 2016, Vol. 36
2016, Vol. 36文章信息
- 吴鹏, 王襄平, 张新平, 朱彪, 周海城, 方精云
- WU Peng, WANG Xiangping, ZHANG Xinping, ZHU Biao, ZHOU Haicheng, FANG Jingyun
- 东北地区森林凋落叶分解速率与气候、林型、林分光照的关系
- Effects of climate, forest type and light availability on litter decomposition rate in forests of Northeast China
- 生态学报, 2016, 36(8): 2223-2232
- Acta Ecologica Sinica, 2016, 36(8): 2223-2232
- http://dx.doi.org/10.5846/stxb201410101991
-
文章历史
- 收稿日期: 2014-10-10
- 网络出版日期: 2015-05-20






2. 北京大学生态学系, 北京大学地表过程与分析模拟教育部重点实验室, 北京 100871;
3. 吉林省长白山自然保护管理中心, 安图 133613
2. Department of Ecology and Key Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China;
3. Center for Natural protection and management of the Mt. Changbai reserve, Antu 133613, China
凋落物分解是森林碳循环和全球碳平衡的一个重要环节[1, 2, 3]。由凋落物分解返回大气中的碳含量是全球碳平衡的重要组成部分[4]。据估计,凋落物(包括根凋落物)分解产生的碳大约占到全球陆地生态系统年际碳流通总量的70%,达到68 Pg C/a(Pg=1015g)[5]。
我国东北地区森林覆盖率高,碳储量大,仅黑龙江、吉林、辽宁三省的森林碳储量就占全国森林总碳储量的24%—31%[6],是东亚地区的一个重要碳汇[7]。而高纬度、寒冷地区凋落物分解由于受热量制约,对气候变暖更为敏感[3, 8]。据估计,到21世纪后期,中国东北地区气温将可能较目前变暖3.0℃或以上,降水将可能增加[9]。在气候变暖的背景下,研究我国东北地区凋落物分解与气候的关系具有重要意义。
研究表明,凋落物分解速率受多种因素的作用,包括气候和局域环境条件,凋落物质量,植被类型及土壤生物等[10, 11, 12, 13]。关于东北森林凋落物分解已经开展了不少研究,如王凤友等对凉水地区红松和几种阔叶树凋落叶的分解和养分释放的研究[14],沈海龙对帽儿山樟子松人工林针阔叶凋落物分解动态的研究[15],张东来对帽儿山6种主要树种凋落物分解速率的研究[16],以及郭忠玲等对长白山主要树种和林型凋落物分解的比较研究等[17]。这些研究虽然揭示了某一地点不同林型或树种的分解速率差异,但地理跨度小,未能探讨大尺度上凋落物分解的影响因素。在整个东北地区的尺度上,宏观气候因素、森林类型、局域环境条件如何共同作用影响凋落叶的分解?这些因素的相对重要性如何?相关问题还需要定量研究,以深入理解森林碳循环过程,并可为生物地球化学模型提供重要参数。
大尺度的研究表明气候和植被类型对凋落物分解有重要影响[13, 18, 19]。凋落物分解速率在植被或森林类型之间的差异,一方面是因为不同的植被分布在不同的气候条件下,气候的差异对分解速率有着直接的影响[20];另一方面是由于气候通过其它因素的间接作用。比如,不同地区的植被物种组成不同、因而植物功能性状不同[21, 22, 23],后者直接影响凋落物质量,是凋落物分解的重要影响因素[13, 24]。前述东北地区的研究也表明,相同气候下的不同林型由于树种组成不同,分解速率有显著差异。然而,这两种因素对凋落物分解的相对影响大小,却很少有研究涉及。气候变化显然会导致物种组成的变化、甚至森林类型的变化[25, 26],如果后一种机制对凋落物分解的影响占主导作用,则意味着不考虑物种组成变化的因素,是无法准确预测气候变化对森林碳循环的影响的。
此外,小尺度的研究常发现林内光照对凋落物分解的重要影响。如林冠空隙可通过改变林内微环境影响土壤生物活动,进而加速或阻碍凋落物分解[27, 28],对于针叶林其作用更是不可低估[29]。在人为干扰不大的情况下,林内光照是和气候条件导致的林型、物种组成差异有关的。如大兴安岭的明亮针叶林的林内光照条件要好于东北东部山地的针阔混交林,有研究表明这种差异对林下植物多样性是有明显影响的[30]。因此光照条件也有可能和气候、森林类型等因素协同作用于凋落物的分解。但迄今为止,大尺度上的凋落物分解研究尚未考虑过这一因素。
