 2017, Vol. 37
2017, Vol. 37文章信息
- 罗敏, 黄佳芳, 刘育秀, 仝川
- LUO Min, HUANG Jiafang, LIU Yuxiu, TONG Chuan.
- 根系活动对湿地植物根际铁异化还原的影响及机制研究进展
- Progress in effects of root bioturbation on dissimilatory iron reduction in the rhizosphere of wetland plants
- 生态学报[J]. 2017, 37(1): 156-166
- Acta Ecologica Sinica[J]. 2017, 37(1): 156-166
- http://dx.doi.org/10.5846/stxb201607261521
-
文章历史
- 收稿日期: 2016-07-26
- 修订日期: 2016-10-23





2. 福州大学环境与资源学院, 福州 350116;
3. 福建师范大学地理科学学院, 福州 350007;
4. 福建师范大学湿润亚热带生态地理-过程教育部重点实验室, 福州 350007
2. School of Environment and Resource, Fuzhou University, Fuzhou 350116, China;
3. School of Geographical Sciences, Fujian Normal University, Fuzhou 350007, China;
4. Key Laboratory of Humid Sub-tropical Eco-geographical Process of the Ministry of Education, Fujian Normal University, Fuzhou 350007, China
铁(Fe)是地壳中丰度第四的化学元素[1]。铁的正三价氧化物及氢氧化物水合物(Fe (Ⅲ))是湿地土壤中含量最丰富的电子受体之一[2-3]。铁异化还原是湿地常见的微生物呼吸方式, 尤其在湿地植物根际附近, 铁异化还原会抑制其他类型的微生物呼吸, 成为最重要的有机质代谢途径之一[4-11], 因此, 湿地植物根系活动可能是提高湿地植物根际铁异化还原速率的关键因素之一。
湿地根际铁异化还原过程不仅耦合了湿地生源元素碳、氮、磷、氧与硫的生物地球化学循环, 而且对温室气体的产生和氧化, 以及痕量金属铬、铅、铜、锌的吸附和解析, 都具有深远的影响[12]。系统、深入地开展湿地植物根系活动对植物根际的铁异化还原作用的研究, 可以更全面地了解湿地铁元素的迁移和转化机制, 同时也为当今根际生态学、土壤学、微生物学和植物学交叉领域的核心与重点内容--根系、土壤与微生物(Root-soil-microbe)三者间的物质和能量循环提供丰富的理论支持。
国际上对于湿地土壤铁异化还原的研究始于20世纪80年代[13], 陆续开展了淡水沼泽湿地[9-11]、盐沼湿地[5-8]、红树林[6-7]和裸滩[8]等不同湿地生境的铁异化还原研究。国内湿地铁异化还原的研究则始于本世纪初期, 研究对象包括水稻田[14-15]、淡水沼泽[16]、河口潮汐沼泽[17-19], 积累了一定的研究成果。相对于上述宏观尺度的铁异化还原研究, 关注湿地植物根际微域内铁异化还原的研究并不多, 更未见到相关的综述文献。鉴于此, 本文从湿地植物根际铁异化还原过程的电子受体、有机质底物、铁还原微生物和根际氧化还原环境入手, 梳理多年来国内外关于湿地根系活动对根际铁异化还原的影响及其机制的科研成果, 以期为未来湿地根际铁异化还原的深入研究提供参考。
1 根系活动和根际湿地植物的根系活动包括根系生长、吸收、分泌和死亡等多种生理现象, 其中根系泌氧和根系分泌有机物是湿地植物根系改造根际环境的重要手段[8]。根系泌氧是指湿地植物长期处于淹水环境中, 为满足根系呼吸的需求, 将光合作用产生的一部分氧输送至根系的生理现象[20-22]。根系分泌有机物是指活体根系以及死亡根系腐烂时向根际微域分泌可溶性有机物的生理现象[23-24]。根际是植物根系活动对周围土壤发挥作用的微环境。根系泌氧和根系分泌有机物的强度自根表开始逐渐向非根际土壤递减, 因此, 根际也是土壤有氧区和无氧区的界面[25]。
2 根际铁异化还原铁异化还原是指铁还原微生物利用外界的Fe (Ⅲ)作为电子受体, 氧化有机质底物, 将Fe (Ⅲ)还原成Fe (Ⅱ)的过程, 微生物在此过程中完成呼吸作用, 具体反应过程见下式[26]:
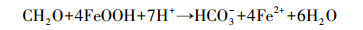
|
(1) |
国内外对湿地植物根际铁异化还原速率的研究主要集中在黑三棱草(Sparganium eurycarpum)、梭鱼草(Pontederia cordata)、宽叶香蒲(Typha latifolia)、互花米草(Spartina alterniflora)、大叶藻(Zostera marina)和灯心草(Juncus effusus)等植物(表 1)。美国马里兰州和弗吉尼亚州的宽叶香蒲根际铁异化速率为18 μmol g-1 d-1, 而非根际铁异化速率仅为8 μmol g-1 d-1; 10d内根际土壤中75%-80%的Fe (Ⅲ)可被还原, 而非根际土壤中只有30%-40%的Fe (Ⅲ)被还原[9]。美国华盛顿州的淡水沼泽野外原位培养实验中发现, 灯心草根系铁膜中95%的Fe (Ⅲ)在12个月内被还原, 铁异化还原速率的最大值可达0.6 mg Fe g-1 d-1干重, 而非根际的铁异化还原速率仅为0.05-0.1 mg g-1 d-1[27]。以上研究结果表明湿地植物根际土壤中铁异化还原的速率和潜力高于非根际土壤。通过比较美国缅因州淡水沼泽(梭鱼草、宽叶香蒲)和潮汐盐沼(互花米草、大叶藻)的铁异化还原速率[28], 可发现淡水沼泽(134-531 mg g-1 d-1)铁异化还原速率明显高于潮汐盐沼(16-83 mg g-1 d-1)。梭鱼草和宽叶香蒲之间、不同根型态(有根毛和无根毛)的梭鱼草之间的铁异化还原速率相差均超过100 mg g-1 d-1[28], 这表明不同种类的湿地植物及同一种类不同根型态间的根际铁异化还原速率相差较大。
| 湿地植物 Wetland plant species | 地点 Site | 湿地类别 Wetland types | 铁异化还原速率 Dissimilatory iron reduction rates/ (mg Fe g-1 d-1干重) |
| 黑三棱草Sparganium eurycarpum[28] | 美国缅因州 | 淡水沼泽 | 463±20 |
| 梭鱼草Pontederia cordata | |||
| 无根毛[28] | 美国缅因州 | 淡水沼泽 | 430±86 |
| 有根毛[28] | 美国缅因州 | 淡水沼泽 | 134±45 |
| 宽叶香蒲Typha latifolia | |||
| 无根毛[28] | 美国缅因州 | 淡水沼泽 | 531±72 |
| 有根毛[28] | 美国缅因州 | 淡水沼泽 | 372±67 |
| 有根毛[9] | 美国马里兰州、弗吉尼亚州 | 淡水沼泽、潮汐淡水沼泽 | 18±4 |
| 互花米草Spartina alterniflor[28] | 美国缅因州 | 潮汐盐沼 | 16±1 |
| 大叶藻Zostera marin[28] | 美国缅因州 | 潮汐盐沼 | 83±15 |
| 灯心草Juncus effusus[27] | 美国华盛顿 | 淡水沼泽 | 0.3±0.3 |
| 此处梭鱼草和宽叶香蒲按根型分为有根毛和无根毛两种根型态[28] | |||
根系活动对根际铁异化还原的影响, 可从电子受体、有机质底物、铁还原微生物、以及氧化还原电位和酸碱度4个方面发挥作用。
3.1 电子受体湿地植物根际铁异化还原的电子受体主要来自根系铁膜(Fe plaque)中大量活性的Fe (Ⅲ)[29-33]。铁膜是由于根系泌氧促使根际土壤中的二价铁(Fe (Ⅱ))被氧化, Fe (Ⅲ)在根系表面富集而形成的[20, 34-36]。目前, 已经在多种沉水植物、挺水植物、浮水植物和红树植物的根系表面发现了铁膜[22]。对大米草(Spartina anglica)、宽叶香蒲等植物的根际和非根际土壤中Fe (Ⅲ)的分析结果表明, 根际Fe (Ⅲ)的含量要比非根际土壤高出30-60 μmol g-1干重[8, 9, 37]。对灯心草和梭鱼草两种湿地植物根系铁膜的产生速率调查结果表明, 根际Fe (Ⅲ)产生速率达到0.33-0.40 mg g-1根d-1, 可为铁异化还原源源不断地补充电子受体[27]。在宽叶香蒲根际和非根际土壤中分别添加有机质、腐殖质以及铁还原微生物之后, 研究发现根际土壤的铁异化还原速率仍高于非根际土壤[9], 该结果说明根际Fe (Ⅲ)含量决定了铁异化还原速率[10, 38]。
