. 应用生态学报 , 2010, 21 (9) : 2235–2240. |
| [26] | Wang Q K, Wang S L. Soil organic matter under different forest types in Southern China. Geoderma , 2007, 142 (3/4) : 349–356. |
| [27] | Rasse D P, Rumpel C, Dignac M F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant and Soil , 2005, 269 (1/2) : 341–356. |
| [28] | Laik R, Kumar K, Das D K, Chaturvedi O P. Labile soil organic matter pools in a calciorthent after 18 years of afforestation by different plantations. Applied Soil Ecology , 2009, 42 (2) : 71–78. DOI:10.1016/j.apsoil.2009.02.004 |
| [29] | Gough C M, Flower C E, Vogel C S, Dragoni D, Curtis P S. Whole-ecosystem labile carbon production in a north temperate deciduous forest. Agricultural and Forest Meteorology , 2009, 149 (9) : 1531–1540. DOI:10.1016/j.agrformet.2009.04.006 |
| [30] | Ussiri D A N, Lal R, Jacinthe P A. Soil properties and carbon sequestration of afforested pastures in reclaimed minesoils of Ohio. Soil Science Society of America Journal , 2005, 70 (5) : 1797–1806. |
| [31] | Dannenmann M, Simon J, Gasche R, Holst J, Naumann P S, Kögel-Knabner I, Knicker H, Mayer H, Schloter M, Pena R, Polle A, Rennenberg H, Papen H. Tree girdling provides insight on the role of labile carbon in nitrogen partitioning between soil microorganisms and adult European beech. Soil Biology and Biochemistry , 2009, 41 (8) : 1622–1631. DOI:10.1016/j.soilbio.2009.04.024 |
| [32] | Yang Y S, Chen G S, Lin P, Xie J S, Guo J F. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in subtropical China. Annals of Forest Science , 2004, 61 (7) : 617–627. DOI:10.1051/forest:2004062 |
| [33] | Peichl M, Leava N A, Kiely G. Above-and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant and Soil , 2012, 350 (1/2) : 281–296. |
| [34] | Aumtong S, Magid J, Bruun S, De Neergaard A. Relating soil carbon fractions to land use in sloping uplands in northern Thailand. Agriculture, Ecosystems & Environment , 2009, 131 (3/4) : 229–239. |
| [35] | Wick A F, Stahl P D, Rana S, Ingram L J. Recovery of reclaimed soil structure and function in relation to plant community composition//Barnhisel R I, Ed. 2007 National Meeting of the American Society of Mining and Reclamation. Lexington, KY:ASMR, 2007, 40502:941-957. |
| [36] | Dexter A R, Richard G, Arrouays D, Czy E A, Jolivet C, Duval O. Complexed organic matter controls soil physical properties. Geoderma , 2008, 144 (3/4) : 620–627. |
| [37] | Tang J, Mo Y H, Zhang J Y, Zhang R D. Influence of biological aggregating agents associated with microbial population on soil aggregate stability. Applied Soil Ecology , 2011, 47 (3) : 153–159. DOI:10.1016/j.apsoil.2011.01.001 |
| [38] | McCarthy J F, Ilavsky J, Jastrow J D, Mayer L M, Perfect E, Zhuang J. Protection of Organic Carbon in soil microaggregates via restructuring of aggregate porosity and filling of pores with accumulating organic matter. Geochimica et Cosmochimica Acta , 2008, 72 (19) : 4725–4744. DOI:10.1016/j.gca.2008.06.015 |