 2016, Vol. 36
2016, Vol. 36文章信息
- 王丽, 王保栋, 陈求稳, 汤新武, 韩瑞
- WANG Li, WANG Baodong, CHEN Qiuwen, TANG Xinwu, HAN Rui
- 三峡三期蓄水后长江口海域浮游动物群落特征及影响因子
- Characteristics of the zooplankton community and impactfactors in the Yangtze estuary coastal eara after third stage impoundment of the Three Gorges Dam
- 生态学报, 2016, 36(9): 2505-2512
- Acta Ecologica Sinica, 2016, 36(9): 2505-2512
- http://dx.doi.org/10.5846/stxb201412062423
-
文章历史
- 收稿日期: 2014-12-06
- 网络出版日期: 2015-08-26






2. 国家海洋局第一海洋研究所, 青岛 266061;
3. 中国科学院生态环境研究中心, 北京 100085;
4. 三峡大学, 宜昌 443002
2. The First Institute of Oceanography, Qingdao 266061, China;
3. Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
4. China Three Gorges University, Yichang 443002, China
长江口是陆海相互作用的集中地带,有着特殊的生态环境,其生物类群中咸淡水种类并存,分布和变化有别于其它类型水体[1, 2]。20世纪90年代以来,人类活动对长江口生态环境的潜在影响越来越突出,长江口深水航道整治一期工程、二期工程、洋山深水港工程、南水北调工程及三峡工程等大型水利工程的建设和运行可能对长江口的生态环境产生了累积或叠加效应。尤其是三峡工程建成运行后,长江口生态环境的变化日益受到公众的广泛关注[3, 4, 5, 6, 7]。
目前,世界上至少修建有45000座大坝用于发电或水量调控,大坝修建对河流最直接最明显的改变是径流的变化,同时拦截泥沙及营养盐的输移,使得下游河流的营养盐浓度及泥沙含量减少[8]。这些物理化学环境的变化必然对河流下游甚至河口的生态系统结构和功能产生影响。浮游动物作为水生态系统组成的重要中间环节,在生态系统的能量流动和物质循环中起着承上启下的作用,其群落结构变化趋势能够在一定程度上反映水生态环境特征的改变,也通常是指示河口及海洋等生态环境变化特征的理想指标[9, 10]。因此,研究三峡工程蓄水后长江口浮游动物群落结构特征变化及其与环境因子关系,可从浮游动物这一生态系统重要功能群的角度科学评价三峡工程对长江口及邻近海域生态环境影响程度,研究结果对于揭示重大水利工程对长江口生态系统的影响过程和机理具有一定的意义,也可为相关研究提供重要基础资料。
1 材料与方法 1.1 调查海域及取样方法分别于2010年8月(夏季、丰水期)、2010年11月(秋季、枯水期)和2011年5月(春季、平水期)3个不同季节进行水样和浮游动物样品采集,在长江口共设24个监测点位(图 1)。

|
| 图 1 长江口研究区域及采样站位 Fig. 1 The study area of Yangtze estuary and the sampling sites |
样品采集方法和分析测试均根据《海洋调查规范》(GB12763-2007)和《海洋监测规范》(GB17378-2007)进行。对于每个采样点位分别在表层、中层和底层采集水样,并测量水文(温度、盐度)及环境化学(硅酸盐、总溶解态氮、总溶解态磷、溶解氧、悬浮物、pH和叶绿素a)等环境数据。其中温度、盐度和深度用CTD仪测量,水样用Niskin采水器同步采集(标准层)。
浮游动物样品采集用浅水Ⅰ型浮游生物网(网口面积0.2 m2)自海底至海面垂直拖拽获得,用5%福尔马林溶液固定后再进行分类、鉴定、计数和称重,样品鉴定到种。
1.2 数据分析方法浮游动物生物群落结构特征指标主要由优势种和物种多样性表征,优势种根据优势度值 Y 来确定:
