 2016, Vol. 36
2016, Vol. 36文章信息
- 陈丹梅, 段玉琪, 杨宇虹, 晋艳, 袁玲
- CHEN Danmei, DUAN Yuqi, YANG Yuhong, JIN Yan, YUAN Ling
- 轮作模式对植烟土壤酶活性及真菌群落的影响
- Influence of crop rotation on enzyme activities and fungal communities in flue- cured tobacco soil
- 生态学报, 2016, 36(8): 2373-2381
- Acta Ecologica Sinica, 2016, 36(8): 2373-2381
- http://dx.doi.org/10.5846/stxb201411052183
-
文章历史
- 收稿日期: 2014-11-05
- 网络出版日期: 2015-08-24






2. 云南省烟草农业科学研究院, 昆明 650031
2. Yunnan Tobacco Agriculture Research Institute, Kunming 650031, China
同一作物或近缘作物长期连续种植会造成土壤养分异常积累或过度消耗,病原微生物迅速繁衍,化感物质积累,土壤微生物种群结构失衡,作物产量品质降低和病害严重发生等[1]。因此,多种作物轮作辅以休闲一般优于种植单一作物,研究不同轮作模式对土壤物理、化学、生物学性质的影响,可为合理轮作提供科学依据。
在我国热带和亚热带地区,通常采用一年三熟或两年五熟的高强度土壤利用模式,需要大量施用化肥,导致土壤有机质减少,结构破坏,病害严重[2]。马铃薯长期连作后,根际土壤中的细菌数显著低于其他种植模式,但真菌则相反,作物发生真菌病害的风险提高[3]。玉米连作年限不同,根际微生物的数量、种群和多样性也不一样。随着连作时间延长,它们的数量和种群减少,多样性逐渐降低[4]。烤烟长期连作,提高黑胫病和青枯病的发病率,降低烟叶产量和质量[5]。相反,轮作则能有效中断病原菌寄主,降低作物病害发生率[5, 6, 7]。值得注意的是,轮作模式不同,对土壤物理、化学、生物学性质的影响也不一样。连续5a实施水稻-油菜轮作,土壤有机质含量显著高于水稻-小麦和水稻-黑麦草轮作[8]。在我国东北黑土上,实施黄瓜连作、小麦-黄瓜和毛苕子-黄瓜轮作,轮作显著增加根际细菌种类,但减少结瓜后期的真菌种群,尤以毛苕子-黄瓜轮作最为显著[9]。微生物是土壤的重要组成部分,释放多种酶类,参与土壤有机质降解、腐殖质合成和养分循环等生物化学过程[10]。土壤微生物数量和种群随种植植物的种类不同而发生变化,作物根系分泌物和凋落物对土壤微生物的种群结构产生重要影响[11],并与土传病害密切相关[12]。
在微生物生态学研究中,利用Roche Genome Sequencer FLX测序平台进行454高通量测序是重要的分子生物学手段。该方法通过测定微生物保守性较高的16S rDNA和18S rDNA序列,并与基因库中的已知序列进行对比,能准确、快速、灵敏地确定待测微生物属种。自454高通量测序技术应用以来,已检测出数百至数千种土壤微生物,是传统培养方法所获得微生物数量的数十倍甚至数百倍。此外,该方法还能检测出不可培养的微生物[13]。云南省是我国烤烟主产区,烤烟轮作比例达80%以上,烟地夏季主要轮作玉米和水稻,分别占种植面积的64.7%和27.6%,冬季一般种植油菜和绿肥或休闲[14]。选择科学的轮作模式有益于减轻连作障碍,是保持土地生产力和保障整个农业生产健康和持续发展的重要措施[15]。因此,本研究以云南省烟草农业科学研究院的长期轮作试验为对象,采用化学、酶学方法及454高通量测序技术,揭示了云南烟区主要轮作模式对作物产量和产值、土壤养分、酶活性及真菌种群结构的影响,为合理选择烤烟轮作模式奠定基础。
1 材料与方法 1.1 试验地概况云南省烟草农业科学研究院研究和试验基地位于北纬24°14′,东经 102°30′,海拔1680 m,年均温度15.9℃,年降雨量918mm,雨季(4—9月)降雨量占全年的79.5%,年日照时数2072h。供试土壤为云南省典型、具有代表性的砂质红壤,基础土壤的pH6.4,有机质10.70 g/kg,全氮0.54 g/kg,全磷0.11 g/kg,全钾6.43 g/kg,有效氮82.0 mg/kg,有效磷9.01 mg/kg,有效钾160.0 mg/kg。
