 2016, Vol. 36
2016, Vol. 36文章信息
- 姜瑛, 吴越, 徐莉, 胡锋, 李辉信
- JIANG Ying, WU Yue, XU Li, HU Feng, LI Huixin
- 悉生培养微缩体系食细菌线虫提高土壤激素含量的机制
- The possible mechanisms underlying improvement of soil auxin content by bacterial-feeding nematodes in a gnotobiotic microcosm experiment
- 生态学报, 2016, 36(9): 2528-2536
- Acta Ecologica Sinica, 2016, 36(9): 2528-2536
- http://dx.doi.org/10.5846/stxb201411102223
-
文章历史
- 收稿日期: 2014-11-10
- 网络出版日期: 2015-07-29






2. 河南农业大学资源与环境学院, 郑州 450002;
3. 山东省土壤肥料总站, 济南 250100
2. College of Resources and Environment, Henan Agricultural University, Zhengzhou 450002, China;
3. Soil and Fertilizer Station of Shandong Province, Jinan 250100, China
土壤食细菌线虫与土壤微生物之间存在密切的相互关系,土壤食细菌线虫除了自身的代谢活动作用外(如形成pH较高的微域环境、分泌排泄有机物),还可通过取食土壤细菌,影响微生物的数量、活性和周转从而参与土壤的生态过程[1, 2, 3, 4, 5, 6, 7, 8]。研究土壤食细菌线虫与细菌的相互作用及其生态功能是土壤生态学的核心研究内容之一,研究结果有助于揭示土壤生态过程及机理,丰富生态学理论。而在土壤中,食细菌线虫取食植物促生菌也一定会对其数量以及活性等造成一定的影响。
悉生培养(gnotobitic microcosm culture)是指将供试土壤在密闭的培养系统中灭菌,然后在严格控制的无菌培养条件下引入供试目标土壤线虫和纯培养的微生物进行培养[9]。悉生培养系统可以将复杂的生态系统简化,实现目标生物和环境的可控性,是研究土壤生物之间相互作用的有力工具。自从Aderson以及Coleman等人率先利用该系统研究食细菌线虫与微生物的相互作用,以及对土壤生态系统能量传递和养分动态的影响以来[10, 11],该领域受到了广泛关注[9, 12, 13, 14, 15],南京农业大学土壤生态实验室也在悉生培养系统中对食细菌线虫与微生物之间的相互作用进行了大量的研究[16, 17, 18]。
前期研究通过悉生以及原位富化等试验发现食细菌线虫的取食改变了番茄、小麦以及水稻根际微生物群落结构的同时,使得土壤中生长素(Indole acetic acid,IAA)含量显著增加[19, 20, 21, 22],因此,食细菌线虫有可能是通过刺激土壤中一些植物促生菌分泌大量的激素以及类激素物质从而提高土壤中生长素IAA的含量[4]。但是目前还没有直接的证据表明土著食细菌线虫对土著产IAA细菌的促进作用。
依据线虫在生活史策略连续谱(r-对策到K-对策)中的位置,可将其划分为5个不同的cp(colonizer-persister)类群[23]。其中cp值越低,线虫产卵量越多,世代时间越短,因此不同cp值线虫在生态系统中的作用存在差异。本文就采用简单较易控制条件的悉生培养系统,对比研究两种不同cp值(cp1和cp2)的土著食细菌线虫与土著产IAA细菌的相互作用,为进一步明确土壤食细菌线虫取食细菌所产生的激素效应提供切实可靠的依据。
1 材料与方法 1.1 实验材料 1.1.1 供试土壤采自江苏省南京市雨花台区板桥镇长江南岸冲积地的灰潮土,美国制土壤质地分类为砂质壤土。采集表层0—20cm的土样,采回新鲜土样之后,剔除土壤中肉眼可见的根茬等残体和蚯蚓等大中型土壤动物以及小石块等,风干并且过2mm筛,备用。土壤的基本性状如表 1所示。
| 有机碳Organic C/(g/ kg) | 全氮Total N/(g/ kg) | 铵态氮NH+4-N/(mg/kg) | 硝态氮NO-3-N/(mg/kg) | 矿质氮Mineral N/(mg/kg) | pH(H2O) |
| 9.20 | 0.89 | 6.84 | 2.39 | 9.23 | 6.32 |
两种土著优势食细菌线虫:头叶属(Cephalobus sp.)(cp2),中杆属(Mesorhabditis sp.)(cp1),均筛自供试土壤,保存在22℃恒温培养箱中。
1.1.3 供试菌株两种土著细菌:产IAA细菌解淀粉芽孢杆菌Bacillus amyloliquefaciens (GenBank accession number JX424611)和对照不产IAA的细菌贪噬菌Variovorax sp. (GenBank accession number JX424612),均筛自供试土壤,保存在4℃冰箱中。
1.1.4 培养基线虫培养基(Nematode growth media,NGM):2.5 g 蛋白胨,3g NaCl和17g 琼脂溶于1L蒸馏水中,高压灭菌,加入已通过无菌滤膜的 1mL 1mol/L CaCl2,1mL 1mol/L MgSO4,1mL 5mg/mL 胆固醇和25mL 1mol/L KHPO4。
LB培养基:蛋白胨10g,酵母提取物5g,氯化钠10g,琼脂20g,蒸馏水1000mL,调节pH 7.0—7.2,121℃灭菌20min。