式中,ni代表第i种的个体总数,N为所有种类总个体数,fi为出现频率。Y值大于0.02的种类为优势种[4]。
物种多样性指数采用Shannon-Weaver Index,公式为:
式中,H′为多样性指数,Pi为样品中第i种的丰度比例,S 为种数。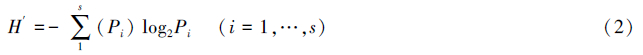
生物与环境因子相关性采用PRIMER 5 (Plymouth Routines In Multivariate Ecological Research)软件进行分析。针对各点位的浮游动物数据和环境数据,首先分别计算基于浮游动物丰度的Bray-Curtis相似性矩阵和基于27个环境因子(3个测量深度上各9个环境因子)的欧氏距离相异矩阵,然后利用Bray-Curtis相似性矩阵和欧氏距离相异矩阵进行BIOENV (Biota-Environment matching)或BVSTEP (Biota-Environment matching using step algorithm)分析,识别与物种种类组成相关性最高的环境因子组合,并计算各环境因子与种类组成的相关性系数。
三峡工程分期蓄水过程包括135(139)、156 m和175 m等3个蓄水期,研究选取三峡蓄水的两个关键节点2002年(135 m)和2006年(156 m)的夏季历史数据与本研究(175 m)夏季数据进行比较分析。考虑到历史数据中浮游动物采集方法和调查侧重点的差异性及局限性,研究将浮游动物种类聚类到门的水平上进行比较,最大程度上增强历史数据的可比性。
2 结果与分析 2.1 浮游动物群落结构组成3个航次共鉴定浮游动物成体164种,浮游幼体14种,共计178种。其中夏季种类最多(152种)、秋季(131种)次之、春季(97种)最少。从图 2可以看出,桡足类、水母类和浮游幼体在3个不同季节均占据优势地位,尤其是桡足类在所有种类数中占绝对优势。春、夏、秋3个季度中桡足类占所有物种种类数的百分比分别为34%、48%和49.6%,水母类为13.4%、6.6%和6.9%,而浮游幼体则为20.6%、17.2%和18.3%。

|
| 图 2 浮游动物种类组成 Fig. 2 Species composition of zooplankton |
此外浮游动物优势种季节变换明显(表 1)。夜光虫(Noctiluca scientillans)与中华哲水蚤(Calanus sinicus)为春季绝对优势种,优势度分别为0.382和0.324。夏季绝对优势种为太平洋纺锤水蚤(Acartia pacifica)(Y=0.226),秋季为针刺拟哲水蚤(Paracalanus aculeatus)(Y=0.187)。3个季节共有的优势种为长尾类幼体(Macrura larva),优势度在不同季节没有太大的差异。春夏季的共同优势种为夜光虫(Noctiluca scientillans),夏秋季的共同优势种为太平洋纺锤水蚤(Acartia pacific)和针刺拟哲水蚤(Paracalanus aculeatus),但优势度存在较大的变化(表 1)。
| 季节Season | 序号Number | 优势种Dominant Species | 类群Ecological Group | 优势度Dominance |
| 春季Spring | 1 | 夜光虫Noctiluca scientillans | 原生动物 | 0.382 |
| 2 | 中华哲水蚤Calanus sinicus | 桡足类 | 0.324 | |
| 3 | 长尾类幼体Macrura larva | 浮游幼体 | 0.020 | |
| 夏季Summer | 1 | 太平洋纺锤水蚤Acartia pacifica | 桡足类 | 0.226 |
| 2 | 长尾类幼体Macrura larva | 浮游幼体 | 0.091 | |
| 3 | 肥胖三角蚤Evadne tergestina | 桡足类 | 0.086 | |
| 4 | 夜光虫Noctiluca scientillans | 原生动物 | 0.086 | |