1.2 试验处理试验始于1998年,设置云南普遍采用的4种种植模式,包括烤烟-休闲-玉米(T-B-M)、烤烟-油菜-玉米(T-C-M)、烤烟-油菜-水稻(T-C-R)和烤烟-苕子-水稻(T-V-R)等。小区面积28m2,重复3次,随机区组排列。烤烟、玉米和水稻一般在5月初移栽,苕子和油菜秋末播种。根据当地大田生产情况,烤烟施用纯氮105kg/hm2,P2O5 74 kg/hm2,K2O 263 kg/hm2;油菜施用纯氮90kg/hm2,P2O5 30 kg/hm2;玉米和水稻施用纯氮112.5kg/hm2,P2O5 40 kg/hm2;苕子施用纯氮75kg/hm2。此外,水稻、玉米和油菜秸秆全部切碎还田;绿肥收割多次,未彻底收割的残桩全部翻压入土。在烤烟种植季节,基肥由N∶P2O5∶K2O=10∶10∶25烤烟专用肥提供,施氮量占施肥总量的70%;剩余30%的氮素由烤烟专用追肥(N∶P2O5∶K2O = 10∶0∶25)提供,在移栽后7—10d和30—40d均分两次穴施。在玉米、油菜及苕子种植季节,化学氮磷分别由尿素和过磷酸钙提供。其中,玉米和油菜70%的氮和全部磷肥作基肥,剩余30%的氮分别在油菜云苔期和玉米大喇叭口期作追肥穴施,苕子全施基肥。
1.3 土样采集与分析在不同轮作模式中,栽种或休闲是其中的重要环节,它们均可能改变土壤理化生物学性质,进而影响后季作物-烤烟生长发育和产量品质。因此,试验于2014年春季油菜和苕子旺长期,采集0—20cm的耕层土壤(含休闲处理),拣去杂物。部分土壤立即液氮冷冻备测微生物量碳氮和真菌18S rDNA序列,另取部分土壤风干,常规分析土壤pH、有机质和碱解氮、有效磷和有效钾[16]。微生物量碳氮采用氯仿熏蒸-0.5mol/L K2SO4提取,用K2Cr2O7氧化法测碳和腚酚蓝比色法测氮[17]。分别用3,5-二硝基水杨酸比色法、磷酸苯二钠比色法、腚酚蓝比色法、高锰酸钾滴定容量法和TTC比色法测定土壤蔗糖酶、磷酸酶、脲酶、过氧化氢酶和脱氢酶活性[18]。
提取土壤样品中的真菌18S rDNA,接着采用真菌18S区段中V5—V7区的通用引物0817F(5′-TTAGCATGGAATAATRRAATAGGA-3′)和1196R(5′-TCTGGACCTGGTGAGTTTCC-3′)进行扩增。再参照454高通量测序方法,纯化、定量和均一化真菌18S rDNA,送上海美吉生物科技有限公司利用Roche Genome Sequencer FLX测序平台进行测序[19]。然后,对有效序列进行去杂、修剪、去除嵌合体序列等过滤处理,得到优化序列,通过聚类分析形成分类单元(operational taxonomic units,OTUs),采用BLAST程序对比GenBank(http://ncbi.nlm.nih.gov)中的已知序列,根据97%的相似度确定18S rRNA基因序列对应的真菌属(种)名称。
于2014年烤烟收获季节,分叶位采收烟叶,三段式烘烤,按烟草行业标准分级定价,记录上、中、下等烟叶产量,均价和产值。
1.4 数据处理利用土壤真菌属(种)数(OTUs)和18S rDNA序列数(Reads)计算土壤真菌的种群特征值,包括多样性指数和优势度指数。
Shannon-Wiener多样性指数H的计算公式为:
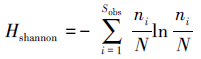
式中,Sobs为得到的OTUs的总数量;ni为真菌i的OTUs数量;N为得到的真菌的总数量。
Simpson优势度指数D的计算公式为:
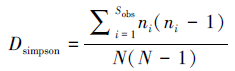
式中,Sobs为得到的OTUs的总数量;ni为真菌i的OTUs数量;N为得到的真菌的总数量[20, 21]。
真菌丰富度为某种真菌的18S rDNA读数占真菌18S rDNA总读数的百分数[22, 23]。
试验数据用Excel进行基本计算,SPSS16.0软件进行统计分析,差异显著性水平为P ≤ 0.05。
2 试验结果 2.1 烟叶产量、产值和上中等烟比例在T-V-R处理的土壤中,烟叶的产量、产值和上中等烟比例都为最高,烟叶产量和产值相比T-C-M处理分别增加了22.88%和25.30%;与T-C-R处理相比,上中等烟比例提高了3.62%(表 1)。
| 种植模式 Treatments | 产量/(kg/hm2) Yield | 产值/(元/hm2) Income | 上中等烟比例/% % of upper and middle grade of leaves |
| T-B-M:烤烟-休闲-玉米轮作 flue-cured tobacco-blank-maize crop rotation; T-C-M:烤烟-油菜-玉米轮作 flue-cured tobacco-canola-maize crop rotation; T-C-R:烤烟-油菜-水稻轮作 flue-cured tobacco-canola-rice crop rotation; T-V-R:烤烟-苕子-水稻轮作 flue-cured tobacco-vetch-rice crop rotation; 表中数据为平均数±标准差;在同一列中,有不同小写字母者表示差异显著,(P ≤ 0.05) | |||