1.2 细菌与线虫的预培养 1.2.1 细菌培养将冰箱中预先准备好的解淀粉芽孢杆菌Bacillus amyloliquefaciens (B) 和贪噬菌Variovorax sp. (V) 的菌悬液接种到NGM液体培养基内,28℃培养2d,5000 r/min离心5 min,收集离心管底部菌体,用无菌水反复洗涤离心3次后重新悬浮于无菌水中,制成约107 cfu/mL的菌悬液,待用。
1.2.2 线虫的富化培养采用单种培养法,将两种供试线虫头叶属Cephalobus sp. (C)或中杆属Mesorhabditis sp. (M)分别接种到长有单一解淀粉芽孢杆菌Bacillus amyloliquefaciens (B) 或者贪噬菌Variovorax sp. (V)的NGM固体培养基上,采用处理中的组合一一对应,于22℃富化培养。当线虫培养至达到实验所需的数量时采用浅盘法[24]分离,分离得到的线虫放置于灭菌的离心管中,离心去上清液,用无菌水重复清洗5—6次,待用。
1.3 试验设计 1.3.1 处理设置本试验采用悉生培养法,为了尽可能少地破坏土壤的理化性质,采用60Co-γ射线灭菌(25kGy)(江苏省农业科学研究院辐照中心进行灭菌)。
本试验共设置7个处理,每个处理设置3次重复:
1)灭菌土 (CK); 2)灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens (B); 3)灭菌土+B+头叶属食细菌线虫Cephalobus sp. (BC); 4)灭菌土+B+中杆属食细菌线虫Mesorhabditis sp. (BM); 5)灭菌土+对照菌贪噬菌Variovorax sp. (V); 6)灭菌土+V+头叶属食细菌线虫Cephalobus sp. (VC); 7)灭菌土+V+中杆属食细菌线虫Mesorhabditis sp. (VM)。
1.3.2 试验步骤将装有80g无菌供试土壤的100mL小烧杯按处理接种细菌,接种量为106 cfu/g干土,于28℃预培养2d后再按处理分别均匀接入对应的两种食细菌线虫悬液,使每克干土中接入的线虫数达到大约30条,然后补充土壤含水量到田间持水量的60%,即实际含水量为24.8%左右,此含水量条件使得食细菌线虫对细菌的促进作用达到最强。用无菌封口膜封住小烧杯口,22℃下恒温培养,分别于接种食细菌线虫后的第10、20、30天破坏性采样,采样时每个处理取3个重复测定相应指标。
1.4 测定指标与方法 1.4.1 土壤含水量烘干称重法,在105℃条件下烘干8h。差值法计算含水量。
1.4.2 线虫分离与计数采用浅盘法对线虫进行分离,称取10g土样,加入浅盘中,在室温下分离48h之后,用两个500目的网筛收集线虫悬液到划有网格的小皿中,在体视显微镜下对线虫进行记数。
1.4.3 土壤中细菌数量的计数采用稀释涂布平板法用LB固体培养基培养细菌并计数。
准确称取待测土样5.00g,放入装有45mL 无菌水并放有小玻璃珠的150mL 三角瓶中,置摇床上振荡30min,使微生物细胞分散,静置20—30s,即成10-1 稀释液;连续稀释,制成10-2、10-3、10-4、10-5、10-6、10-7等一系列稀释菌液。选择10-5、10-6、10-7这3个稀释梯度待涂布。采用涂布平板法,先将LB固体培养基高温灭菌,取出稍凉后倒入灭菌平板中,待凝固后编号,然后用无菌吸管吸取0.1 mL 含菌土壤稀释液对号接种在相应稀释梯度编号的平板上(每个编号设3个重复),再用无菌的涂布器将菌液在平板上涂抹均匀,将涂抹好的平板平放于桌上20—30min,使菌液渗透入培养基内,之后将平板倒转,在28 ℃恒温培养箱中培养48h后计数。
1.4.4 土壤基础呼吸准确称取待测土样15.00g,放入250mL玻璃瓶中,塞紧橡胶塞防止漏气,密闭培养6h后用注射器采集气体样品,用气相色谱仪7890A GC system (Agilent Technologies,USA)中的FID检测器来测定二氧化碳的释放通量。工作条件:柱温为60℃,检测器温度为300℃,H2流量为40mL/min,空气流量为300mL/min,FID检测器的载气为N2(2mL/min),色谱柱为APPQ-6FT填充柱。计算公式如下:
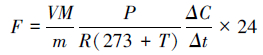
称取土壤样品10 g到100 mL离心管,加入80%丙酮水溶液50 mL,在冰浴条件下超声提取30min,4000r/min离心10min,将上清液转入旋转蒸发器内,减压浓缩(40℃),将丙酮蒸发完全(即没有冷凝液下滴),用0.1mol/L盐酸调pH值为2.8—3.0,再用乙酸乙酯萃取3次,合并有机相,减压浓缩(40℃)蒸干,用5 mL 色谱纯甲醇溶解后,过0.45μm有机相滤膜,滤液保存在棕色小瓶中,用高效液相色谱(HPLC)测定。
色谱条件为流动相甲醇:水:醋酸=50:45:5;检测波长254 nm;流速0.7 mL/min;进样量20μL;色谱柱C18(uBondapak,10u,300×3.9mm)。
1.5 数据统计方法数据统计采用SPSS16.0统计软件,用 Duncan法分析不同处理间的平均值差异(P<0.05),用三因素方差(Three-Way ANOVA)分析在整个培养时间(Time)(10、20、30d)内两种接种细菌(Bacteria)(产IAA细菌和对照菌)以及两种食细菌线虫(Nematode)对细菌数量和活性(处理B、BC、BM、V、VC、VM)以及土壤IAA含量(处理CK、B、BC、BM、V、VC、VM)的影响;作图采用OriginPro 8.0 作图软件。