| 5 | 针刺拟哲水蚤Paracalanus aculeatus | 桡足类 | 0.060 | |
| 秋季Autumn | 1 | 针刺拟哲水蚤Paracalanus aculeatus | 桡足类 | 0.187 |
| 2 | 拟哲水蚤幼体Paracalanus larva | 桡足类 | 0.072 | |
| 3 | 长尾类幼体Macrura larva | 浮游幼体 | 0.043 | |
| 4 | 太平洋纺锤水蚤Acartia pacifica | 桡足类 | 0.038 | |
| 5 | 背针胸刺水蚤Centropages dorsispinatus | 桡足类 | 0.033 |
3个季节浮游动物丰度和生物量(以湿重计)空间分布如图 3所示,平均值及峰值位置见表 2。长江口调查海域浮游动物平均生物量季节相差较大(表 2),春季平均生物量为571.5 mg/m3,峰值区出现在长江口东部海域(122.75°,E 31.00°N附近);夏季平均生物量为970.6 mg/m3,峰值区同样出现在长江口东部海域,但比春季向南偏移;秋季生物量为613.8 mg/m3,高值区出现在长江口口门(122°E,31.00°N附近)。从平均丰度来看,浮游动物丰度春夏季较高,分别为691.3 个/m3和783.5个/m3,秋季最低,丰度仅为399.5 个/m3。同时,夏季和秋季的浮游动物丰度分布基本与生物量分布一致,但春季丰度与生物量分布存在空间差异(图 3)。

|
| 图 3 长江口浮游动物丰度及生物量空间分布 Fig. 3 Spatial distribution of abundance and biomass of zooplankton in the Yangtze estuary |
| 季节Season | 丰度Abundance/(个/m3) | 生物量Biomass/(mg/m3) | ||||
| 变化范围Range | 平均值Average | 峰值坐标Peak Location | 变化范围Range | 平均值Average | 峰值坐标Peak Location | |
| 春季Spring | 71—3185.4 | 691.3 | 122.25E, 31.00N | 10—2563.8 | 571.5 | 122.75E,31.50N |
| 夏季Summer | 67—2187.5 | 783.5 | 123.50E, 31.00N | 393.7—2021.4 | 970.6 | 122.75E,31.50N |
| 秋季Autumn | 33.6—2717.4 | 399.5 | 122.05E, 31.55N | 164.5—2437.5 | 613.8 | 122.75E,31.50N |
在不同季节中,春季浮游动物对丰度贡献率最大的物种是个体较大的中华哲水蚤和夜光虫,二者之和占到40%以上;夏秋季的主要贡献物种分别是太平洋纺锤水蚤和针刺拟哲水蚤,其中太平洋纺锤水蚤在这两个季度的贡献率分别为12.8%和15.9%,而针刺拟哲水蚤则为9.5%和8.0%。
2.3 影响浮游动物分布的关键环境因子识别浮游动物种类组成和群落结构特征与水温、盐度、水系、径流等非生物因子密切相关。生物与环境因子相关性的BIOENV和BVSTEP分析结果表明,春季能够解释浮游动物群落结构的最佳环境因子组合为中层温度、底层盐度及底层硅酸盐,该环境变量组合与物种分布的相关系数为0.654;夏季为中层盐度、中层硅酸盐及中层总溶解态氮,相关系数为0.746;秋季为底层温度、表层悬浮物及中层硅酸盐,相关系数为0.833。
2.4 3个蓄水时期浮游动物群落结构比较本研究选择了调查区域和调查季节高度重合的历史数据(已发表文献)进行比较分析,3个蓄水期夏季浮游动物丰度和群落结构组成分别见表 5和图 4。总体上三峡蓄水后浮游动物生物量及丰度均呈上升趋势,而浮游动物的Shannon-Wiener多样性指数变化范围不大,在2.12—2.34之间。
| 季节Season | 序号Number | 种名Spcise | 平均丰度/(个/m3)Averageabundance | 贡献率/%Contributionrate |
| 春季Spring | 1 | 中华哲水蚤Calanus sinicus | 233.7 | 21.7 |