| T-B-M | 192.4±11.5b | 5714.2±322.8b | 75.2±5.3b |
| T-C-M | 173.5±12.1c | 5291.7±350.4c | 75.3±6.8b |
| T-C-R | 185.7±16.7bc | 5403.8±476.4c | 74.5±8.2b |
| T-V-R | 213.2±8.5a | 6630.5±286.1a | 77.2±3.7a |
在不同轮作的土壤中,pH、有机质和有效养分含量都不同程度的增加或无显著变化(表 2)。与原始土壤相比,T-B-M和T-C-M处理使土壤pH值显著增加,最高为6.4,T-V-R和T-C-R处理对土壤的pH值无显著影响;T-C-M、T-C-R和T-V-R分别使有机质增加25.49%、16.23%和45.11%,T-B-M则无显著变化;T-V-R显著增加土壤碱解氮含量,为原始土壤的1.32倍;4种轮作处理均提高有效磷含量,为原始土壤的1.12—3.67倍;T-B-M的有效钾含量最高,其余3种轮作的有效钾含量与原始土壤相似。
| 种植模式 Rotation modals | pH | 有机质 OM/(g/kg土) | 有效养分Available nutrients/(mg/kg土) | ||
| N | P | K | |||
| 表 中数据为平均数±标准差;在同一列中,有不同小写字母者表示差异显著,(P≤ 0.05) | |||||
| 初始土壤Initial soil | 5.8±0.3bc | 11.77±0.4c | 57.2±3.4bc | 9.9±0.3e | 176.0±2.1b |
| T-B-M | 6.2±0.1a | 11.84±0.28c | 55.2±1.5c | 46.2±1.3a | 288.8±0.5a |
| T-C-M | 6.4±0.06a | 14.77±0.91b | 57.9±1.2bc | 39.5±0.08ab | 200.2±1.2b |
| T-C-R | 5.9±0.1b | 13.68±0.23b | 62.9±3.7b | 21.0±0.9cd | 195.4±1.7b |
| T-V-R | 5.6±0.02c | 17.08±0.69a | 75.8±1.8a | 29.9±0.4c | 199.4±0.6b |
不同轮作模式对土壤酶活的影响各不相同(表 3)。在T-V-R处理的土壤中,蔗糖酶、磷酸酶、脱氢酶、过氧化氢酶以及脲酶活性最高;在T-B-M、T-C-M和T-C-R处理的土壤中,蔗糖酶和脲酶活性分别变化于36.07—39.05和0.63—0.65之间,处理之间无显著差异;在T-C-M处理的土壤中,磷酸酶和脱氢酶活性低于其余3种轮作处理;T-B-M和T-C-R处理的土壤中,过氧化氢酶活性无显著差异,但显著低于T-C-M和T-V-R。
| 轮作处理 Rotation modals | 蔗糖酶 Sucrase/(g葡萄糖 g-1 h-1) | 磷酸酶 Phosphatase/ (mg pNP kg-1 h-1) | 脱氢酶 Dehydrogenase/(g TPF kg-1 h-1) | 过氧化氢酶 Catalase/(mL(0.1mol/L K2MnO4) h-1 g-1) | 脲酶 Urease/(mg NH+4-N kg-1 h-1) |
| 表 中数据为平均数±标准差;在同一列中,有不同小写字母者表示差异显著(P ≤ 0.05) | |||||
| T-B-M | 39.05±2.35b | 1.56±0.12b | 5.80±0.14b | 0.22±0.02b | 0.65±0.06b |
| T-C-M | 36.07±2.93b | 0.72±0.02c | 0.72±0.04c | 0.23±0.02a | 0.63±0.04c |
| T-C-R | 38.26±3.40b | 1.55±0.08b | 4.42±0.16b | 0.22±0.03b | 0.64±0.02bc |
| T-V-R | 73.41±4.06a | 2.21±0.11a | 10.84±1.04a | 0.23±0.03a | 0.68±0.05a |
图 1可见,在T-V-R处理的土壤中,微生物碳含量最高,为170.0mg/kg;T-C-M和T-C-R处理微生物碳含量次之但无显著差异,为117.2—119.4mg/kg;T-B-M最低,仅为61.9mg/kg。微生物氮含量的变化趋势则有所不同,T-V-R和T-C-M含量最高,达到8.9—9.4mg/kg,其次为T-B-M,T-C-R含量最低。实施不同轮作处理后,土壤微生物量碳氮比变化于7.5—19.1之间。

|