结合传统和现代分析技术, 目前学界对湿地植物根际Fe (Ⅲ)的矿物形态已有了一定的认识。利用稀盐酸和草酸连续浸提法对根际和非根际土壤中Fe (Ⅲ)的形态进行研究, 结果表明植物根际土壤中约66%的Fe (Ⅲ)为无定形态, 约50%的Fe (Ⅲ)为晶质态; 而在非根际土壤中50%的Fe (Ⅲ)为无定形态, 约18%的Fe (Ⅲ)为晶质态[27]。通过扫描式电子显微镜(SEM)观察发现宽叶香蒲和芦苇(Phragmites australis)根系铁膜中Fe (Ⅲ)的主要成分为无定形态的水铁矿和纤铁矿[39]; 通过X射线衍射(XRD)和X射线吸收技术(XAFS)发现湿地植物铁膜中的Fe (Ⅲ)除了无定形态Fe (Ⅲ), 还包括部分结晶态铁Fe (Ⅲ), 如针铁矿、磁铁矿[34, 40]。无定形态Fe (Ⅲ)矿物的比表面积(Specific surface area)更大且吸附性(Affinity)更高, 会更加受到铁还原微生物的青睐[41]。
除了Fe (Ⅲ)的含量和形态, Fe (Ⅲ)矿物的异质性也会对铁异化还原能力产生重要的影响[42-43]。矿物异质性越高的Fe (Ⅲ)矿物组成越复杂, 铁还原微生物对其利用的效率越低。利用活性连续模型发现大米草根系铁膜中Fe (Ⅲ)的矿物异质性要远远低于非根际土壤中Fe (Ⅲ)的矿物异质性[9], 使得根际铁异化还原的速率要高于非根际。
3.2 有机质底物湿地植物通过光合作用固定的碳可通过根系分泌物的形式释放到土壤中, 为土壤微生物提供丰富的有机质底物[44-45]。这也是潮汐盐沼土壤溶解性有机碳(DOC)含量要远远高于裸滩土壤的重要原因[4]。目前学界对铁还原微生物利用的主要碳源还存在分歧, 一种观点认为死亡的根系是铁还原微生物呼吸的最大碳源[45], 另一种观点认为大部分死亡根系有机物相对难分解, 很难被微生物利用, 根系分泌的可溶性有机物才是铁还原微生物的主要碳源[44, 46]。有研究发现淡水湿地沉积物中铁异化还原速率和有机质底物的含量成正比[43]。
利用(UV-VIS)、红外光谱仪(IR)、毛细管电泳仪(CK)、气相色谱仪(GC)、液相色谱仪(LC)、离子色谱仪(HPIC)、质谱仪(MS)和核磁共振仪(NMR)分析以及色谱质谱联用技术(GC-MS、LC-MS), 已经发现湿地植物的根系分泌物包括糖类、氨基酸类、有机酸、酚酸类、脂肪酸、甾醇类、蛋白质与生长因子等[24]。有机物的C:N比可影响铁还原微生物。例如, 木质素的C:N比值较高, 不易被铁还原微生物利用[6], 而单糖、脂肪酸、蛋白质和核酸的C:N比较低, 更易被铁还原微生物利用[47]。根系分泌的氨基酸有可能发生聚合作用生成腐殖质, 腐殖质虽然难被分解, 但可作为电子传输体对铁异化还原产生重要的影响[48]。
3.3 铁还原微生物已有不少研究在湿地植物的根际分离出铁还原微生物:利用平板计数法发现根际铁还原微生物的相对丰度(-12%)远高于非根际土壤中铁还原微生物的相对丰度(0.5%)[38]; 有植物生长的湿地土壤铁还原微生物的丰度要远高于无植物的裸滩[8]。另外, 通过调查宽叶香蒲根际的铁还原微生物的丰度, 发现铁还原微生物的丰度和无定形Fe (Ⅲ)的含量呈正相关关系[9, 38]。
不同湿地植物根系分泌物中包含的有机底物、酶和生长因子也不尽相同, 将对根际铁还原微生物群落结构起到选择塑造作用[49-51]。研究发现, 在河口潮汐湿地互花米草和宽叶香蒲植物根际发现以醋酸、乙醇和琥珀酸作为有机质底物的地杆菌(Geobacter)[27, 38]。同样, 在湖泊湿地植物轮叶黑藻(Hydrilla verticillata)根际土壤中也发现了这种铁还原微生物[52]。利用变形梯度凝胶电泳(PCR-DGGE)和实时定量荧光PCR技术(Real-time PCR)发现水稻田根际富含地杆菌、孢杆菌(Bacillus)与梭菌(Clostridia)等[53]。利用16S rDNA法发现在滨海沼泽植物根际, 地杆菌和希瓦氏菌(Shewanella)等水稻田常见的铁还原微生物丰度较低, 而雷尔氏菌(Ralstonia)与梭状芽胞杆菌(Clostridium)这些较罕见的铁还原微生物的丰度较高[54]。
根际与非根际土壤中电子受体、有机质底物和氧化还原环境的不同, 也可能造成两者微生物群落结构的差异[23-24]。已有文献报道过红树林[55]、淡水湿地[50, 56]、水稻田和盐沼湿地[57]植物根际和非根际土壤总微生物、产甲烷微生物、反硝化微生物、硫酸根微生物群落结构的差异, 但目前尚未有研究分析根际与非根际土壤中铁还原微生物群落结构的差异。
3.4 氧化还原电位和酸碱度根系活动对根际周围的氧化还原电位(Eh)和酸碱度(pH)造成较大的改变, 并由此间接对铁异化还原过程产生影响。
植物根系通过根系泌氧作用, 使得根际土壤环境中的Eh水平高于非根际土壤环境[35, 36]。对巴西不同红树植被根际的氧化还原环境进行研究, 发现由于根系泌氧作用使得植物根际划分为Eh > 350 mV的氧化区和Eh介于100-350 mV的亚氧区, 非根际则处于厌氧环境(Eh < -100 mV), 而铁异化还原主要集中在根际的亚氧区[58, 59]。Kostka等[8]认为Eh高于200 mV时, 铁的异化还原过程受到抑制, Fe (Ⅲ)不会还原成Fe (Ⅱ); 而当Eh介于100-200mV时, 铁异化还原强度增大且土壤中的Fe (Ⅱ)浓度明显增高。上述研究结果都表明亚氧状态下, 铁还原微生物才会表现出较高的活性, 且铁异化还原的Eh临界值并不是一个固定值[60]。
根系分泌的有机酸会促使根际微域的pH降低[48], 增进根际Fe (Ⅲ)的溶解。例如, 根系释放的羧酸盐会使得根际土壤的pH比非根际土壤pH低1-2, 同时土壤中Fe2+的浓度升高[61]。也有研究认为在淹水环境下, 沼泽植物根际Eh < 400 mV且pH < 6时, Fe (Ⅲ)易被铁还原微生物利用而被还原为Fe (Ⅱ)[62]。
4 湿地植物根际铁异化还原与其他有机质代谢途径的竞争湿地植物根系活动还会影响根际好氧微生物, 以及硫酸盐还原微生物和产甲烷微生物等厌氧微生物, 从而影响根系铁还原微生物和这些微生物对有机质底物的竞争, 使得有机物的代谢途径发生改变。
4.1 与好氧微生物的竞争根系泌氧激发了好氧微生物的活性。由于有氧代谢的速率快, 且能够代谢大分子的有机质底物, 从而一定程度上抑制铁异化还原[63]。但是, 也有研究发现在富含Fe (Ⅲ)的红树林湿地, 有氧代谢对有机质矿化的贡献率仅为10%[6]。有观点认为这是由于大部分的氧并未参与有机质代谢, 而是氧化根际的还原性化学物, 如Fe (Ⅱ)、HS-及铁的硫化物[64]。
4.2 与硫酸盐还原微生物的竞争湿地植物根系分泌物量不足时, 将引起硫酸盐还原微生物与铁还原微生物争夺有机质底物[5, 46]。在滨海潮汐湿地沉积物中, 由于SO42-的高含量以及Fe (Ⅲ)的难溶性, 使得铁还原微生物在有机质底物竞争过程中常处于劣势, 从而使铁异化还原对有机质代谢的贡献率低于硫酸盐异化还原[5, 44, 46]。但也有研究发现当滨海沼泽沉积物中Fe (Ⅲ)的含量较高时, 二者的竞争优劣关系将发生转换。当滨海潮汐沼泽沉积物中Fe (Ⅲ)的含量大于20 μmol/cm3时, 铁异化还原可压制硫酸盐异化还原, 并完成根际区大于99%的有机质代谢[65, 66]。淡水湿地土壤的SO42-含量低, 植物根际铁异化还原超过硫酸盐异化还原成为最重要的有机质代谢途径[37]。植物根系高速的铁循环及铁还原微生物较高的腐殖质酸的代谢能力, 也是铁异化还原成为淡水湿地植物根系附近最重要的有机质代谢途径的重要原因[62]。
此外, 硫酸盐异化还原和铁异化还原还与沉积物的成分有关。若沉积物中矿物含量较高, 根际有机质的代谢途径以铁异化还原为主, 若在有机质含量较高的土壤中, 根际有机质的代谢途径则以硫酸盐异化还原为主[10]。
4.3 与产甲烷微生物的竞争在滨海湿地中, 铁还原微生物的竞争能力一般都高于产甲烷微生物。在泰国普吉岛滨海红树林湿地的研究中发现产甲烷微生物对有机质的贡献率小于1%, 但铁异化还原的贡献率可达73%-84%[6]。在中国闽江河口短叶茳芏湿地沉积物中的研究中也发现较高(20%-89%)的铁异化还原贡献率[19, 67]。
在淡水湿地中, 产甲烷微生物与铁还原微生物的竞争更加复杂。美国马里兰州Jug湾淡水沼泽植物根际土壤中铁异化还原对有机质代谢的贡献率为58%, 产甲烷过程对有机质代谢的贡献率为40%[10]。在中国湖北省水稻田根际土壤中添加葡萄糖, 发现铁异化还原有效地抑制了产甲烷过程[14]。根系泌氧作用源源不断为铁异化还原补充电子受体被认为是淡水沼泽植物根际高铁异化还原速率的关键[37]。
