 2016, Vol. 36
2016, Vol. 36文章信息
- 周绍春, 孟根同, 尹远新, 杨娇, 于洪伟, 任梦飞, 刘浩
- ZHOU Shaochun, MENG Gengtong, YIN Yuanxin, YANG Jiao, YU Hongwei, REN Mengfe, LIU Hao
- 完达山东部林区生境特征对东北林蛙产卵的影响
- Effect of habitat characteristics on the reproductive effort of dybowsky's frogs (Rana Dybowskii) in eastern Wanda Mountains, Heilongjiang Province
- 生态学报, 2016, 36(9): 2521-2527
- Acta Ecologica Sinica, 2016, 36(9): 2521-2527
- http://dx.doi.org/10.5846/stxb201411232318
-
文章历史
- 收稿日期: 2014-11-23
- 网络出版日期: 2015-08-26






2. 东北大学, 沈阳 110819;
3. 东北师范大学环境学院, 长春 130042
2. College of Information Science and Engineering, Northeastern University, Shenyang 110819, China;
3. College of Urban and Environmental Sciences, Northeast Normal University, Changchun 130042, China
对于在池塘生境完成繁育的两栖类动物,他们的生存需要水生和陆地生境才能完成其生活周期。但在过去30年有关两栖类动物生境选择的研究中,其研究重点主要集中在繁育池内的生境特征与繁育关系方面,而没考虑繁育池周围的生境与两栖类动物生存之间的关系[1]。然而,在两栖类动物的生活周期中,其陆地生活时间较水环境生活时间更长。因此,近年来对两栖类动物研究的焦点已经从单一的繁育池繁育阶段延伸到陆地生存阶段[2, 3]。
在21世纪初期的研究表明,从蛙产卵到孵化期间,繁育池内的隐蔽度、植被组成和水文是影响两栖类动物繁育的关键因素[4, 5, 6];随着研究的不断深入,Ficetola等人的研究表明,陆地环境不仅是蛙类等两栖动物繁育后陆地栖息生活阶段的必须生境,而且繁育池周围陆地生境特征也能影响到繁育池及周边水湿地内的生境因子[7]。研究表明,许多两栖类物种的繁育都与繁育池分布范围在200—1000m内的植被特征紧密联系[8, 9]。在蛙类动物陆地栖息生活阶段,栖息生境上坡位丛林结构的变化可能严重影响蛙类动物的迁徙[10]。两栖类动物从水生环境迁徙到陆生环境表明了该类物种在不同阶段对生境的不同需求[11]。尽管逐渐增多的研究已经证明了在不同生活阶段生境因子对两栖类动物的生活产生影响,但至今未见有关东北林蛙繁殖期,在不同空间尺度生境因子对该物种产卵团数关系的研究。本研究目标是在林蛙繁育期5—7月,通过对微生境选择(繁育池选择)和景观尺度生境选择(由于在野外调查过程中发现,在距繁育池约5km的生境中也发现林蛙个体,本研究定义距繁育池5km范围内的生境作为景观尺度的生境选择)开展研究,探究在不同的空间尺度各生境因子对东北林蛙繁育期产卵团数的影响,这将有助于了解东北林蛙的生态习性,加强该物种栖息生境的保护。
1 研究地区概况研究地区位于黑龙江省完达山东部林区迎春林业局五泡林场。地处中国东北隅,隶属于黑龙江省虎林市,佳木斯市饶河县和宝清县境内。地理坐标为:东经127°04′04″—127°16′45″,北纬46°27′31″—46°36′60″,面积155.6 km2。
该区属寒温带大陆性季风气候,受季风影响强烈,冬季漫长,无霜期为120d左右。冰冻期较长,一般在10月份结冻至第2年4月下旬解冻,冰期约7个月。年平均温度在2.5—3.0℃,年降雨量为500—600mm,60%—70%降雨量集中在6—8月份。年平均湿度为67%—71%,年平均日照为2400 h,无霜期为110—130 d。气温较低,年平均气温1.4—2.2℃,夏季最高气温可达34.6℃,冬季最低气温-34.8℃。这里森林覆盖率达85%,主要由针叶林、针阔混交林、阔叶混交林、灌丛杂木林和沼泽湿地组成。针叶林主要有红松(Pinus koraiensis)、云杉(Picea polita)、冷杉(Akjes fabri)和落叶松林(Lalix olgensis);针阔混交林中高大乔木以落叶松、红松、紫椴(Tilia Amurensis)、核桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、榆树(Ulmus propinqua)、山桃(Padus maackii davidiana)等组成的混合林型;阔叶林由柞树(Quercus mongolica)、白桦(Betula platyphylla)等组成;阔叶混交林以杨树(Populus spp.)