| 图 1 种植模式对土壤微生物碳氮含量的影响 Fig.1 Influence of rotation treatments on microbial C and N in soil T-B-M:烤烟-休闲-玉米轮作 flue-cured tobacco-blank-maize crop rotation; T-C-M:烤烟-油菜-玉米轮作 flue-cured tobacco-canola-maize crop rotation; T-C-R:烤烟-油菜-水稻轮作 flue-cured tobacco-canola-rice crop rotation; T-V-R:烤烟-苕子-水稻轮作 flue-cured tobacco-vetch-rice crop rotation |
随机抽取测序样品中的18S rDNA读数(reads),以真菌属(种)数(OTUs)为纵坐标,18S rDNA读数为横坐标,获得稀释曲线(图 2)[24]。结果表明,抽样读数大约在1500以下时,细菌种(属)类数迅速增加;读数在1500—4000之间,细菌种(属)数缓慢增加;读数超过4000之后,其种(属)类数的增长逐渐趋于平缓。但是,细菌稀释曲线因轮作处理不同而异,T-V-R最高,T-B-M次之,T-C-R和T-C-M最低,且几乎重叠。

|
| 图 2 不同轮作处理的土壤中,真菌稀释性曲线 Fig.2 Fungi rarefaction curves in soil under different rotation treatments |
在T-B-M、T-C-M、T-C-R和T-V-R处理的土壤中,454-高通量测序分别获得了13097,11345,12939和13763个18S rDNA 序列,分别代表 530、378、395和581种真菌(OTUs),归属于子囊菌门(Ascomycota)、壶菌门(Chytridiomycota)、担子菌门(Basidiomycota)、接合菌门(Zygomycota)和尚待鉴定的类型 (表 4)。其中,T-B-M、T-C-M和T-C-R 3个处理中待定真菌OTUs数量最多,占总OTUs数的46.03%—53.40%,其次是子囊菌门,比例为34.53%—37.22%;T-V-R处理中子囊菌OTUs数量最多,占总数的45.78%,其次是未知类型,占总数的39.76%。4个处理中,接合菌门、担子菌门和壶菌门合计约占总量的12.08%—15.61%。
| 真菌群落门组成 Phyla in fungal community | T-B-M | T-C-M | T-C-R | T-V-R |
| *表中数据为平均数;在同一行中,有不同小写字母者表示差异显著(P ≤ 0.05) | ||||
| 子囊菌门OTUs Ascomycota OTUs | 183b | 145bc | 147bc | 266a |
| 担子菌门OTUs Basidiomycota OTUs | 28b | 25b | 27b | 43a |
| 接合菌门OTUs Zygomycota OTUs | 31a | 8c | 24b | 34a |
| 壶菌门 OTUs Chytridiomycota OTUs | 5b | 26a | 10b | 7b |
| 未知类型 OTUs Unidentified OTUs | 283a | 174c | 187c | 231b |
| ∑OTUs | 530b | 378c | 395c | 581a |
在各轮作处理的土壤中,前15种优势真菌的丰富度合计占总量的29.46%—62.86%,尚待鉴定的真菌丰富度高达6.21%—45.75%。其中,在T-C-M和T-C-R处理中,待定菌株最多,高达9株,累计丰富度分别为45.75%和41.09%;其次是T-B-M处理,待定真菌有7株,丰富度为21.92%;在T-V-R处理中,待定菌株数量最少,仅有5株,丰富度为6.21%(表 5)。
| T-B-M | T-C-M | T-C-R | T-V-R | ||||
| 种类 Fungal Types | 丰富度/% Abundance | 种类 Fungal Types | 丰富度/% Abundance | 种类 Fungal Types | 丰富度/% Abundance | 种类 Fungal Types | 丰富度/% Abundance |
| 待定真菌-1 unclassified fungi-1 | 11.29 | 待定真菌-7 unclassified fungi-7 | 8.59 | 待定真菌-7 unclassified fungi-7 | 10.69 | 镰刀菌 Fusarium | 5.73 |
| 盘菌-1 Pezizomycetes-1 | 4.31 | 待定真菌-8 unclassified fungi-8 | 7.70 | 黑附球菌 Epicoccum_nigrum | 7.46 | 尖孢镰刀菌Fusarium_oxysporum | 3.41 |