2 结果与分析 2.1 土壤中食细菌线虫数量的变化特征由表 2可知,在所有添加食细菌线虫的处理中,食细菌线虫都能够得到很好的生存和繁殖,线虫数量先升高,到第20天的时候达到最大值,为初始线虫的2.17—3.12倍,到第30天时线虫数量有所降低,但依然能达到初始线虫的1.61—1.89倍。添加产激素菌Bacillus amyloliquefaciens和中杆属食细菌线虫Mesorhabditis sp.的处理(BM)线虫数在培养时期内均高于添加产激素菌Bacillus amyloliquefaciens和头叶属食细菌线虫Cephalobus sp.的处理(BC),但是差异均没有达到显著。而在培养第10天和第20天的时候添加对照菌Variovorax sp.和头叶属食细菌线虫Cephalobus sp.的处理(VC)线虫数量显著高于添加对照菌Variovorax sp.和中杆属食细菌线虫Mesorhabditis sp.的处理(VM)。显然,在BM和VC的处理中,线虫存活较好,这可能与食细菌线虫的取食偏好有关。
| 处理Treatments | 线虫数量(条/g干土) The number of nematodes | ||
| 10 d | 20 d | 30 d | |
| BC | 41.75±7.57b | 72.71±12.77b | 48.44±4.26a |
| BM | 48.09±12.35b | 83.76±5.19ab | 51.99±6.84a |
| VC | 67.76±11.25a | 99.51±7.22a | 56.70±10.84a |
| VM | 41.16±3.86b | 64.98±13.89b | 50.83±5.12a |
| 同一列内字母不同表示有显著性差异(P <0.05);BC:灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens +头叶属食细菌线虫Cephalobus sp.; BM:灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens +中杆属食细菌线虫Mesorhabditis sp.; VC:灭菌土+对照菌贪噬菌Variovorax sp.+头叶属食细菌线虫Cephalobus sp.; VM:灭菌土+对照菌贪噬菌Variovorax sp.+中杆属食细菌线虫Mesorhabditis sp. | |||
方差分析结果表明:接种不同种的细菌(Bacteria)对细菌数量和活性并没有显著的影响,而接种不同种的线虫(Nematode)和培养时间(Time)均显著地(P<0.05)影响了细菌的数量和活性,但是它们三者之间并不存在显著的交互作用(表 3)。
| 变异来源Source of variation | df | F | df | F | |
| IAA含量IAA content | 细菌数量BNBacteria number | 基础呼吸BRBasal respiration | |||
| *显著性差异检验Test significant at the 5% level (P<0.05); NS: 无显著性差异检验 | |||||
| 细菌Bacteria (B) | 2 | 2381.97* | 1 | 0.24 NS | 1.71 NS |
| 线虫Nematode (N) | 2 | 17.84* | 2 | 12.60* | 3.89* |
| 培养时间Time (T) | 2 | 104.79* | 2 | 32.64* | 5.01* |
| B × N | 2 | 20.07* | 2 | 3.21 NS | 0.31 NS |
| B × T | 4 | 55.19* | 2 | 0.29 NS | 0.09 NS |
| N × T | 4 | 0.66 NS | 4 | 0.75 NS | 0.23 NS |
| B × N × T | 4 | 1.30 NS | 4 | 0.32 NS | 0.10 NS |
| 误差均方Error mean squares | 42 | 0.00 | 36 | 2.76 | 4.75 |
接种不同种的食细菌线虫对产IAA细菌以及对照菌数量和活性的影响如图 1、图 2所示。在整个培养期内,所有处理的细菌数量(图 1)和活性(图 2)均表现出先升高后降低的趋势,土壤中接种两种食细菌线虫的处理(BC,BM,VC,VM)的细菌数量和活性均高于单独接种产IAA细菌(B)和对照不产IAA菌(V)的处理。不同食细菌线虫和细菌在不同培养期对细菌数量和活性的影响之间存在差异,由图 1可知,在第10天和第20天,BM处理的细菌数量均显著高于单接产IAA菌的B处理,BC处理虽然也高于B处理,但并不显著。而第10天和第30天,VC处理的细菌数量均显著高于单接对照菌的V处理,VM处理虽然也高于V处理,但是也没有达到显著水平;由图 2可知,虽然接种食细菌线虫均提高了两种细菌的活性,但是在3次采样以及相对应的处理之间并没有达到显著水平,只有在第10天的时候,VC处理的细菌活性最高,显著高于单独接种产IAA菌的处理B。

|
| 图 1 食细菌线虫对土壤细菌数量的影响 Fig. 1 Effects of bacteria-feeding nematodes on the number of bacteria during the incubation time. Error bars represent standard deviation. For each incubation time, significant differences (P<0.05, n=3) are indicated by different letters. |

|