、白桦、黄檗(Phellodendron Amurense)等为主;灌丛杂木林多分布在坡缘谷地,在整个地区呈分散分布。树种构成主要以亚乔木暴马子(Syringa Amurensis)、刺五加(Acanthopanx senticosus)、龙牙楤木(Aralia elata)、空心柳(Spiraea salicifola)、榛子(Corylusheterophylla)、杜香(Ledumpalustre)等树种以及天然更新的椴树丛林和桦树丛林组成。丰富的植物多样性使该地区成为东北林蛙的主要分布区。
2 研究方法 2.1 研究地点选择为了识别潜在的林蛙繁育地,本研究通过访问当地的林区居民,再结合1:50000地形图、Sopt5(309/256)卫星影像遥感图片最终选择了105个林蛙繁育池。根据已有的研究成果[12, 13],选择的潜在繁育池必须满足3个条件:1)由于永久性开放的水池(有大量的水源流进和流出)可能存在鱼类,因此选择潜在繁育池都是非永久性开放;2)繁育池周围至少50%生境与森林接壤,森林可为繁殖期的林蛙提供陆地栖息生境;3)在距繁育池5km范围内至少50%生境被森林覆盖。
2.2 繁育池资料收集调查时间为2012—2014年的5月初至7月末林蛙产卵至幼蛙上山期间。在繁育池内部,收集了8个变量。
1)水文:水文时间单位为周(从5月1日至7月30日共12周)。从2012年至2014年研究期间,研究人员每周观察1次繁育池是否有水并测量水表面面积;2)繁育池水深:用米尺测量(m);3 )海拔:由GPS定位获取(m);4)繁育池水表面面积:研究人员在接近水面区域围绕水池行走一周,并利用GPS记录航迹,然后利用GIS软件计算面积,并根据不同水文周期的单个繁育池水体表面面积变化计算其平均值;5)繁育池水表面杂物(如树叶、杂草)盖度:水池表面的覆盖度百分比(%);6)林木郁闭度:繁育池上空林木遮挡百分比(%);7)灌丛郁闭度:繁育池内灌木垂直于水面,遮挡繁育池水面的百分比(%);8)水温:温度计测量(℃)。
2.3 景观变量收集购买的分辨率为10m的SPOT 5卫星影像图片(2011年09月310/255),利用ENVY软件进行景观分类,最终获得景观变量。
1)居民区面积百分比(%):通过购买的SPOT 5卫星影像进行景观分类,然后计算居民区面积; 2)距公路距离:利用野外调查收集的繁育池坐标点和景观分类图层,在GIS 9.3软件中分析获得;3)林型及面积(km2):通过购买的SPOT 5卫星影像进行景观分类,然后计算获得。虽然已有的研究表明,在繁育池周围1km范围内的森林分布是影响部分蛙类动物生境选择的关键因子[12, 14],但由于野外调查发现距繁育池5km远的区域仍然发现东北林蛙个体,因此本研究计算的森林面积百分比为距繁育池5km范围内的森林面积比例。
2.4 东北林蛙雌性繁殖个体与卵团数关系野外调查了105个水池,其中98个繁育池发现蛙卵团。已有的研究和本次调查结果表明,每1只林蛙在1个繁育池只产1个卵团[15]。卵团数与繁育雌性林蛙个体成1:1的关系,该比例关系提供了繁育池内蛙卵团数与雌性繁育蛙个体数的关系系数,同时也为建立环境因子对繁育池内蛙卵团数,即林蛙繁育期环境因子对林蛙生境选择的影响提供了科学依据。
2.5 统计分析广义可加模型(Generalized Additive Models,GAM)由Hastie 和Tibshirani[16]提出,该模型的优点在于适用于响应变量与解释变量之间的关系是非线性和非单调的数据分析[17]。本研究建立蛙卵团数与环境因子之间关系的GAM预测模型分析环境因子对单个繁育池内蛙卵团数的影响,所有的统计分析均在R软件的MGCV包完成。繁育池内蛙卵数与繁育池内生境因子的表达式为:
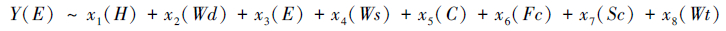
景观水平蛙卵团数与生境因子关系的表达式为:

繁育池内林蛙的卵团平均数为29.00团,范围在0—121之间。在繁育池内生境因子中水文周期平均值为11.91周,在2012年夏季,研究地区雨水相对2013和2014少,发现3个繁育池出现干涸情况。繁育池面积在4—300m2之间,平均值为55.88m2,最小面积为4m2,繁育池内环境因子统计详见表 1。
| 变量Variable | 均值Mean | 范围Range |
| 卵团数Egg masse counts/团 | 29 | 0—121 |