| 盘菌-2 Pezizomycetes-2 | 3.35 | 待定真菌-9 unclassified fungi-9 | 7.15 | 待定真菌-10 unclassified fungi-10 | 7.23 | 肉座菌-1 Hypocreales-1 | 2.69 |
| 待定真菌-2 unclassified fungi-2 | 2.74 | 待定真菌-10 unclassified fungi-10 | 6.19 | 待定真菌-4 unclassified fungi-4 | 6.57 | 肉座菌-2 Hypocreales-2 | 2.19 |
| 待定真菌-3 unclassified fungi-3 | 2.24 | 待定真菌-11 unclassified fungi-11 | 4.37 | 枝孢菌 Cladosporium | 5.53 | 小孢绿盘菌 Cercophora_coprophila | 2.14 |
| 链格孢菌 Alternaria | 2.15 | 待定真菌-12 unclassified fungi-12 | 4.25 | 待定真菌-12 unclassified fungi-12 | 5.27 | 肉座菌-3 Hypocreales-3 | 1.82 |
| 尖孢镰刀菌 Fusarium_oxysporum | 2.11 | 接合菌-2 Zygomycota-2 | 3.60 | 格孢腔菌-1 Pleosporales-1 | 3.82 | 待定真菌-20 unclassified fungi-20 | 1.63 |
| 孢黑团壳菌 Massarinaceae | 2.03 | 待定真菌-13 unclassified fungi-13 | 2.50 | 待定真菌-16 unclassified fungi-16 | 3.35 | 炭角菌-1 Xylariales-1 | 1.55 |
| 待定真菌-4 unclassified fungi-4 | 1.81 | 真菌GD27a fungal_GD27a | 2.29 | 待定真菌-17 unclassified fungi-17 | 3.11 | 接合菌-1 Zygomycota-1 | 1.34 |
| 待定真菌-5 unclassified fungi-5 | 1.54 | 灰葡萄孢霉菌-1 Botryotinia_fuckeliana-1 | 2.22 | 待定真菌-18 unclassified fungi-18 | 1.90 | 待定真菌-4 unclassified fungi-4 | 1.34 |
| 接合菌-1 Zygomycota-1 | 1.44 | 待定真菌-14 unclassified fungi-14 | 1.68 | 柄孢壳菌Podospora_intestinacea | 1.89 | 枝孢菌 Cladosporium | 1.24 |
| 镰刀菌 Fusarium | 1.33 | 待定真菌-15 unclassified fungi-15 | 1.68 | 格孢腔菌-2 Pleosporales-2 | 1.58 | 待定真菌-21 unclassified fungi-21 | 1.20 |
| 待定子囊菌-1 Ascomycota-1 | 1.16 | 待定子囊菌-2 Ascomycota-2 | 1.64 | 待定真菌-14 unclassified fungi-14 | 1.55 | 链格孢菌 Alternaria | 1.13 |
| 待定真菌-6 unclassified fungi-6 | 1.14 | 不可培养的轮枝菌 uncultured_Verticillium | 1.61 | 芸苔油壶菌 Olpidium_brassicae | 1.50 | 待定真菌-22 unclassified fungi-22 | 1.02 |
| 嗜鱼外瓶霉菌 Exophiala_pisciphila | 1.05 | 灰葡萄孢霉菌-2 Botryotinia_fuckeliana-2 | 1.58 | 待定真菌-19 unclassified fungi-19 | 1.42 | 待定真菌-23 unclassified fungi-23 | 1.02 |
| 合计 Total | 39.69 | 57.06 | 62.86 | 29.46 | |||
| 待定真菌 | 21.92 | 45.75 | 41.09 | 6.21 |