本文对东北主要山地(大小兴安岭、张广才岭、长白山)主要森林类型的凋落叶分解进行了系统研究。目的在于:(1)研究气候对东北地区主要森林类型的凋落叶分解速率的影响;(2)探讨森林类型、林冠透光率如何与气候因子共同作用,影响凋落叶的分解速率。
1 研究地点及方法 1.1 研究地点及样地设置研究选取东北林区的4个地点,分别是长白山(14块样地),张广才岭的帽儿山(6块样地),小兴安岭的凉水(4块样地)和大兴安岭的根河(2块样地)共设置26块固定观测样地,样地面积为1 000 m2 (20 m×50 m)。各研究地点的概况见张新平等[31]。长白山的样地沿海拔梯度设置(海拔从650m至1940m),包含了各森林垂直带。其余海拔梯度不大的地点则选择当地典型的森林类型设立样地。样地设置时选择没有明显干扰痕迹的郁闭林分。本研究的样地包括了东北地区5种主要森林类型,在纬度上跨越了温带和寒温带,在海拔梯度上包括了从低山带到亚高山带的各种群落类型,具有较好的代表性。林型包括落叶阔叶林、针阔叶混交林、落叶针叶林、常绿针叶林、岳桦林。岳桦林本属于落叶阔叶林,但由于其生长在高山林线附近,与低海拔的落叶阔叶林不论在气候条件、物种组成、群落结构等方面均有很大的不同(表 1),因此在分析时将其单独作为一种林型予以考虑。
| 林型 Forest type | 温暖指数 WI/ (℃·month) | 降水 MAP/mm | 主要树种组成 Major species composition | 样地数 No. plots |
| DB: Deciduous broadleaf forest; DNM: Deciduous broadleaf and needle-leaf mixed forest; DN: Deciduous needle-leaf forest; EN: Evergreen needle-leaf forest; BE: Betula ermanii forest,WI,warmth index; MAP,mean annual precipitation. | ||||
| 落叶阔叶林 DB | 45.0—57.8 | 607—838 | 白桦 Betula platyphylla (30.0%),蒙古栎 Quercus mongolica (16.8%),青杨 Populus cathayana (16.7%),色木槭 Acer mono (8.6%) | 5 |
| 针阔叶混交林 DNM | 44.4—53.6 | 626—844 | 红松 Pinus koraiensis (53.7%),紫椴 Tilia amurensis (11.9%), 蒙古栎 Quercus mongolica (9.9%),大青杨 Populus ussuriensis (8.8%) | 6 |
| 落叶针叶林 DN | 29.3—59.1 | 470—981 | 兴安落叶松 Larix gmelinii (67.5%),长白落叶松 Larix olgensis (28.3%) | 7 |
| 常绿针叶林 EN | 24.6—43.3 | 856—1049 | 长白落叶松 Larix olgensis (36.2%),鱼鳞云杉 Picea jezoensis (21.4%),臭冷杉 Abies nephrolepis (19.0%),红松 Pinus koraiensis (10.8%),红皮云杉 Picea koraiensis (5.2%) | 5 |
| 岳桦林 BE | 17.9—19.9 | 1105—1131 | 岳桦 Betula ermanii (85.1%),长白落叶松 Larix olgensis (14.5%) | 3 |
用网袋法进行分解实验,网袋大小为15 cm×20 cm,网孔为1 mm×1 mm[32]。于2003年9月底在4个地点的每块样地内收集新鲜凋落叶、自然风干。每个样地取风干叶样300 g于65 ℃烘至恒重,测定含水率。每个分解袋内装入30 g风干叶样品放回原样地的土壤表层。每个样地放置60袋,于每年4—11月的月初取2袋。研究持续了3a,到2006年11月为止。分解袋取回后去除杂质、土壤,在65℃下烘至恒重后称重。
凋落叶分解过程采用修正的[33]指数衰减函数进行拟合[34, 35]:

林冠空隙会影响到光照、热量、水分等在林冠层下的再分配,是影响林内微环境的重要因素,因此进行了林冠透光状况的测定,以研究它对凋落物分解速率的可能影响。在每块样地内均匀布设5个点,通过WinScanopy林冠分析系统拍摄数码照片。拍摄时间选在天气晴朗的8:00—10:00和14:00—16:00,以消除太阳直射光斑的影响。对拍摄的180°角的林冠层数码照片,应用WinScanopy For Canopy Analysis软件进行分析(加拿大Regent Instruments公司),得到林冠空隙度、林冠开度等光照参数,以反映林冠层的透光率。本研究使用的指标为林冠空隙度(取5个点的平均值),林冠开度与其生物学意义、解释力接近,因而在最终数据分析中没有使用。
1.4 气候指标的计算采用如下方法推算各样地的气温、降水等气候指标:
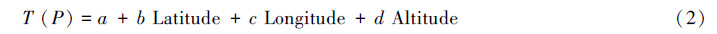
利用上述方法得到所有样地的1—12月月平均温度和降水,并计算如下气候指标:
1)温暖指数(WI) WI=∑(ti-5),其中ti为5℃以上的月均温。
2)年降水量(MAP) 一年各月降水量之和。
1.5 数据分析采用回归分析和方差分析探讨三类因素对分解速率的影响。1)气候因子:包括温暖指数、年降水量,分别反映热量和水分的有效性[38];2)林分光照状况,以林冠空隙度反映林冠层透光率;3)林型,即落叶阔叶林、针阔叶混交林、落叶针叶林、常绿针叶林、岳桦林等5种林型。数据分析之前均先进行正态性检验和方差齐性检验。各林型之间凋落叶分解参数的比较采用多重比较Duncan检验。
为了分析气候、林冠透光率和林型之间的协同作用和独立作用,采用变异分离的方法,将总变异分解为如下组分:(1)3种因素各自的独立作用(a,b,c);(2)两因素之间的协同作用(d,e,f);(3)3种因素之间的协同作用(g);(4)未解释的变异。各组分的计算方法见Heikkinen等[39]。采用Wang等提出的随机化检验方法进行各组分显著性的检验[40]。
数据分析使用R 3.1.1 (R Development Core Team 2014)软件进行。
2 结果与讨论 2.1 各林型的凋落叶分解速率比较不同林型凋落叶分解参数可以看出(图 1),各林型凋落叶分解常数k的平均值依次为:落叶阔叶林(0.338)、针阔叶混交林(0.293)、落叶针叶林(0.207)、常绿针叶林(0.146)、岳桦林(0.097),并且落叶阔叶林、针阔叶混交林与常绿针叶林、岳桦林具有显著性差异。这与长白山其他研究中[17, 41]得出的相同(似)林型的k值相比偏小,可能主要是由于这些研究的时间跨度一般为1—2a,而本研究为3a。凋落叶分解速率前期较大,后期则逐渐递减,因此研究时间较短时得到的k值会偏大。不少研究都指出过长时间凋落物分解实验的重要性[42, 43]。

|
| 图 1 不同林型凋落叶分解速率的差异 Fig.1 Comparison of litter decomposition rate between forest types decomposition coefficient (k) 上标字母有相同者在P<0.05水平上无显著差异; BE、DB、DN、DNM、EN,森林林型,分别为岳桦林(Betula ermanii forest,)、落叶阔叶林(Deciduous broadleaf forest)、落叶针叶林(Deciduous needle-leaf forest)、针阔叶混交林(Deciduous broadleaf and needle-leaf mixed forest)、常绿针叶林(Evergreen needle-leaf forest) |
对于分解95%所需时间,落叶阔叶林、针阔叶混交林的平均值(分别为9.2、9.9a)均显著小于常绿针叶林、岳桦林(分别为19.7、24.6a),落叶针叶林介于中间(14.5a)。
2.2 凋落叶分解与各影响因素的关系 2.2.1 分解参数与海拔、纬度的关系为考察分解速率与海拔的关系,选取长白山的样地进行分析(表 2,图 2)。结果表明,分解常数k、分解95%所需时间与海拔之间的回归关系均极为显著(P<0.001)。随海拔的升高,k值下降,分解95%所需时间延长。由于各研究地点样地的海拔差异较大(长白山样地在650—1940 m之间,其余4个地点的样地在300—840 m之间),没有单独分析分解参数和纬度的相关性。不过,对分解参数与纬度、海拔的多元回归分析表明,在回归模型中纬度的作用并不显著(k: P=0.92; t95%: P=0.83),而海拔则在P=0.001水平显著。这一关于纬度的统计结果可能与本研究的不同地点海拔差异较大、纬度梯度还不够大有关,在全国和全球大尺度上的研究都发现,分解速率随纬度上升显著下降[13, 19]。
| 林型 Forest type | 海拔 Altitude /m | 样地数 No. plots |
| 落叶阔叶林 DB | 950 | 1 |
| 针阔叶混交林 DNM | 650—970 | 3 |
| 落叶针叶林 DN | 1420—1440 | 2 |
| 常绿针叶林 EN | 1010—1660 | 5 |
| 岳桦林 BE | 1850—1940 | 3 |