5 根系活动的异质性及其对根际铁异化还原的影响 5.1 根系活动的时间异质性根系活动的强度受到根系生长发育、光合作用和呼吸作用的调节[21, 68-70], 使得根系活动强度在昼夜和季节等时间尺度内发生波动[71, 72]。
春夏两季, 湿地植物的根系活动逐渐增强。伴随着根系铁膜的产生速率和根系分泌有机物的强度的增加, 淡水沼泽湿地植物根际铁异化还原的速率及其对有机质的贡献率也达到峰值; 冬季植物根系活动减弱, 铁异化还原的速率也随之减弱[10]。在滨海沼泽湿地中, 根际铁异化还原除了受根系活动的扰动, 还受到硫酸盐异化还原的影响[8, 73]。夏季硫酸盐异化还原速率达到最高, 大量Fe (Ⅲ)被H2S还原, 铁异化还原被抑制; 冬季硫酸盐异化还原对有机质代谢的贡献率降低, 铁异化还原速率增加[74]。
白天湿地植物以光合作用为主, 根系泌氧能力增加使得铁异化还原能力较强; 夜晚湿地植物以呼吸作用为主, 此时根系泌氧能力减弱, 铁异化还原能力也相应减弱[62]。对人工湿地的调查中, 研究发现芦苇、美人蕉、富贵竹等湿地植物根系泌氧呈现明显的日变化, 且根系泌氧强度与光合作用的速率成正相关, 说明根系泌氧的O2来源主要为光合作用产生的O2[75]。
5.2 根系活动的空间异质性根系活动在根际微域的变化也会影响铁异化还原的空间分布。利用微电极技术(Microelectrode)研究发现, 距离根系表面越远的位置根系泌氧的强度越弱, 且当距离根系大于5mm-2cm处, 根系扩散的氧气消耗殆尽[29, 35]。受根系泌氧和根际分泌有机物的双重影响, 铁还原微生物最活跃的位置并非在氧浓度最高的根系表面, 也不在完全厌氧的非根际土壤中, 而应位于根系表面和非根际土壤之间的亚氧环境中, 使得铁还原微生物既能适应其氧化还原环境, 又能有效地补充电子受体和有机质底物[62]。也就是说, 根际铁异化还原主要集中在受根系泌氧影响的几毫米或几厘米处的狭小空间[76]。
与上述观点不同的是, 在淡水沼泽植物灯心草根系表面发现了铁还原微生物[38], 这可能是由于湿地植物根系细胞的直径(10-20μm)和铁异化还原微生物的直径(0.4-0.9 μm)相差较大, 使得部分铁还原微生物附着在根系的泌氧屏障区表面[10]。另外, 湿地植物根基和根尖处的泌氧屏障发育不一致, 也可能会造成根基和根尖处泌氧强度差异, 从而影响铁还原微生物在根系上的空间分布[22, 77]。
5.3 根系活动的种间和种内异质性植物的根系活动受到通气组织[69, 78-79]、根密度[78]、泌氧屏障[78]和地下生物量[23]等因素的影响。通过水培实验研究菹草(Potamogeton crispus)和伊乐藻(Elodea nuttallii)两种植物根系铁膜时[80], 发现铁膜形成的差异主要是根的形态差异造成的, 伊乐藻的根粗, 但数量少, 因此其表面积小, 根表形成的铁膜量也相对较少, 而菹草根细且多, 其表面积大, 根表形成的铁膜量也较多。另有研究表明, 灯心草根表铁膜含量分别是茭白(Zizania latifolia)和美人蕉(Canna indica)的2.93倍和10-58倍[40]。芦竹(Arundo donax)和宽叶香蒲根系铁膜的量(以根系鲜重计)分别为20170.8 mg/kg和7640.3 mg/kg[41]。因此, 不同植物[78], 同一植物不同品种[28], 甚至不同基因组成的同种植物[62, 81], 其根系活动的强度都可能不一致。这有可能是造成湿地植物种内和种间铁异化还原的异质性的重要原因之一, 但其机理仍有待进一步研究。
5.4 环境因子造成的根系活动异质性温度、光照、淹水条件、土壤含水率、土壤氧化还原电位等环境因子的波动变化, 也将造成根系活动的异质性。温度的增加可增强湿地植物根系泌氧的能力。例如, 在对潜流型人工湿地的调查中发现, 芦苇、茭白、美人蕉和空心菜(Ipomoea aquatica)的泌氧能力受温度的影响, 夏季根系泌氧的能力高于冬季, 白天根系泌氧的能力要高于夜晚[75]。光照加强了湿地植物的蒸腾作用, 使得植物体内和大气的压强梯度增大, 从而促使O2快速通过通气组织到达根系, 增强湿地植物根系的泌氧能力[82]。例如, 水稻(Oryza sativa)随光照强度的増加, 根系泌氧呈明显的上升趋势[83]; 对狐尾藻(Myriophyllum spicatum)和菹草泌氧能力的研究发现根系泌氧在光照条件下比黑暗条件下大很多[84]。淹水条件和土壤含水率会影响根系铁膜的厚度[85]。长期生活在淹水条件下的水稻根系铁膜较厚; 而生长在干湿交替处理环境中的水稻铁膜则相对较薄[86]。在加拿大魁北克南部和安大略省的芦苇根系铁膜的厚度与土壤的含水率成正相关关系[87]。而对苦草(Vallisneria spiralis)的研究则表明土壤的氧化还原条件可以反过来影响根系泌氧的强度变化[88]。环境因子造成的根系活动异质性可能进一步作用于根际铁异化还原。
6 根际铁异化还原的概念模型湿地植物根际铁异化还原概念模型如图 1所示。根系活动为湿地植物根际输入O2和有机质, 根际微域内O2和有机质的浓度由根表开始逐渐向非根际区递减; 土壤中的Fe2+被根系泌氧释放的O2氧化, 并在根系表面积累大量Fe (Ⅲ); Fe (Ⅲ)在铁还原微生物的介导下被还原为Fe2+, 有机质被代谢生成二氧化碳和营养盐, 产生的Fe2+重新被根系泌氧释放的O2氧化成Fe (Ⅲ)。

根际铁异化还原和铁膜的形成是湿地植物根际铁循环重要组成部分[62]。若湿地植物的根表铁膜过厚, 将在根系表面形成泌氧屏障, 降低根系泌氧能力[89-90], 并影响根系活力及根系对元素的吸收和利用[70]。此外, 根系铁膜的形成过程可释放出H+, 降低根际pH值[62]。铁异化还原作用使得湿地根系的铁膜不会无止境的增厚[62, 65]。当铁膜厚度到达一定程度时, 铁膜中的Fe (Ⅲ)被还原成Fe2+, 同时释放出, 提高根际的pH, 同时降低根际氧化还原电位[9]。湿地植物根系也将逐渐恢复活力和泌氧能力, 促进对营养元素的吸收[62]。因此, 根际铁异化还原对促进根际有机质代谢分解, 维持湿地植物根系的健康生长、适应淹水环境、促进营养元素的吸收和维持根际酸碱平衡具有重要的生态意义[91]。
7 总结和展望根系活动是湿地铁元素生物地球化学循环的动力泵。综合国内外学者对根际铁异化还原的研究进展, 我们可归纳出:根系活动通过电子受体、有机质底物、铁还原微生物以及根际的氧化还原电位和酸碱度深刻影响铁异化还原过程。同时, 根系活动也会影响铁异化还原和其他有机质代谢途径的竞争关系。根系活动的时空波动变化、种内和种间差异性以及环境因子对根系活动的影响, 都有可能造成湿地植物根际铁异化还原的异质性。对于根际铁异化还原的研究虽已取得一定的进展, 但还有几个方面需要深入:
(1)加强我国滨海湿地植物根际铁异化还原的研究。目前国内对根际铁异化还原的研究大多侧重于水稻田和内陆湿地, 而滨海沼泽受潮汐的影响, 使得湿地植物的根际酸碱性、盐分、硫酸盐含量、淹水条件等和其他湿地生态系统相差较大, 环境更为复杂, 亟待深入研究。
(2)加强环境影响因子和全球变化对根际铁异化还原的研究。目前关于温度、湿度、盐分、淹水条件等环境因子对根际铁异化还原的影响研究较弱, 对气候变暖、氮沉降、硫沉降和外来物种入侵等全球变化影响下根际铁异化还原的变化也鲜见报道。
(3)结合植物根际活动的时空动态变化展开研究。目前的研究中并没有将铁异化还原的过程和湿地植物根系的生长特征(地下生物量、根系长度、根系通气组织、根密度、根系泌氧屏障等)以及生命活动周期(季节节律、日夜节律等)有机地结合起来, 因此, 仍无法从植物学视角把握根际铁异化还原特殊的时空动态特征。
(4)全面提升研究技术和手段。传统的测定方法已经不能满足根际铁异化还原的研究需求, 需结合和运用多领域、多学科的先进技术。例如, 利用微电极来测定根系泌氧的动态变化; 利用同位素法区别铁的异化还原和化学还原; 利用光谱、电泳、色谱和质谱等技术分析根系分泌物中有机物的种类和数量; 利用X射线衍射和X射线吸收等技术研究根际Fe (Ⅲ)的矿物形态, 利用实时定量荧光PCR技术、荧光原位杂交技术(FISH)从分子生物学水平上测定根际铁还原微生物的丰度和群落结构等。
| [1] | Weaver B L, Tarney J. Empirical approach to estimating the composition of the continental crust. Nature , 1984, 310 (5978) : 575–577. DOI:10.1038/310575a0 |