在前15种优势菌株中,4种处理之间真菌属(种)的相似性极低,无共同存在的真菌属(种),且每种处理特有的真菌种类均超过9种(表 5)。此外,待定真菌-4(unclassified fungi-4)共同存在于T-B-M、T-C-R和T-V-R 3个处理中;枝孢菌(Cladosporium)同时存在于T-C-R和T-V-R处理中;T-B-M和T-V-R处理共有的真菌是尖孢镰刀菌(Fusarium_oxysporum)、接合菌-1(Zygomycota-1)、镰刀菌(Fusarium)和链格孢菌(Alternaria);T-C-M和T-C-R处理共有的真菌为待定真菌-7(unclassified fungi-7)、待定真菌-10(unclassified fungi-10)、待定真菌-12(unclassified fungi-12)、待定真菌-14(unclassified fungi-14)。
2.5.4 真菌群落特征值在T-V-R的土壤中,真菌多样性指数最高,高达5.34;T-B-M次之,为4.97;T-C-M和T-C-R最低,变化于4.10—4.28之间。但是,优势度指数T-C-R最高,达到0.04;T-V-R最低,仅0.01(表 6)。
| 种植模式 Rotation modals | 多样性指数 Diversity index | 优势度指数 Dominant index |
| 表 中数据为平均数;在同一列中,有不同小写字母者表示差异显著,(P ≤ 0.05) | ||
| T-B-M | 4.97b | 0.02c |
| T-C-M | 4.28c | 0.03b |
| T-C-R | 4.10c | 0.04a |
| T-V-R | 5.34a | 0.01d |
有机质是土壤的重要成分,显著影响土壤物理、化学、生物学性质。秸秆还田既可补充土壤有机质和作物需要的养分,又能改良土壤理化生物学性质[25]。在16年的长期轮作过程中,每年都有大量的秸秆还田,土壤有机质和有效养分含量提高或无显著变化,说明在高强度轮作种植条件下,辅以秸秆还田或冬季休闲均可保持或提高土壤生产力。此外,各轮作处理的有效磷含量远远超过原始土壤,故适量减施磷肥很有必要。土壤pH值变化于5.6—6.4,都处于适合多种作物生长的pH值范围内[26]。综合作物产量、产值、土壤pH、有机质和有效养分的状况,以T-V-R种植模式最佳。
土壤有机质是微生物的碳源和氮源[27]。在轮作处理的土壤中,每年都实施秸秆还田,各种作物秸秆腐解所释放的有机质组成具有多样性,可满足多种微生物,尤其是异养型微生物-真菌对碳源和养分的需要。在实施T-V-R轮作的处理中,水旱轮作,嫌/好气交替,创造了适合多种微生物繁衍的不同土壤环境;加之烤烟、水稻和苕子的近缘性小,有机成分差异较大,可满足不同微生物的碳源和营养需要,有益于它们的繁殖生长。此外,油菜为直根系,而苕子为须根系,后者根系发达,能分泌大量的单(多)糖、氨基酸和有机酸等碳水化合物,向根际微生物提供碳源,促进其生长繁殖[28]。因此,在T-V-R土壤中,微生物量碳氮最高,18S rNDA读数最大,真菌属(种)类最多,土壤酶活性最强。脱氢酶、脲酶和磷酸酶活性与土壤微生物量呈显著正相关,推测与土壤微生物的分泌作用有关,类似前人研究结果[29, 30]。
在不同轮作处理的土壤中,微生物碳氮比相差2.5倍以上,意味着土壤微生物的组成结构各不相同。本文采用454高通量测序方法,揭示了真菌从门类到属(种)的组成、优势菌株、丰富度和多样性特征等。结果表明,稀释性曲线呈迅速增加-缓慢增加-逐渐平缓的变化趋势,说明18S rDNA的样本量和测序数据合理,准确地反映了样本中的真菌数及组成情况;稀释曲线的峰值高低不一,意味着轮作处理影响了土壤真菌的多样性。一般认为,多样性指数表示生物群落中的物种多寡,数值愈大表示群落中的物种越丰富;优势度指数越大,生物群落内的奇异度越高,优势种群突出[31]。生物种群丰富和多样性指数高是生态环境健康稳定的重要表现[32]。在健康稳定的生态环境中,生物多样性指数较高,优势度指数较低。T-V-R显著提高真菌群落的多样性指数,但降低优势度指数。因此,T-V-R处理的土壤生态环境较好,使之适合多种真菌的繁殖生长,种群数量增加。多种真菌共同存在,互相制约,可防止某些病原真菌过度繁殖,抑制作物病害的发生。值得注意的是,454高通量测序表明,在不同种植模式的土壤中,真菌的丰富度因轮作处理和它们的种类不同而异,存在378—581种(属)的真菌,远远超过了目前的常规培养、磷脂脂肪酸和PCR-DGGE能达到的水平。此外,用形态和生理生化方法鉴定微生物需要丰富的经验,程序复杂,耗时长,容易出现误判。说明454高通量测序检测灵敏度高,鉴定准确性可靠,更能深入研究土壤真菌的组成和群落结构。本项研究表明,18S rDNA读数和真菌OTUs以T-V-R最高,T-C-M最低,说明T-V-R土壤中的真菌数和属(种)类最多。