| 水文Hydroperiod/周 | 11.91 | 7.00—12.00 |
| 水深Water depth/m | 32.37 | 15—80 |
| 海拔Elevation/m | 178.00 | 17.1—316.90 |
| 繁育池面积Pond size/m2 | 55.88 | 4—300 |
| 池塘水表面杂物盖度 Sundries coverage ration over pond water/% | 18.37 | 0—45 |
| 林木郁闭度Wood canopy density/% | 26.04 | 0—99 |
| 水温 Water Temperature/℃ | 10.77 | 7.5—13 |
| 灌丛郁闭度Shrub canopy density/% | 15.37 | 0—36 |
| 居民区面积百分比Size percent of residential area/% | 0.05 | 0.05 |
| 灌丛湿地面积Size of shrub and wetland/km2 | 0.02 | 0.01—0.15 |
| 森林面积百分比(Area of forest size)/% | 0.87 | 0.56—1.00 |
| 距公路距离(Distance to roads)/km | 1.46 | 0.00—5.00 |
GAM模型统计结果表明,从偏差变化、AIC和GCV值看出,该模型对孵化池内林蛙卵团数的总偏差解释率为76.53%,截距为35.32,繁育池面积对模型贡献为0.17,森林郁闭度为-0.30,池塘表面杂物覆盖度为-0.43,其他统计参数见表 2。
| 因子Factor | 累计解释偏差Sum of deviance | 赤池信息量准则Akaike information criterion | 广义交叉验证Generalized cross validation |
| 繁育池面积 Breeding pond size(b)/m2 | 35.06 | 473.63 | 1.00 |
| 林木郁闭度Wood canopy (b)/% | 52.89 | 463.54 | 0.93 |
| 池塘水表面杂物盖度Sundries coverageration over surface of pond water (b)/% | 76.53 | 474.39 | 1.01 |
在GAM预测模型中,繁育池面积、森林郁闭度和池塘水表面杂物盖度对林蛙卵团数产生影响,其关系如图 1。

|
| 图 1 蛙卵团数在繁育池微生境尺度的广义可加模型变量曲线变化图 Fig. 1 Generalized additive model response curves (solid lines) with 95% confidence intervals (dashed parts) presented for each of the covariates in the linear predictor scale from the best-fit models for egg mass within ponds microhabitat scale level. |
GAM模型统计结果表明,森林面积百分比和距公路距离对该模型的总偏差解释率为69.15%,截距为14.47,生境因子森林面积比例对模型贡献为17.99,距公路的距离对模型贡献为1.40,其他参数见表 3。
| 因子Factor | 累计解释偏差Sum of deviance | 赤池信息量准则Akaike information criterion | 广义交叉验证Generalized cross validation |
| 森林面积百分比Percent of forest size(b)/% | 38.50 | 433.62 | 0.98 |
| 距公路距离Distance to roads(b)/km | 69.15 | 393.45 | 0.95 |
在景观尺度,森林面积和距公路距离对蛙卵团数产生影响如图 2。

|
| 图 2 蛙卵团数在景观尺度的广义可加模型变量曲线变化图 Fig. 2 Generalized additive model response curves (solid lines) with 95% confidence intervals (dashed parts) presented for each ofthe covariates in the linear predictor scale from the best-fit models for egg mass at landscape scale level. |