| [2] | Kostka J E, Luther Ⅲ G W. Partitioning and speciation of solid phase iron in saltmarsh sediments. Geochimica et Cosmochimica Acta , 1994, 58 (7) : 1701–1710. DOI:10.1016/0016-7037(94)90531-2 |
| [3] | Kostka J E, Luther Ⅲ G W. Seasonal cycling of Fe in saltmarsh sediments. Biogeochemistry , 1995, 29 (2) : 159–181. |
| [4] | Gribsholt B, Kristensen E. Benthic metabolism and sulfur cycling along an inundation gradient in a tidal Spartina anglica salt marsh. Limnology and Oceanography , 2003, 48 (6) : 2151–2162. DOI:10.4319/lo.2003.48.6.2151 |
| [5] | Hyun J H, Smith A C, Kostka J E. Relative contributions of sulfate-and iron (Ⅲ) reduction to organic matter mineralization and process controls in contrasting habitats of the Georgia saltmarsh. Applied Geochemistry , 2007, 22 (12) : 2637–2651. DOI:10.1016/j.apgeochem.2007.06.005 |
| [6] | Kristensen E, Andersen F, Holmboe N, Holmer M, Thongtham N. Carbon and nitrogen mineralization in sediments of the Bangrong mangrove area, Phuket, Thailand. Aquatic Microbial Ecology , 2000, 22 (2) : 199–213. |
| [7] | Kristensen E, Mangion P, Tang M, Flindt M R, Holmer M, Ulomi S. Microbial carbon oxidation rates and pathways in sediments of two Tanzanian mangrove forests. Biogeochemistry , 2011, 103 (1-3) : 143–158. DOI:10.1007/s10533-010-9453-2 |
| [8] | Kostka J E, Roychoudhury A, Van Cappellen P. Rates and controls of anaerobic microbial respiration across spatial and temporal gradients in saltmarsh sediments. Biogeochemistry , 2002, 60 (1) : 49–76. DOI:10.1023/A:1016525216426 |
| [9] | Weiss J V, Emerson D, Megonigal J P. Geochemical control of microbial Fe (Ⅲ) reduction potential in wetlands:comparison of the rhizosphere to non-rhizosphere soil. FEMS Microbiology Ecology , 2004, 48 (1) : 89–100. DOI:10.1016/j.femsec.2003.12.014 |
| [10] | Neubauer S C, Givler K, Valentine S, Megonigal J P. Seasonal patterns and plant-mediated controls of subsurface wetland biogeochemistry. Ecology , 2005, 86 (12) : 3334–3344. DOI:10.1890/04-1951 |
| [11] | Keller J K, Sutton-Grier A E, Bullock A L, Megonigal J P. Anaerobic metabolism in tidal freshwater wetlands:Ⅰ. Plant removal effects on iron reduction and methanogenesis. Estuaries and Coasts , 2013, 36 (3) : 457–470. DOI:10.1007/s12237-012-9527-6 |
| [12] | 罗敏, 曾从盛, 仝川, 黄佳芳. 滨海潮滩铁异化还原研究进展. 湿地科学 , 2014, 12 (4) : 527–532. |
| [13] | Lovley D R, Phillips E J. Organic matter mineralization with reduction of ferric iron in anaerobic sediments. Applied and Environmental Microbiology , 1986, 51 (4) : 683–689. |
| [14] | 曲东, 贺江舟, 孙丽蓉. 不同水稻土中氧化铁的微生物还原特征. 西北农林科技大学学报:自然科学版 , 2005, 33 (4) : 97–101. |
| [15] | 曲东, 张一平, SchnellS, ConradR. 水稻土中铁氧化物的厌氧还原及其对微生物过程的影响. 土壤学报 , 2004, 40 (6) : 858–863. |
| [16] | 姜明, 吕宪国, 杨青, 佟守正. 湿地铁的生物地球化学循环及其环境效应. 土壤学报 , 2006, 43 (3) : 493–499. |
| [17] | Luo M, Zeng C S, Tong C, Huang J F, Yu Q, Guo Y B, Wang S H. Abundance and speciation of iron across a subtropical tidal marsh of the Min River Estuary in the East China Sea. Applied Geochemistry , 2014, 45 : 1–13. DOI:10.1016/j.apgeochem.2014.02.014 |
| [18] | Luo M, Zeng C S, Tong C, Huang J F, Yu Q, Guo Y B, Wang S H. Kinetics of chemical and microbial iron reduction along an inundation gradient in a tidal marsh of the Min River Estuary, Southeastern China. Geomicrobiology Journal , 2015, 32 (7) : 635–647. DOI:10.1080/01490451.2014.950362 |
| [19] | Luo M, Zeng C S, Tong C, Huang J F, Chen K, Liu F Q. Iron reduction along an inundation gradient in a tidal sedge (Cyperus malaccensis) marsh:the rates, pathways, and contributions to anaerobic organic matter mineralization. Estuaries and Coasts , 2016, 39 (6) : 1679–1693. DOI:10.1007/s12237-016-0094-0 |