除了待定真菌外,子囊菌占各处理真菌的绝大多数,都为40%左右甚至超过40%以上。经16a不同轮作后,4种轮作处理的土壤中,前15种优势菌株的相似性极低,说明长期轮作极大地改变了土壤中真菌的组成及类型。需要指出的是,土壤真菌参与土壤有机质矿化,分解进入土壤的作物残体和有机肥中的有机质,供给植物营养[33]。但是,大多数真菌同时也为植物病原菌,引起多种真菌病害。例如,子囊菌引起根腐、茎腐、果(穗)腐和枝枯等;镰刀菌普遍为植物真菌性病害;葡萄肉坐菌几乎能危害所有果树,如香蕉蕉腐病、桃树流胶病等;座囊菌是我国主要的果树病菌,引起香蕉叶斑病,梨叶斑病,葡萄黑痘病等;链格孢菌95%以上兼性寄生于植物,引起多种植物病害,如小麦叶枯病,玉米大斑病,茄子早疫病,白菜黑斑病等;炭角菌广泛分布于全球各地,在热带和亚热带地区尤为常见,能引起桑、茶、栎、苹果根腐病和黑腐病等[34]。从防治作物土传真菌病害的角度看,合理轮作是完全可能和必要的。
4 结论总之,4种轮作方式均有益于保持或提高土壤肥力和土地生产力。从作物产量、产值、土壤有机质、有效养分、酶活性、微生物量、真菌种群结构和多样性等方面看,烤烟-苕子-水稻轮作最佳,是值得推广的一种种植模式。
| [1] | 王飞, 李世贵, 徐凤花, 顾金刚. 连作障碍发生机制研究进展. 中国土壤与肥料, 2013, (5): 6-13. |
| [2] | 张北赢, 陈天林, 王兵. 长期施用化肥对土壤质量的影响. 中国农学通报, 2010, 26(11): 182-187. |
| [3] | 刘高远, 郭天文, 谭雪莲, 马得祯, 张国宏, 陈光荣. 不同栽培方式下马铃薯土壤微生物区系的动态变化. 华中农业大学学报, 2014, 33(4): 19-24. |
| [4] | 邢会琴, 肖占文, 闫吉智, 马建仓, 孟嫣. 玉米连作对土壤微生物和土壤主要养分的影响. 草业科学, 2011, 28(10): 1777-1780. |
| [5] | 晋艳, 杨宇虹, 段玉琪, 孔光辉. 烤烟轮作、连作对烟叶产量质量的影响. 西南农业学报, 2004, 17(增刊): 267-271. |
| [6] | Zhang W, Long X Q, Huo X D,Chen Y F, Lou K. 16S rRNA-based PCR-DGGE analysis of actinomycete communities in fields with continuous cotton cropping in Xinjiang, China. Soil Microbiology, 2013, 66(2): 385-393. |
| [7] | 邹雨坤, 张静妮, 杨殿林, 陈秀蓉, 张天瑞, 赵建宁, 赵帅. 不同利用方式下羊草草原土壤生态系统微生物群落结构的PLFA分析. 草业学报, 2011, 20(4): 27-33. |
| [8] | 王先华. 稻田不同轮作方式对培肥地力的作用. 耕作与栽培, 2002, (6): 9-10. |
| [9] | 于高波, 吴凤芝, 周新刚. 小麦、毛苕子与黄瓜轮作对土壤微生态环境及产量的影响. 土壤学报, 2011, 48(1): 175-184. |
| [10] | Kennedy A C, Smith K L. Soil microbial diversity and the sustainability of agricultural soils. Plant and Soil, 1995, 170(1): 75-86. |
| [11] | 张鼎华, 叶章发, 李宝福. 杉木、马尾松轮作对林地土壤肥力和林木生长的影响. 林业科学, 2011, 37(5): 10-15. |
| [12] | 袁龙刚, 张军林, 张朝阳, 别国勇. 连作对辣椒根际土壤微生物区系影响的初步研究. 陕西农业科学, 2006, (2): 49-50. |
| [13] | 段曌, 肖炜, 王永霞, 赖泳红, 崔晓龙. 454测序技术在微生物生态学研究中的应用. 微生物学杂志, 2011, 31(5): 76-81. |
| [14] | 李天福, 王树会, 王彪, 邵岩. 云南烤烟轮作现状与对策分析. 烟草农业科学, 2006, 2(2): 216-222. |
| [15] | 郁飞燕, 李友军. 轮作对土壤肥力的影响. 科技传播, 2011, (16): 60-60. |
| [16] | 杨剑虹, 王成林, 代亨林. 土壤农化分析与环境监测. 北京: 中国大地出版社, 2008: 26-75. |
| [17] | Frostegård Å, Tunlid A, Bååth E. Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Applied and Environmental Microbiology, 1993, 59(11): 3605-3617. |
| [18] | 关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986: 274-339. |