| 图 2 食细菌线虫对土壤呼吸的影响 Fig. 2 Effect of bacteria-feeding nematodes on soil respiration during the incubation time. Error bars represent standard deviation (For each incubation time, significant differences (P<0.05, n=3) are indicated by different letters) B:灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens;BC:灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens +头叶属食细菌线虫Cephalobus sp.;BM:灭菌土+产IAA菌解淀粉芽孢杆菌Bacillus amyloliquefaciens +中杆属食细菌线虫Mesorhabditis sp.;V: 灭菌土+对照菌贪噬菌Variovorax sp.;VC:灭菌土+对照菌贪噬菌Variovorax sp.+头叶属食细菌线虫Cephalobus sp.;VM:灭菌土+对照菌贪噬菌Variovorax sp.+中杆属食细菌线虫Mesorhabditis sp.;CFU (colony-forming units) |
方差分析结果表明:接种两种不同的细菌(Bacteria)、两种不同的食细菌线虫(Nematode)以及整个培养时间(Time)均显著影响了土壤中IAA的含量,同时,细菌和食细菌线虫二者之间的交互、细菌和培养时间二者之间的交互显著影响土壤中IAA的含量(表 3)。
接种不同种的食细菌线虫与产IAA细菌和对照菌相互作用对土壤中IAA含量的影响如图 3所示,灭菌土本身(CK)的IAA含量并不高,且在整个培养过程中变化不大,并且在接种不产IAA的对照菌的处理中,不管是否接种食细菌线虫,对土壤中IAA的含量均没有显著的影响。对比对照处理以及接种对照菌的各个处理,接种产IAA细菌的处理不管是否接种食细菌线虫,均显著增加了土壤中的IAA含量。相对于单独接种产IAA细菌的处理B来说,接种两种食细菌线虫均提高了土壤中IAA的含量,而两种不同的食细菌线虫对产IAA细菌产IAA量的影响也有差异,在第10天时,与B处理相比,BM处理显著提高了IAA的含量,BC处理虽然也提高了IAA含量,但是差异并没有达到显著;到第20天时,BM和BC处理的IAA含量均显著高于B处理的,但是BM和BC处理之间的差异并不显著;到第30天时,BM和BC处理的IAA含量均显著高于B处理的同时,BM处理的土壤IAA含量也要显著高于BC处理。

|
| 图 3 食细菌线虫对土壤IAA含量的影响 Fig. 3 Effects of bacterial-feeding nematodes on soil IAA content during the incubation time. Error bars represent standard deviation. For each incubation time, significant differences (P<0.05, n=3) are indicated by different letters IAA:吲哚乙酸 indole acetic acid |
本文通过悉生培养系统,探讨了两种不同cp值的食细菌线虫对产IAA细菌的数量、活性、分泌IAA的能力等的影响。通过研究发现,接种食细菌线虫的所有处理之间的线虫数虽然存在差异,但是它们在整个培养时期内的线虫数均高于初始接种的30条/g干土的数量,而所有处理在整个培养期内,不管有没有接种食细菌线虫,细菌的数量均高于初始接种的106CFU/g干土,这可能是由于本实验所用的供试材料均是筛自供试土壤,土著的细菌和线虫都很适应原土中的环境,在原土中都具有良好的生存和定殖能力,并且筛选的产IAA细菌和对照不产IAA的细菌都可以被这两种食细菌线虫取食并且影响到线虫的繁殖。而在BM和VC处理中的食细菌线虫数量要多于其他处理,这可能与食细菌线虫的取食偏好性有关[7, 25],Djigal等[26]也通过悉生培养实验发现4种不同的细菌对食细菌线虫Zeldia punctata繁殖能力的影响不同,这与本实验得到了相同的结果,肖海峰在对模式线虫Caenorhabditis elegans研究后提出,线虫的繁殖率与其取食的偏好性是相关的[27]。
食细菌线虫的取食活动对细菌的数量和活性都会造成一定的影响,本实验研究发现,接种两种不同的食细菌线虫均可以增加细菌的数量,然而,有研究表明,线虫的过度取食也会造成细菌数量的减少,而线虫的过度取食会掩盖其对细菌数量的促进作用[9, 28, 29],食细菌线虫与细菌的数量之间存在着一种动态平衡的关系,因此本实验中细菌的数量也随着培养时间的延长存在着明显的上下波动的趋势。
相对于细菌数量来说,细菌活性的测定更能反映食细菌线虫的取食对细菌的真实影响[30],并且在大量的研究中,线虫对细菌活性(及CO2的释放强度)的影响相比细菌数量更加具备一致的明显的趋势。在研究食细菌线虫对细菌活性的影响时,我们不得不考虑的问题是,食细菌线虫与细菌一样也有呼吸,那么这个呼吸的量是否会影响我们对结果的分析呢?研究发现,食细菌线虫的生物量较少,通常还要少于原生动物,因此其对土壤呼吸的贡献率通常只有1%甚至更少[31]。在本实验中,接种食细菌线虫提高了细菌的活性,这与以往大量的研究结果是一致的[32, 33, 34, 35]。食细菌线虫对细菌数量和活性的促进作用可能是因为以下几个方面:线虫的取食过程分泌出了可供细菌利用的养分,线虫摄食的细菌在线虫的肠道中获得了某些营养物质,被排出后反而加快了生长,并且线虫能将细菌的数量控制在相对较低但是已经足够的水平,减少了基质的限制作用,使得细菌能够保持在对数生长期的高活性水平,同时线虫也具有较强的移动能力,可以将细菌传播到细菌本身很难迅速到达的营养物质丰富的地方[36, 37, 38]。同时发现,接种不同生活史策略(cp值)的食细菌线虫对不同种细菌的数量和活性的影响不同,这与Djigal 2004年的研究结果是一致的[26],他的研究表明相比较A. nanus 和 C. pseudoparvus.这两种线虫,Z. punctata 对微生物的数量及活性的影响均较弱。研究发现,线虫对细菌的影响会因线虫的种类、代谢速率和繁殖速率的不同而不同[3, 23, 39, 40],不同生活史策略的线虫对细菌生长也有不同的影响[41]。同时,食细菌线虫与细菌之间的相互作用也随着细菌种类的不同而不同[29, 34, 42, 43, 44]。