| [20] | Armstrong W. Oxygen diffusion from the roots of some British bog plants. Nature , 1964, 204 (4960) : 801–802. |
| [21] | 邓泓, 叶志鸿, 黄铭洪. 湿地植物根系泌氧的特征. 华东师范大学学报:自然科学版 , 2007 (6) : 69–76. |
| [22] | 刘春英, 陈春丽, 弓晓峰, 周文斌, 杨菊云. 湿地植物根表铁膜研究进展. 生态学报 , 2014, 34 (10) : 2470–2480. |
| [23] | 陆松柳, 张辰, 徐俊伟. 植物根系分泌物分析及对湿地微生物群落的影响研究. 生态环境学报 , 2011, 20 (4) : 676–680. |
| [24] | 吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望. 植物生态学报 , 2014, 38 (3) : 298–310. |
| [25] | Cheng W X, Zhang Q L, Coleman D C, Carroll C R, Hoffman C A. Is available carbon limiting microbial respiration in the rhizosphere?. Soil Biology and Biochemistry , 1996, 28 (10-11) : 1283–1288. DOI:10.1016/S0038-0717(96)00138-1 |
| [26] | Canfield D E, Thamdrup B, Hansen J W. The anaerobic degradation of organic matter in Danish coastal sediments:iron reduction, manganese reduction, and sulfate reduction. Geochimica et Cosmochimica Acta , 1993, 57 (16) : 3867–3883. DOI:10.1016/0016-7037(93)90340-3 |
| [27] | Weiss J V, Emerson D, Megonigal J P. Rhizosphere Iron (Ⅲ) deposition and reduction in a Juncus effusus L.-Dominated wetland. Soil Science Society of America Journal , 2005, 69 (6) : 1861–1870. DOI:10.2136/sssaj2005.0002 |
| [28] | King G M, Garey M A. Ferric iron reduction by bacteria associated with the roots of freshwater and marine macrophytes. Applied and Environmental Microbiology , 1999, 65 (10) : 4393–4398. |
| [29] | Armstrong W, Cousins D, Armstrong J, Turner D, Beckett P. Oxygen distribution in wetland plant roots and permeability barriers to gas-exchange with the rhizosphere:a microelectrode and modelling study with Phragmites australis. Annals of Botany , 2000, 86 (3) : 687–703. DOI:10.1006/anbo.2000.1236 |
| [30] | Cheng H, Wang Y S, Fei J, Jiang Z Y, Ye Z H. Differences in root aeration, iron plaque formation and waterlogging tolerance in six mangroves along a continues tidal gradient. Ecotoxicology , 2015, 24 (7-8) : 1659–1667. DOI:10.1007/s10646-015-1474-0 |
| [31] | Fu Y Q, Yang X J, Ye Z H, Shen H. Identification, separation and component analysis of reddish brown and non-reddish brown iron plaque on rice (Oryza sativa) root surface. Plant and Soil , 2016, 402 (1-2) : 277–290. DOI:10.1007/s11104-016-2802-8 |
| [32] | Hansel C M, Lentini C J, Tang Y Z, Johnston D T, Wankel S D, Jardine P M. Dominance of sulfur-fueled iron oxide reduction in low-sulfate freshwater sediments. The ISME Journal , 2015, 9 (11) : 2400–2412. DOI:10.1038/ismej.2015.50 |
| [33] | Sutton-Grier A E, Megonigal J P. Plant species traits regulate methane production in freshwater wetland soils. Soil Biology and Biochemistry , 2011, 43 (2) : 413–420. DOI:10.1016/j.soilbio.2010.11.009 |
| [34] | Crowder A, St-Cyr L. Iron oxide plaque on wetland roots. Trends in Soil Science , 1991, 1 : 15–329. |
| [35] | Jensen S I, Kühl M, Glud R N, Jørgensen L B, Priemé A. Oxic microzones and radial oxygen loss from roots of Zostera marina. Marine Ecology Progress Series Online , 2005, 293 : 49–58. DOI:10.3354/meps293049 |
| [36] | Nóbrega G N, Ferreira T O, Romero R E, Marques AG B, Otero X L. Iron and sulfur geochemistry in semi-arid mangrove soils (Ceará, Brazil) in relation to seasonal changes and shrimp farming effluents. Environmental Monitoring and Assessment , 2013, 185 (9) : 7393–7407. DOI:10.1007/s10661-013-3108-4 |
| [37] | Roden E E, Wetzel R G. Organic carbon oxidation and suppression of methane production by microbial Fe (Ⅲ) oxide reduction in vegetated and unvegetated freshwater wetland sediments. Limnology and Oceanography , 1996, 41 (8) : 1733–1748. DOI:10.4319/lo.1996.41.8.1733 |
| [38] | Weiss J V, Emerson D, Backer S M, Megonigal J P. Enumeration of Fe (Ⅱ)-oxidizing and Fe (Ⅲ)-reducing bacteria in the root zone of wetland plants:implications for a rhizosphere iron cycle. Biogeochemistry , 2003, 64 (1) : 77–96. DOI:10.1023/A:1024953027726 |