| 2 | 夜光虫Noctiluca scientillans | 275.7 | 15.8 | |
| 3 | 箭虫幼体Arrow worm larva | 13.5 | 9.6 | |
| 4 | 长尾类幼体Macrura larva | 14.9 | 9.7 | |
| 5 | 中华箭虫Sagitta sinica | 12.6 | 7.2 | |
| 夏季Summer | 1 | 太平洋纺锤水蚤Acartia pacifica | 176.7 | 12.8 |
| 2 | 针刺拟哲水蚤Paracalanus aculeatus | 52.4 | 9.5 | |
| 3 | 长尾类幼体Macrura larva | 75.3 | 7.5 | |
| 4 | 箭虫幼体Arrow worm larva | 17.7 | 6.2 | |
| 5 | 中华哲水蚤Calanus sinicus | 53.7 | 5.7 | |
| 秋季Autumn | 1 | 针刺拟哲水蚤Paracalanus aculeatus | 77.9 | 15.9 |
| 2 | 长尾类幼体Macrura larva | 19.8 | 8.0 | |
| 3 | 精致真刺水蚤Euchaeta concinna | 10.7 | 5.9 | |
| 4 | 小毛猛水蚤Microsetella norvegica | 10.4 | 5.8 | |
| 5 | 箭虫幼体Arrow worm larva | 16.8 | 5.7 |
| 季节Season | 影响因子Governingfactor | 相关系数Correlationcoefficient |
| s:表层;m:中层;b:底层 | ||
| 春季Spring | 温度(m)、盐度(b)、 硅酸盐(b) | 0.654 |
| 温度(m)、盐度(m)、硅酸盐(b) | 0.646 | |
| 温度(b)、盐度(b)、硅酸盐(b) | 0.645 | |
| 夏季Summer | 盐度(m)、硅酸盐(m)、总溶解态氮(m) | 0.746 |
| 盐度(m)、硅酸盐(b)、总溶解态氮(m) | 0.744 | |
| 硅酸盐(m) | 0.744 | |
| 秋季Autumn | 温度(b)、悬浮物(s)、硅酸盐(m) | 0.833 |
| 温度(b)、悬浮物(s)、总溶解态氮(s) | 0.828 | |
| 温度(b)、悬浮物(s)、总溶解态氮(m) | 0.828 | |

|
| 图 4 三峡蓄水前后长江口浮游动物种类比例变化 Fig. 4 Zooplankton species percentage in Yangtze estuary before and after Three Gorges Dam |
三峡工程不同蓄水时期的浮游动物群落组成也不相同(图 4),蓄水前后桡足类所属节肢动物门所占比例呈轻微下降趋势,由2002年的65.4%下降到2010年的57.9%,而水母类所属腔肠动物门所占比例呈上升趋势,2002年夏季未检测到其存在,而到2010年夏季调查中其在浮游动物物种组成中所占比率达到6.6%。
3 讨论 3.1 浮游动物群落结构的季节变化特征相关文献表明,近年来长江口及其毗邻海域饵料生物组成者桡足类的组成比例呈下降趋势,而非饵料生物水母比例呈上升趋势[11]。尽管本研究调查时段内桡足类在3个不同季节的种类数均占有绝对优势地位,但水母类种类数所占比例相比其他种类要高,特别是水母类种类数在春季所占比例最高,而桡足类种类数在春季所占比例最小,这可能与水母类几乎无选择的摄食一切可获得的浮游动物(尤其桡足类),从而控制其种群数量有关[12]。同时,徐兆礼等[13]相关研究表明水母类主要分布在水温和盐度都较低的近海海域(水温14—20℃,盐度28—32),并且长江冲淡水扩展方向是决定水母类高密度聚集区位置的主要因素。三峡蓄水后,年内洪季(夏、秋季)的径流量和所占比例减小,枯季(春季)径流量和所占比例相应增大[7],春季长江口水域环境变得更有利于水母类生长,这可能是导致水母类所占比例上升的原因之一。另外,调查海域浮游植物群落结构也发生了明显变化,甲藻种类组成比例和密度比例均呈现明显上升趋势,而硅藻密度正处于下降过程[14]。郑珊研究表明水母的代谢及分解过程使水体表现出明显的低氧(缺氧)和酸化现象,同时分解过程还释放出大量的营养盐并改变原有的营养盐结构,可以刺激甲藻和绿藻的生长,甚至引发藻华[15]。