而食细菌线虫在促进了细菌的增殖以及活性的过程中,也影响到了细菌代谢产物的种类和数量,在本实验中,接种食细菌线虫增加了产IAA细菌的数量,促进了产IAA细菌的活性,同时我们发现,土壤中的IAA含量也得到了显著的增加,目前还没有报道称食细菌线虫本身能够分泌生长素IAA,而通过研究发现接种对照不产IAA菌和食细菌线虫相互作用对土壤中的IAA含量没有任何影响,更加肯定了食细菌线虫本身是不能够分泌IAA的,因此土壤中IAA含量的增加是由于土壤中产IAA细菌数量和活性增强的结果。有报道称,食细菌线虫比细菌的移动能力更强,这种移动能力可以促进植物促生菌在根际的定殖,而食细菌线虫的取食促进了植物促生菌的增殖和活性,并在这个过程中刺激了细菌代谢物的增多,比如一些有利于根系生长的激素类物质[45, 46],本实验结果也显示,接种食细菌线虫与产IAA细菌相互作用显著增加了土壤中IAA的含量,并且不同种食细菌线虫对IAA含量影响的验证结果也均达到了显著水平。由此,可以得出,土壤食细菌线虫是通过提高土壤中产IAA细菌的数量和活性,刺激其产生更多代谢产物,从而提高土壤中激素含量的。
4 小结食细菌线虫的繁殖能力因其取食细菌种类的不同而不同,cp1的中杆属食细菌线虫与产IAA细菌的组合更能促进线虫数量的增殖;两种食细菌线虫的取食均能促进细菌数量和活性的增强,食细菌线虫与产IAA细菌相互作用也均能显著增加土壤中IAA的含量,对照菌处理则对IAA含量没有影响。这些促进作用也都随着接种细菌和食细菌线虫种类以及培养时间的变化而变化,接种cp1的中杆属食细菌线虫比cp2的头叶属食细菌线虫更能有效地促进产IAA细菌的数量、活性从而提高土壤中IAA的含量,这可能与中杆属食细菌线虫cp值低,产卵量多,周转速率更快,同时也与食细菌线虫的取食偏好性有关。
| [1] | Bouwman L A, Bloem J, van den Boogert P H J F, Bremer F, Hoenderboom G H J, de Ruiter P C. Short-term and long-term effects of bacterivorous nematodes and nematophagous fungi on carbon and nitrogen mineralization in microcosms. Biology and Fertility of Soils, 1994, 17(4): 249-256. |
| [2] | Alphei J, Bonkowski M, Scheu S. Protozoa, Nematoda and Lumbricidae in the rhizosphere of Hordelymus europeaus (Poaceae): faunal interactions, response of microorganisms and effects on plant growth. Oecologia, 1996, 106(1): 111-126. |
| [3] | Ferris H, Venette R C, Van Der Meulen H R, Lau S S. Nitrogen mineralization by bacterial-feeding nematodes: verification and measurement. Plant and Soil, 1998, 203(2): 159-171. |
| [4] | Bonkowski M, Cheng W X, Griffiths B S, Alphei J, Scheu S. Microbial-faunal interactions in the rhizosphere and effects on plant growth1. European Journal of Soil Biology, 2000, 36(3/4): 135-147. |
| [5] | Chen J, Ferris H. Growth and nitrogen mineralization of selected fungi and fungal-feeding nematodes on sand amended with organic matter. Plant and Soil, 2000, 218(1/2): 91-101. |
| [6] | Savin M C, Görres J H, Neher D A, Amador J A. Uncoupling of carbon and nitrogen mineralization: role of microbivorous nematodes. Soil Biology and Biochemistry, 2001, 33(11): 1463-1472. |
| [7] | Salinas K A, Edenborn S L, Sexstone A J, Kotcon J B. Bacterial preferences of the bacterivorous soil nematode Cephalobus brevicauda (Cephalobidae): Effect of bacterial type and size. Pedobiologia, 2007, 51(1): 55-64. |