| [39] | Taylor G J, Crowder A, Rodden R. Formation and morphology of an iron plaque on the roots of Typha latifolia L. grown in solution culture. American Journal of Botany , 1984, 71 (5) : 666–675. DOI:10.2307/2443363 |
| [40] | Hansel C M, Fendorf S, Sutton S, Newville M. Characterization of Fe plaque and associated metals on the roots of mine-waste impacted aquatic plants. Environmental Science & Technology , 2001, 35 (19) : 3863–3868. |
| [41] | Li X M, Liu T X, Li F B, Zhang W, Zhou S G, Li Y T. Reduction of structural Fe (Ⅲ) in oxyhydroxides by Shewanella decolorationis S12 and characterization of the surface properties of iron minerals. Journal of Soils and Sediments , 2012, 12 (2) : 217–227. DOI:10.1007/s11368-011-0433-5 |
| [42] | Postma D. The reactivity of iron oxides in sediments:a kinetic approach. Geochimica et Cosmochimica Acta , 1993, 57 (21-22) : 5027–5034. DOI:10.1016/S0016-7037(05)80015-8 |
| [43] | Roden E E, Wetzel R G. Kinetics of microbial Fe (Ⅲ) oxide reduction in freshwater wetland sediments. Limnology and Oceanography , 2002, 47 (1) : 198–211. DOI:10.4319/lo.2002.47.1.0198 |
| [44] | Hines M E, Knollmeyer S L, Tugel J B. Sulfate reduction and other sedimentary biogeochemistry in a northern New England salt marsh. Limnology and Oceanography , 1989, 34 (3) : 578–590. DOI:10.4319/lo.1989.34.3.0578 |
| [45] | Howarth R. Microbial processes in salt-marsh sediments//Ford T E, eds. Aquatic Microbiology:An Ecological Approach. Oxford:Blackwell Scientific, 1993:239-260. |
| [46] | Hyun J H, Mok J S, Cho H Y, Kim S H, Lee K S, Kostka J E. Rapid organic matter mineralization coupled to iron cycling in intertidal mud flats of the Han River estuary, Yellow Sea. Biogeochemistry , 2009, 92 (3) : 231–245. DOI:10.1007/s10533-009-9287-y |
| [47] | Beck M, Dellwig O, Holstein J M, Grunwald M, Liebezeit G, Schnetger B, Brumsack H J. Sulphate, dissolved organic carbon, nutrients and terminal metabolic products in deep pore waters of an intertidal flat. Biogeochemistry , 2008, 89 (2) : 221–238. DOI:10.1007/s10533-008-9215-6 |
| [48] | Colombo C, Palumbo G, He J Z, Pinton R, Cesco S. Review on iron availability in soil:interaction of Fe minerals, plants, and microbes. Journal of Soils and Sediments , 2014, 14 (3) : 538–548. DOI:10.1007/s11368-013-0814-z |
| [49] | Koretsky C M, Van Cappellen P, DiChristina T J, Kostka J E, Lowe K L, Moore C M, Roychoudhury A N, Viollier E. Salt marsh pore water geochemistry does not correlate with microbial community structure. Estuarine, Coastal and Shelf Science , 2005, 62 (1-2) : 233–251. DOI:10.1016/j.ecss.2004.09.001 |
| [50] | Dennis P G, Miller A J, Hirsch P R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities?. FEMS Microbiology Ecology , 2010, 72 (3) : 313–327. DOI:10.1111/fem.2010.72.issue-3 |
| [51] | Kristensen E. Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. Journal of Sea Research , 2008, 59 (1-2) : 30–43. DOI:10.1016/j.seares.2007.05.004 |
| [52] | 田翠翠, 肖邦定. 轮叶黑藻(Hydrilla verticillata)根系泌氧对沉积物中典型铁氧化菌和铁还原菌的影响. 湖泊科学 , 2016, 28 (4) : 835–842. |
| [53] | 拓晓骅, 朱辉, 王保莉, 曲东. 淹水培养过程中水稻土地杆菌科微生物群落结构变化特征. 农业环境科学学报 , 2012, 31 (6) : 1165–1171. |
| [54] | Lowe K L, Dichristina T J, Roychoudhury A N, Van Cappellen P. Microbiological and geochemical characterization of microbial Fe (Ⅲ) reduction in salt marsh sediments. Geomicrobiology Journal , 2000, 17 (2) : 163–178. DOI:10.1080/01490450050023836 |
| [55] | Alzubaidy H, Essack M, Malas T B, Bokhari A, Motwalli O, Kamanu F K, Jamhor S A, Mokhtar N A, Antunes A, Simões M F. Rhizosphere microbiome metagenomics of gray mangroves (Avicennia marina) in the Red Sea. Gene , 2015, 576 (2) : 626–636. |