此外,从年内来看浮游动物优势种季节变换明显。中华哲水蚤作为优势种仅在春季出现。温度被认为是限制中华哲水蚤分布的最主要原因[16],春季长江口温度变化范围16—17℃正好在中华哲水蚤最适水温范围内(10—20℃)[17],其丰度春季最大,夏季次之,秋季最小。春夏季的共同优势种夜光虫是亚热带和热带海区引发赤潮的主要物种之一,也是引起我国赤潮频发的重要种类,其分布和营养盐的分布密切相关,夜光虫出现数量的多寡可作为指示水环境污染程度的一个依据[18, 19]。夜光虫是偏冷水性的种类,最适温度一般是16—24℃,主要分布在近岸低盐水体[20]。3个航次春季夜光虫平均丰度最高,达275.7 个/m3,可能为近岸富营养加剧所致。夏秋季的共同优势种为太平洋纺锤水蚤和针刺拟哲水蚤,均属于近岸低盐种,广泛分布在长江口、厦门港、台湾海峡、黄海、渤海等许多海域,季节变化主要受温度的限制[18]。
3.2 浮游动物丰度及生物量的空间变化特征从浮游动物空间分布可以看出,春季浮游动物丰度高值区出现在长江口口门附近,夜光虫是对该季节浮游动物丰度贡献率最大的物种。一方面可能由于近岸水体富营养化严重,盐度较低,利于夜光虫生长;另一方面可能是近岸水母类聚集,使桡足类数量下降,夜光虫等其他种类浮游动物生长压力减小。而春季生物量高值区出现在长江口口门外,主要原因可能在于此处是个体较大的中华哲水蚤密集区,从而导致春季丰度与生物量空间分布不同步现象的出现。夏季浮游动物丰度与生物量分布趋势基本一致,主要呈现出由近岸向外海增高趋势,这主要由于太平洋纺锤水蚤的影响,该物种最适温度在25℃左右,最适盐度在26左右[21]。调查区域夏季温度范围21—30℃,而盐度变化范围较大,在7—34之间,可以看出浮游动物夏季空间分布主要受到盐度影响。秋季浮游动物主要分布在近岸口门附近,主要由于秋季的绝对优势种针刺拟哲水蚤出现在近海海域,其分布受温度的限制。
3.3 影响浮游动物分布的关键环境因子长江口水文环境复杂,同时受到长江径流、苏北沿岸流、闽江沿岸流及台湾暖流等水团的影响,物理化学指标由近岸向外海及由表层向底层都存在浓度梯度。从BIOENV结果看,底层温度在春秋季与浮游动物分布相关性较大,而在夏季相关性不显著。长江口水温变化范围13—30℃,从浮游动物生态类型可知长江口浮游动物以暖水种为主,长江口夏季水温最高,研究区域水温适合浮游动物优势种群桡足类生长。因此,温度不是长江口夏季主要影响因子。浮游动物的分布除与温度有关外,与盐度也有较密切的关系[22, 23],本研究结果表明盐度与长江口春季和夏季浮游动物分布相关性显著,尤其是在夏季。另外,3个季节浮游动物分布与硅酸盐的相关性较高,而长江口浮游动物主要优势种桡足类皆属植食性,主要摄食硅藻[24],作者认为这主要是硅酸盐通过影响硅藻对浮游动物产生间接作用。
4 结论长江口浮游动物群落结构正在发生潜在变化。由于浮游动物生长、发育受到多种因素的影响,且生物对环境变化的响应存在一定的适应性和滞后性,从目前来看,三峡工程2003—2011年的运行对长江口浮游动物的影响还不显著,其长期效应尚需要持续观测研究。
就三期蓄水后的现状来看,影响长江口浮游动物分布的关键环境因子为底层盐度、底层温度及底层硅酸盐(浮游植物)。
致谢:研究数据主要来源于国家“973计划”重大水利工程影响下长江口环境与生态安全项目,由国家海洋局第二海洋研究所监测提供。王保栋、孙霞和辛明等在样品采集、室内样品处理方面做了大量工作,特此致谢。| [1] | 陆健健. 河口生态学. 北京: 海洋出版社, 2003. |
| [2] | 郑金秀, 胡菊香, 彭建华, 汪红军. 长江口南北支浮游动物群落生态学研究. 生态环境学报, 2011, 20(6/7): 1102-1106. |
| [3] | 孙薇. 长江口轮虫群落生态特征分析[D]. 上海: 上海海洋大学, 2012. |
| [4] | 章飞燕, 唐静亮, 李道季, 方涛, 王彪. 夏、秋季长江口及毗邻海域浮游动物的分布与变化. 水生生物学报, 2009, 33(6): 1219-1225. |