本研究重点探究了在中国林蛙繁殖期,繁育池内的蛙卵团数与繁育池内和景观尺度生境因子之间相互关系,该研究成果对于估计林蛙种群大小,保护东北林蛙的栖息生境具有重要的意义。在繁育池内微生境水平,繁育池内蛙卵团数受繁育池面积(b=0.17)、林木郁闭度(b=-0.30)、水表面杂物盖度(b=-0.43)的影响。在景观尺度水平,繁育池内蛙卵团数受森林覆盖面积比例(b=17.99)和距公路距离的影响(b=1.40)。
野外调查过程中在93.33%的繁育池中发现了林蛙卵团。繁育池平均卵团数为29.00(0—121);7个繁育池研究期间没有发现林蛙卵团,67个池卵团数小于29。有研究表明,蛙卵数与繁育池面积成正相关[3],本研究结果表明,繁育池面积从4 m2增加到150m2的过程中,蛙卵团数也呈现增加趋势;但当繁育池面积增加到150m2后,蛙卵团数随繁育池面积增加呈现下降趋势。这可能与东北林蛙选择繁育池的环境因子,包括周围的植被种类、植被构成,以及水文环境等因子有关[5, 19]。此外,Skidds and Golet的研究表明,两栖类动物繁育池面积与繁育池深度成正相关[3],这与本文的研究结果相同(r=0.85)。繁育池水越深,越利于林蛙越冬。同时,繁育池面积大,水越深,能保持水温稳定,降雨爆发山洪,面积大的繁育池不宜泛滥,能够保证蛙卵安全。在本研究地区,调查中面积大的繁育池多位于公路边,由于人类活动干扰,导致这部分繁育池蛙卵团数较少。因此,繁育池内蛙卵团数随繁育池面积的增加呈现波浪式变化,在繁育池面积达到150m2后,繁育池蛙卵团开始下降(图 1)。
此外,林木郁闭度也是影响林蛙繁育生境选择的一个重要因素,本研究表明,随着林木郁闭度的增加,繁育池内林蛙卵团数将减少(图 1),这可能与繁育池温度有关。本研究在林蛙繁育初期测量了55个繁育池水温,水温范围为7.5—13℃,郁闭度大于40℃和郁闭度小于或等于40℃的水温平均相差1.85℃,水温较高的繁育池林蛙卵孵化期比温度低的繁育池蛙卵孵化期短3—7d,平均孵化天数少4.5d,林木郁闭度对蛙产卵产生负面影响(
有研究表明,繁育池内的杂物对蛙卵数有重要影响。Skidds 和 Golet的研究表明[3],繁育池内蛙卵数随繁育池水表面杂物盖度增加而增加。相关的研究认为,繁育池水表面杂物有利于蛙产卵,也可以为蝌蚪、幼蛙提供隐蔽物[20]。但Egan 和 Paton的研究认为,繁育池蛙卵团数不受繁育池水表面杂物的影响[5]。本研究结果表明,繁育池水表面杂物覆盖度总体与繁育池内蛙卵数成负相关,但这种变化趋势可分为正相关和负相关两个部分。当水表面杂物覆盖度从0增加到约14%的范围内,繁育池内的蛙卵数与繁育池水表面杂物覆盖度呈现正相关;但当繁育池水表面杂物盖度大于35%以后,繁育池内蛙卵数则呈现下降趋势;此外,蛙卵数还受诸多环境要素的影响,繁育池内蛙卵数与繁育池内水表面杂物盖度呈现非直线性变化还需要进一步研究(图 1)。
在景观尺度水平,森林覆盖率是影响两栖类动物生存的重要影响因素之一[21]。Porej的研究表明,繁育池附近200m范围属于林蛙陆地生活期间的核心区域[8];Guerry and Hunter的研究发现,在林蛙繁殖季节5km范围内森林覆盖率与该区域内池塘被林蛙侵占比率成正相关[22]。在研究地区,森林覆盖率较平均值为85.00%,繁育池1km范围内森林覆盖达到87.00%。繁育池内蛙卵数随1km范围内森林覆盖面积比例增加而增加(图 2)。本文的研究结果,在距繁育池5km范围内,蛙卵团数仍然与森林面积成正相关,再一次证明了森林覆盖对两栖类动物生存的重要性。
此外,有研究表明,蛙类等两栖动物由于对水生生境和陆地生境的双重需求,生境退化将对该物种的生存产生严重影响,其中公路修建是影响两栖类动物生境选择的主要因素之一[23]。Fahrig 1995年的研究表明,两栖类动物分布的丰富度与公路密度成反比[24]。本研究表明,在距公路约3km的范围内,林蛙卵团数随公路距离的增加而增加,当距公路距离大于3km后,蛙卵团数将随公路距离的增加而减少,本研究和其它研究认为这可能与环境因子地形相关。在研究地区北部,在距公路大于1km,从灌丛过度到沼泽湿地等混合景观的过程中,这里分布的繁育池内林蛙蛙卵团数较其它区域分布的蛙卵团数少,该类生境是导致林蛙卵团数与公路距离呈现曲线变化,在距公路较远的区域内繁育池内蛙卵团数量减少的主要因素(图 2)。
本研究在东北林蛙繁殖期开展生境因子对该物种产卵的影响,并将空间尺度划分为繁育池内微生境和繁育池外景观尺度,通过研究确定了在两个尺度环境因子对东北林蛙产卵的影响,这将有利于林蛙种群的繁育保护。此外,在林蛙保护的过程中,由于林蛙属于经济类动物,林区居民捕捉、农田开垦、公路建设对林蛙种群数量减少的影响都有待深入研究。
| [1] | Beebee T J C. Discriminant analysis of amphibian habitat determinants in South-East England. Amphibia-Reptilia, 1985, 6(1): 35-43. |