| [56] | Song K, Lee S H, Kang H. Denitrification rates and community structure of denitrifying bacteria in newly constructed wetland. European Journal of Soil Biology , 2011, 47 (1) : 24–29. DOI:10.1016/j.ejsobi.2010.10.003 |
| [57] | Zhang Y, Cui BS, Xie T, Wang Q, Yan JG. Gradient distribution patterns of rhizosphere bacteria associated with the coastal reclamation. Wetlands , 2016, 36 (S1) : 69–80. DOI:10.1007/s13157-015-0719-2 |
| [58] | Ferreira T, Otero X L, Vidal-Torrado P, Macías F. Effects of bioturbation by root and crab activity on iron and sulfur biogeochemistry in mangrove substrate. Geoderma , 2007, 142 (1-2) : 36–46. DOI:10.1016/j.geoderma.2007.07.010 |
| [59] | Otero X L, Ferreira T O, Huerta-Díaz M A, Partiti C S M, Souza V Jr, Vidal-Torrado P, Macías F. Geochemistry of iron and manganese in soils and sediments of a mangrove system, Island of Pai Matos (Cananeia-SP, Brazil). Geoderma , 2009, 148 (3-4) : 318–335. DOI:10.1016/j.geoderma.2008.10.016 |
| [60] | 唐罗忠, 生原喜久雄, 户田浩人, 黄宝龙. 湿地林土壤的Fe2+, Eh及pH值的变化. 生态学报 , 2012, 25 (1) : 103–107. |
| [61] | Hinsinger P, Plassard C, Tang C X, Jaillard B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints:a review. Plant and Soil , 2003, 248 (1-2) : 43–59. |
| [62] | Neubauer S, Emerson D, Megonigal J. Microbial oxidation and reduction of iron in the root zone and influences on metal mobility//Violante A, Huang P M, Gadd G M, eds. Biophysico-Chemical Processes of Heavy Metals and Metalloids in Soil Environments. Hoboken:John Wiley & Sons, Inc, 2008:339-371. |
| [63] | Laanbroek H. Bacterial cycling of minerals that affect plant growth in waterlogged soils:a review. Aquatic Botany , 1990, 38 (1) : 109–125. DOI:10.1016/0304-3770(90)90101-P |
| [64] | Canfield D E. Reactive iron in marine sediments. Geochimica et Cosmochimica Acta , 1989, 53 (3) : 619–632. DOI:10.1016/0016-7037(89)90005-7 |
| [65] | Gribsholt B, Kostka J E, Kristensen E. Impact of fiddler crabs and plant roots on sediment biogeochemistry in a Georgia saltmarsh. Marine Ecology Progress , 2003, 259 : 237–251. DOI:10.3354/meps259237 |
| [66] | Kristensen E, Alongi D M. Control by fiddler crabs (Uca vocans) and plant roots (Avicennia marina) on carbon, iron, and sulfur biogeochemistry in mangrove sediment. Limnology and Oceanography , 2006, 51 (4) : 1557–1571. DOI:10.4319/lo.2006.51.4.1557 |
| [67] | Tong C, Wang W Q, Huang J F, Gauci V, Zhang L H, Zeng C S. Invasive alien plants increase CH4 emissions from a subtropical tidal estuarine wetland. Biogeochemistry , 2012, 111 (1-3) : 677–693. DOI:10.1007/s10533-012-9712-5 |
| [68] | Rogers M E, Colmer T D, Frost K, Henry D, Cornwall D, Hulm E, Deretic J, Hughes S R, Craig A D. Diversity in the genus Melilotus for tolerance to salinity and waterlogging. Plant and Soil , 2008, 304 (1-2) : 89–101. DOI:10.1007/s11104-007-9523-y |
| [69] | Wu C, Ye Z H, Li H, Wu S C, Deng D, Zhu Y G, Wong M H. Do radial oxygen loss and external aeration affect iron plaque formation and arsenic accumulation and speciation in rice?. Journal of Experimental Botany , 2012, 63 (8) : 2961–2970. DOI:10.1093/jxb/ers017 |
| [70] | Yang J X, Tam N F Y, Ye Z R. Root porosity, radial oxygen loss and iron plaque on roots of wetland plants in relation to zinc tolerance and accumulation. Plant and Soil , 2014, 374 (1-2) : 815–828. DOI:10.1007/s11104-013-1922-7 |
| [71] | Colmer T D. Long-distance transport of gases in plants:a perspective on internal aeration and radial oxygen loss from roots. Plant, Cell & Environment , 2003, 26 (1) : 17–36. |
| [72] | Lu Y H, Watanabe A, Kimura M. Contribution of plant-derived carbon to soil microbial biomass dynamics in a paddy rice microcosm. Biology and Fertility of Soils , 2002, 36 (2) : 136–142. DOI:10.1007/s00374-002-0504-2 |