| [8] | Dos Santos G A P, Derycke S, Fonsêca-Genevois V G, Coelho L C B B, Correia M T S, Moens T. Differential effects of food availability on population growth and fitness of three species of estuarine, bacterial-feeding nematodes. Journal of Experimental Marine Biology and Ecology, 2008, 355(1): 27-40. |
| [9] | Ingham R E, Trofymow J A, Ingham E R, Coleman D C. Interactions of bacteria, fungi, and their nematode grazers: effects on nutrient cycling and plant growth. Ecological Monographs, 1985, 55(1): 119-140. |
| [10] | Anderson R V, Elliott E T, McClellan J F, Coleman D C, Cole C V, Hunt H W. Trophic interactions in soils as they affect energy and nutrient dynamics. III. Biotic interactions of bacteria, amoebae, and nematodes. Microbial Ecology, 1977, 4(4): 361-371. |
| [11] | Coleman D C, Cole C V, Hunt H W, Klein D A. Trophic interactions in soils as they affect energy and nutrient dynamics. I. Introduction. Microbial Ecology, 1977, 4(4): 345-349. |
| [12] | Abrams B I, Mitchell M J. Role of nematode-bacterial interactions in heterotrophic systems with emphasis on sewage sludge decomposition. Oikos, 1980, 35(3): 404-410. |
| [13] | Anderson R V, Coleman D C, Cole C V, Elliott E T. Effect of the nematodes Acrobeloides sp. and Mesodiplogaster lheritieri on substrate utilization and nitrogen and phosphorous mineralization in soil. Ecology, 1981, 62(3): 549-555. |
| [14] | Anderson R V, Coleman D C, Cole C V, Elliott E T, McClellan J F. The use of soil microcosms in evaluating bacteriophagic nematode responses to other organisms and effects on nutrient cycling. International Journal of Environmental Studies, 1979, 13(2): 175-182. |
| [15] | Sundin P, Valeur A, Olsson S, Odham G. Interactions between bacteria-feeding nematodes and bacteria in the rape rhizosphere: effects on root exudation and distribution of bacteria. FEMS Microbiology Ecology, 1990, 6(1): 13-22. |
| [16] | 毛小芳, 胡锋, 陈小云, 李辉信. 不同土壤水分条件下华美新小杆线虫对枯草芽孢杆菌数量、活性及土壤氮素矿化的影响. 应用生态学报, 2007, 18(2): 405-410. |
| [17] | 陈小云, 胡锋, 李辉信, 毛小芳. 不同悉生培养条件下食细菌线虫对细菌种群的影响. 应用生态学报, 2003, 14(9): 1585-1587. |
| [18] | 毛小芳, 李辉信, 龙梅, 胡锋. 不同食细菌线虫取食密度下线虫对细菌数量、活性及土壤氮素矿化的影响. 应用生态学报, 2005, 16(6): 1112-1116. |
| [19] | Cheng Y H, Jiang Y, Griffiths B S, Li D M, Hu F, Li H X. Stimulatory effects of bacterial-feeding nematodes on plant growth vary with nematode species. Nematology, 2011, 13(3): 369-372. |