| [2] | Turner M G, Gardner R H, O'Neill R V. Landscape Ecology. New York, Berlin, Heidelberg: Springer and Verlag,2001. |
| [3] | Skidds D E, Golet F C, Paton P W C, Mitchell J C. Habitat correlates of reproductive effort in wood frogs and spotted salamanders in an urbanizing watershed. Journal of Herpetology, 2007, 41(3): 439-450. |
| [4] | Babbitt K J, Tanner G W. Use of temporary wetlands by anurans in a hydrologically modified landscape. Wetlands, 2000, 20(2): 313-322. |
| [5] | Egan R S, Paton P W C. Within-pond parameters affecting oviposition by wood frogs and spotted salamanders. Wetlands, 2004, 24(1): 1-13. |
| [6] | Skelly D K, Golon J. Assimilation of natural benthic substrates by two species of tadpoles. Herpetologica, 2003, 59(1): 37-42. |
| [7] | Ficetola G F, Marziali L, Rossaro B, De Bernardi F, Padoa-Schioppa E. Land-scape-stream interactions and habitat conservation for amphibians. Ecological Applications, 2011, 21(4): 1272-1282. |
| [8] | Porej D, Micacchion M, Hetherington T E. Core terrestrial habitat for conservation of local populations of salamanders and Wood Frogs in agricultural landscapes. Biological Conservation, 2004, 120(3): 399-409. |
| [9] | Hermann H L, Babbitt K J, Babe M J, Congalton R G. Effects of landscape characteristics on amphibian distribution in a forest-dominated landscape. Biological Conservation, 2005, 123(2): 139-149. |
| [10] | Rothermel B B, Semlitsch R D. An experimental investigation of landscape resistance of forest versus old-field habitats to emigrating juvenile amphibians. Conservation Biology, 2002, 16(5): 1324-1332. |
| [11] | Millspaugh J J, Marzluff J M. Radio Tracking and Animal Populations. San Diego: Academic Press, 2001. |
| [12] | Homan R N, Windmiller B S, Reed J M. Critical thresholds associated with habitat loss for two vernalpool-breeding amphibians. Ecological Applications, 2004, 14(5): 1547-1553. |