| [73] | Kostka J E, Gribsholt B, Petrie E, Dalton D, Skelton H, Kristensen E. The rates and pathways of carbon oxidation in bioturbated saltmarsh sediments. Limnology and Oceanography , 2002, 47 (1) : 230–240. DOI:10.4319/lo.2002.47.1.0230 |
| [74] | Sundby B, Vale C, Caetano M, Luther G W III. Redox chemistry in the root zone of a salt marsh sediment in the Tagus Estuary, Portugal. Aquatic Geochemistry , 2003, 9 (3) : 257–271. DOI:10.1023/B:AQUA.0000022957.42522.9a |
| [75] | 鄢璐, 王世和, 雒维国, 黄娟, 钟秋爽. 运行条件下潜流型人工湿地溶氧状态研究. 环境科学 , 2006, 27 (10) : 2009–2013. |
| [76] | Sobolev D, Roden E E. Characterization of a neutrophilic, chemolithoautotrophic Fe (Ⅱ)-oxidizing β-proteobacterium from freshwater wetland sediments. Geomicrobiology Journal , 2004, 21 (1) : 1–10. DOI:10.1080/01490450490253310 |
| [77] | Batty L C, Baker A J M, Wheeler B D, Curtis C D. The effect of pH and plaque on the uptake of Cu and Mn in Phragmites australis (Cav.) Trin ex. Steudel. Annals of Botany , 2000, 86 (3) : 647–653. DOI:10.1006/anbo.2000.1191 |
| [78] | Li H, Ye Z H, Wei Z J, Wong M H. Root porosity and radial oxygen loss related to arsenic tolerance and uptake in wetland plants. Environmental Pollution , 2011, 159 (1) : 30–37. DOI:10.1016/j.envpol.2010.09.031 |
| [79] | Hupfer M, Dollan A. Immobilisation of phosphorus by iron-coated roots of submerged macrophytes. Hydrobiologia , 2003, 506-509 (1-3) : 635–640. DOI:10.1023/B:HYDR.0000008605.09957.07 |
| [80] | Croal L R, Johnson C M, Beard B L, Newman DK. Iron isotope fractionation by Fe (Ⅱ)-oxidizing photoautotrophic bacteria. Geochimica et Cosmochimica Acta , 2004, 68 (6) : 1227–1242. DOI:10.1016/j.gca.2003.09.011 |
| [81] | Mei X Q, Wong M H, Yang Y, Dong H Y, Qiu R L, Ye Z H. The effects of radial oxygen loss on arsenic tolerance and uptake in rice and on its rhizosphere. Environmental Pollution , 2012, 165 : 109–117. DOI:10.1016/j.envpol.2012.02.018 |
| [82] | Bendix M, Tornbjerg T, Brix H. Internal gas transport in Typha latifolia L. and Typha angustifolia L. 1. Humidity-induced pressurization and convective throughflow. Aquatic Botany , 1994, 49 (2-3) : 75–89. DOI:10.1016/0304-3770(94)90030-2 |
| [83] | Colmer T D, Pedersen O. Oxygen dynamics in submerged rice (Oryza sativa). New Phytologist , 2008, 178 (2) : 326–334. DOI:10.1111/j.1469-8137.2007.02364.x |
| [84] | Laskov C, Horn O, Hupfer M. Environmental factors regulating the radial oxygen loss from roots of Myriophyllum spicatum and Potamogeton crispus. Aquatic Botany , 2006, 84 (4) : 333–340. DOI:10.1016/j.aquabot.2005.12.005 |
| [85] | Chen X P, Kong W D, He J Z, Liu W J, Smith S E, Smith F A, Zhu YG. Do water regimes affect iron-plaque formation and microbial communities in the rhizosphere of paddy rice?. Journal of Plant Nutrition & Soil Science , 2008, 171 (2) : 193–199. |
| [86] | 孟冬梅, 朱永官, 周建国. 水稻根系通气组织与根表铁膜关系的研究. 现代农业科学 , 2008, 15 (4) : 55–58. |
| [87] | St-Cyr L, Crowder A A. Factors affecting iron plaque on the roots of Phragmites australis (Cav.) Trin. ex Steudel. Plant and Soil , 1989, 116 (1) : 85–93. DOI:10.1007/BF02327260 |
| [88] | Soana E, Bartoli M. Seasonal variation of radial oxygen loss in Vallisneria spiralis L. An adaptive response to sediment redox? Aquatic Botany , 2013, 104 : 228–232. |
| [89] | Møller C L, Sand-Jensen K. Iron plaques improve the oxygen supply to root meristems of the freshwater plant, Lobelia dortmanna. New Phytologist , 2008, 179 (3) : 848–856. DOI:10.1111/nph.2008.179.issue-3 |
| [90] | 钟顺清. 根表铁膜对2种景观湿地植物根系发育及活力的影响. 水生态学杂志 , 2015 (1) : 74–79. |
| [91] | 张弛, 王树功, 郑耀辉, 陈桂珠. 生物扰动对红树林沉积物中AVS和重金属迁移转化的影响. 生态学报 , 2010, 30 (11) : 3037–3045. |