| [19] | Herlemann D P R, Lundin D, Labrenz M, Jürgens K, Zheng A L, Aspeborg H, Andersson A F. Metagenomic de novo assembly of an aquatic representative of the verrucomicrobial class spartobacteria. MBIO, 2013, 4(3): e00569-12. |
| [20] | Krebs C J. Ecological Methodology. New York:Benjamin Cummings, 1989: 654-654. |
| [21] | Schloss P D, Gevers D, Westcott S L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rDNA-based studies. PLoS ONE, 2011, 6(12): e27310. |
| [22] | Roesch L F W, Fulthorpe R R, Riva A, Casella G, Hadwin A K M, Kent A D, Daroub S H, Camargo F A O, Farmerie W G, Triplett E W. Pyrosequencing enumerates and contrasts soil microbial diversity. The ISME Journal, 2007, 1(4): 283-290. |
| [23] | Lim Y M, Kim B K, Kim C, Jung H S, Kim B S, Lee J H, Chun J. Assessment of soil fungal communities using pyrosequencing. The Journal of Microbiology, 2010, 48(3): 284-289. |
| [24] | Amato K R, Yeoman C J, Kent A, Righini N, Carbonero F, Estrada A, Gaskins H R, Stumpf R M, Yildirim S, Torralba M, Gillis M, Wilson B A, Nelson K E, White B A, Leigh S R. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. The ISME Journal, 2013, 7: 1344-1353. |
| [25] | 徐国伟, 李帅, 赵永芳, 陈明灿, 李友军. 秸秆还田与施氮对水稻根系分泌物及氮素利用的影响研究. 草业学报, 2014, 23(2): 140-146. |
| [26] | 李念胜, 王树声. 土壤pH值与烤烟质量. 中国烟草, 1986, (2): 12-14. |
| [27] | Katarina H. Soil microbial community structure in relation to vegetation management on former agricultural land. Soil Biology & Biochemistry, 2002, 34(9): 1299-1307. |
| [28] | 焦彬. 中国绿肥. 北京: 农业出版社, 1986: 97-110. |
| [29] | 袁亮. 设施栽培土壤微生物量和酶活性的变化规律及其与土壤肥力的关系[D]. 泰安: 山东农业大学, 2007. |
| [30] | 单贵莲, 初晓辉, 罗富成, 马玉宝, 李临杭, 陈功. 围封年限对典型草原土壤微生物及酶活性的影响. 草原与草坪, 2012, 32(1): 1-6. |
| [31] | Hunter P R, Gaston M A. Numerical index of the discriminatory ability of typing systems: An application of Simpson's index of diversity. Journal of Clinical Microbiology, 1988, 26(11): 2465-2466. |
| [32] | 高东, 何霞红. 生物多样性与生态系统稳定性研究进展. 生态学杂志, 2010, 29(12): 2507-2513. |
| [33] | 徐严. 若尔盖高原湿地土壤真菌的初步研究[D]. 成都: 四川农业大学, 2008. |
| [34] | 康振生. 我国植物真菌病害的研究现状及发展策略. 植物保护, 2010, 36(3): 9-12. |