| [20] | Mao X F, Hu F, Griffiths B, Chen X Y, Liu M Q, Li H X. Do bacterial-feeding nematodes stimulate root proliferation through hormonal effects?. Soil Biology and Biochemistry, 2007, 39(7): 1816-1819. |
| [21] | Mao X F, Hu F, Griffiths B, Li H X. Bacterial-feeding nematodes enhance root growth of tomato seedlings. Soil Biology and Biochemistry, 2006, 38(7): 1615-1622. |
| [22] | Jiang Y, Wu Y, Xu W S, Cheng Y H, Chen J D, Xu L, Hu F, Li H X. IAA-producing bacteria and bacterial-feeding nematodes promote Arabidopsis thaliana root growth in natural soil. European Journal of Soil Biology, 2012, 52: 20-26. |
| [23] | Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19. |
| [24] | Goodfriend W L, Olsen M W, Frye R J. Soil microfloral and microfaunal response to Salicornia bigelovii planting density and soil residue amendment. Plant and Soil, 2000, 223(1/2): 23-32. |
| [25] | Newsham K K, Rolf J, Pearce D A, Strachan R J. Differing preferences of Antarctic soil nematodes for microbial prey. European Journal of Soil Biology, 2004, 40(1): 1-8. |
| [26] | Djigal D, Sy M, Brauman A, Diop T A, Mountport D, Chotte J L, Villenave C. Interactions between Zeldia punctata (Cephalobidae) and bacteria in the presence or absence of maize plants. Plant and Soil, 2004, 262(1/2): 33-44. |
| [27] | 肖海峰, 焦加国, 胡锋, 李辉信. 食细菌线虫 Caenorhabditis elegans 的取食偏好性. 生态学报, 2010, 30(24): 7101-7105. |
| [28] | Griffiths B S, Ritz K, Wheatley R E. Nematodes as indicators of enhanced microbiological activity in a Scottish organic farming system. Soil Use and Management, 1994, 10(1): 20-24. |
| [29] | Venette R C, Ferris H. Influence of bacterial type and density on population growth of bacterial-feeding nematodes. Soil Biology and Biochemistry, 1998, 30(7): 949-960. |
| [30] | Djigal D, Brauman A, Diop T A, Chotte J L, Villenave C. Influence of bacterial-feeding nematodes (Cephalobidae) on soil microbial communities during maize growth. Soil Biology and Biochemistry, 2004, 36(2): 323-331. |
| [31] | Sohlenius B. Abundance, biomass and contribution to energy flow by soil nematodes in terrestrial ecosystems. Oikos, 1980, 34(2): 186-194. |
| [32] | Coleman D C, Reid C P P, Cole C V. Biological strategies of nutrient cycling in soil systems. Advances in Ecological Research, 1983, 13: 1-55. |
| [33] | Woods L E, Cole C V, Elliott E T, Anderson R V, Coleman D C. Nitrogen transformations in soil as affected by bacterial-microfaunal interactions. Soil Biology and Biochemistry, 1982, 14(2): 93-98. |