| [13] | Calhoun A J K, Klemens M W. Best development practices for conserving pool-breeding amphibians in residential and commercial developments in the Northeastern U. S. Metropolitan Conservation Alliance, Wildlife Conservation Society, MCA Technical Paper No. 5, Bronx, NY, 2002. |
| [14] | Pope S E, Fahig L, Merriam H G. Landscape complementation and metapopulation effects on Leopard Frog populations. Ecology, 2000, 81(9): 2498-2508. |
| [15] | Crouch W B, III, Paton P W C. Using egg mass counts to monitor wood frog populations. Wildlife Society Bulletin, 2000, 28(4): 895-901. |
| [16] | Hastie T J, Tibshirani R J. Generalized Additive Models. London: Chapman and Hall, 1990: 1-335. |
| [17] | 贾彬, 王彤, 王琳娜, 陈长生. 广义可加模型共曲线性及其在空气污染问题研究中的应用. 第四军医大学学报, 2005, 26(3): 280-283. |
| [18] | 欧阳芳,戈峰. 基于广义可加模型的昆虫种群动态非线性分析及R语言实现. 应用昆虫学报, 2013, 50(4): 1170-1177. |
| [19] | Ficetola G F, Adamo M, Bonardi A, Pasquale V D, Liuzzi C, Lovergine F, Marcone F, Mastropasqua F, Tarantino C, Blonda P, Padoa-Schioppa E. Importance of landscape features and Earth observation derived habitat maps for modelling amphibian distribution in the Alta Murgia National Park. International Journal of Applied Earth Observation and Geoinformation, 2015, 37: 152-159. |
| [20] | Seale D B. Physical factors influencing oviposition by the wood frog,Rana sylvatica, in Pennsylvania. Copeia, 1982: 627-635. |
| [21] | Todd B D, Luhring T M, Rothermel B B, Gibbons J W. Effects of forest removal on amphibian migrations: implications for habitat and landscape connectivity. Journal of Applied Ecology, 2009, 46(3): 554-561. |
| [22] | Guerry A D, Hunter M L Jr. Amphibian distributions in a landscape of forests and agriculture: an examination of landscape composition and configuration. Conservation Biology, 2002, 16(3): 745-754. |
| [23] | Carr L W, Fahrig L. Effect of road traffic on two amphibian species of differing vagility. Conservation Biology, 2001, 15(4): 1071-1078. |
| [24] | Fahrig L, Pedlar J H, Pope S E, Taylor P D, Wegner J F. Effect of road traffic on amphibian density. Biological Conservation, 1995, 73(1): 177-182. |