| [5] | 余立华, 李道季, 方涛, 李云, 高磊. 三峡水库蓄水前后长江口水域夏季硅酸盐、溶解无机氮分布及硅氮比值的变化. 生态学报, 2006, 26(9): 2817-2826. |
| [6] | 崔彦萍, 王保栋, 陈求稳, 黄国鲜, 牛智星. 三峡水库三期蓄水前后长江口硅酸盐分布及其比值变化. 环境科学学报, 2013, 33(7): 1974-1979. |
| [7] | 曹勇, 陈吉余, 张二凤, 陈沈良, 曹卫承. 三峡水库初期蓄水对长江口淡水资源的影响. 水科学进展, 2006, 17(4): 554-558. |
| [8] | Domingues R B, Barbosa A B, Galvão H M. River damming leads to decreased phytoplankton biomass and disappearance of cyanobacteria blooms. Estuarine, Coastal and Shelf Science, 2014, 136: 129-138. |
| [9] | 纪焕红, 叶属峰. 长江口浮游动物生态分布特征及其与环境的关系. 海洋科学, 2006, 30(6): 23-30. |
| [10] | 刘镇盛. 长江口及其邻近海域浮游动物群落结构和多样性研究[D]. 青岛: 中国海洋大学, 2012. |
| [11] | 杨宇峰, 黄祥飞. 浮游动物生态学研究进展.湖泊科学, 2000, 12(1): 81-89. |
| [12] | 马喜平, 凡守军. 水母类在海洋食物网中的作用. 海洋科学, 1998, 22(2): 38-42. |
| [13] | 徐兆礼. 东海水母类丰度的动力学特征. 动物学报, 2006, 52(5): 854-861. |
| [14] | 李云, 李道季, 唐静亮, 王益鸣, 刘志刚, 丁平兴, 何松琴. 长江口及毗邻海域浮游植物的分布与变化. 环境科学, 2007, 28(4): 719-729. |
| [15] | 郑珊. 水母暴发与浮游植物群落的相互影响研究[D]. 北京: 中国科学院研究生院(海洋研究所), 2013. |
| [16] | 李少普. 福建沿海太平洋哲镖溞(Calanus pacificus Brodsky)的比较形态研究. 厦门大学学报, 1963, 10(1): 57-81. |
| [17] | Huang J Q, Zhen Z. The effects of temperature and salinity on the survival of some copepods from Xiamen harbour. Oceanologia Et Limnologia Sinica, 1986, 17(2): 161-167. |
| [18] | 徐家铸, 苏翠荣. 海州湾浮游动物的种类组成和分布(Ⅰ)-夜光虫和砂壳纤毛虫. 南京师大学报: 自然科学版, 1995, 18(4): 125-129. |
| [19] | 张天文, 朱丽岩, 徐培培, 周浩, 戚本金. 胶州湾夜光虫丰度的季节变化和分布特征. 中国海洋大学学报, 2009, 39(增刊): 89-93. |
| [20] | 徐兆礼. 长江口邻近水域浮游动物群落特征及变动趋势. 生态学杂志, 2005, 24(7): 780-784. |
| [21] | 高亚辉, 林波一. 几种因素对太平洋纺锤水蚤摄食率的影响. 厦门大学学报, 1999, 38(5): 751-757. |
| [22] | 刘光兴, 陈洪举, 朱延忠, 齐衍萍. 三峡工程一期蓄水后长江口及其邻近水域浮游动物的群落结构. 中国海洋大学学报, 2007, 37(5): 789-794. |
| [23] | 陈洪举, 刘光兴. 2006 年夏季长江口及其邻近水域浮游动物的群落结构. 北京师范大学学报: 自然科学版, 2009, 45(4): 393-398. |
| [24] | 杨纪明. 渤海桡足类(Copepoda)的食性和营养级研究. 现代渔业信息, 2001, 16(6): 6-10. |