| [34] | Fu S L, Ferris H, Brown D, Plant R. Does the positive feedback effect of nematodes on the biomass and activity of their bacteria prey vary with nematode species and population size?. Soil Biology and Biochemistry, 2005, 37(11): 1979-1987. |
| [35] | Mikola J, Setälä H. Productivity and trophic-level biomasses in a microbial-based soil food web. Oikos, 1998, 82(1): 158-168. |
| [36] | 胡锋, 李辉信, 谢涟琪, 吴珊眉. 土壤食细菌线虫与细菌的相互作用及其对 N, P 矿化-生物固定的影响及机理. 生态学报, 1999, 19(6): 914-920. |
| [37] | Griffiths B S, Bardgett R D. Interactions between microbe-feeding invertebrates and soil microorganisms// van Elsas J D, Trevors J T, Wellington E M H, eds. Modern Soil Microbiology. New York: CRC Press, 1997: 165-182. |
| [38] | Mamilov A S, Byzov B A, Zvyagintsev D G, Dilly O M. Predation on fungal and bacterial biomass in a soddy-podzolic soil amended with starch, wheat straw and alfalfa meal. Applied Soil Ecology, 2001, 16(2): 131-139. |
| [39] | Ferris H, Venette R C, Lau S S. Population energetics of bacterial-feeding nematodes: carbon and nitrogen budgets. Soil Biology and Biochemistry, 1997, 29(8): 1183-1194. |
| [40] | Blanc C, Sy M, Djigal D, Brauman A, Normand P, Villenave C. Nutrition on bacteria by bacterial-feeding nematodes and consequences on the structure of soil bacterial community. European Journal of Soil Biology, 2006, 42(S1): S70-S78. |
| [41] | Mikola J, Setälä H. Relating species diversity to ecosystem functioning: mechanistic backgrounds and experimental approach with a decomposer food web. Oikos, 1998, 83(1): 180-194. |
| [42] | Ferris H, Venette R C, Scow K M. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Applied Soil Ecology, 2004, 25(1): 19-35. |
| [43] | Marschner P, Yang C H, Lieberei R, Crowley D E. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biology & Biochemistry, 2001, 33(11): 1437-1445. |
| [44] | Grewal P S. Influence of bacteria and temperature on the reproduction of caenorhabditis elegans (Nematoda: Rhabditidae) infesting mushrooms (Agaricus Bispor Us). Nematologica, 1991, 37(1/4): 72-82. |
| [45] | Knox O G G, Killham K, Artz R R E, Mullins C, Wilson M. Effect of nematodes on rhizosphere colonization by seed-applied bacteria. Applied and Environmental Microbiology, 2004, 70(8): 4666. |
| [46] | Knox O G G, Killham K, Mullins C E, Wilson M J. Nematode-enhanced microbial colonization of the wheat rhizosphere. FEMS Microbiology Letters, 2003, 225(2): 227-233. |