|
| 图 2 分解常数k、分解95%所需时间(t95%)随海拔的变化 Fig.2 Changes of leaf litter decomposition coefficient (k) and the time for 95% litter decomposed (t95%) with altitude |
海拔、纬度等地理因子与分解速率的关系,实质是通过对热量、水分的分配而起作用。对气候因子WI、MAP与分解参数关系的分析表明(图 3),各相关关系均极为显著,分解常数k与热量(WI)呈正相关,与水分(MAP)呈负相关;凋落叶分解95%所需时间则反之。东北森林区降水较充沛,热量是生产力的主要限制因子,而水分不构成限制[30, 31]。因此分解速率与降水的负相关关系可能主要是由于MAP和WI的负相关关系所导致(R2=0.51; P<0.001)。一般认为,在热带生态系统,降水量增加对凋落物分解有明显的促进作用,但在一些温带生态系统中,较高的降水量反而可能使凋落物分解减慢[44, 45]。一方面,过多的雨水导致土壤嫌气条件,不利于土壤生物的活动;另一方面,降水可能改变凋落物基质质量,比如降低磷浓度、增加木质素浓度等[46],从而对分解产生负效应。本文中分解速率与降水的负相关也可能与这种因素有关,但这种因素的贡献有多大还需进一步研究。

|
| 图 3 分解常数k、分解95%所需时间(t95%)与气候因子温暖指数(WI)和年降水量(MAP)的关系 Fig.3 Relationship of decomposition coefficient k and the time for 95% litter decomposed (t95%) with climate indices WI,warmth index; MAP,Mean annual precipitation |
对林冠空隙度与分解参数关系的分析表明(图 4),林冠空隙度与分解常数k呈显著负相关,与分解95%所需时间相反。即林冠空隙越大,凋落叶分解速率减小。Zhang和Zak的研究发现林隙较小处的凋落物比林隙大的分解得更快,并且认为较大的林冠空隙会通过改变微环境条件而显著减小微生物活性和分解速率[47]。一些其他研究也得出了类似的结果[28, 48]。图 4与这些研究结果是一致的。

|
| 图 4 分解常数k、分解95%所需时间(t95%)与林冠空隙度的关系 Fig.4 The relationship between decomposition coefficient (k) and the time for 95% litter decomposed (t95%) with gap fraction |
前述结果表明,气候、林冠透光率、林型等因素对凋落叶分解都有显著的影响。为了研究这些因素如何共同作用于分解速率,采用变异分离来反映各因素的协同作用和独立作用。图 5表明,气候因子(WI和MAP)共计解释了k值的55.5%的变异(a+d+e+g),而林型的总解释力为66.0% (b+d+f+g),林冠空隙度的总解释力为24.2% (c+e+f+g)。分解95%所需时间的结果与k值相似。这一结果与以往的研究一致,即气候和林型是凋落叶分解的重要影响因素[13, 19]。

|
| 图 5 气候、林型、林冠空隙度对分解参数影响的变异分解图 Fig.5 Variance partitioning for the effects of climate,forest type and gap fraction on decomposition constant (k) and the time for 95% litter decomposed(t95%),in terms of the percentage of sum of squares explained 图中给出了各组分占总变异的百分比; a,b,c表示各因素之间的独立解释力; d,e,f,g表示各因素之间的协同解释力 |
不过,图 5进一步揭示,气候、林型、林冠透光率对分解速率的解释力主要属于三种因素之间的协同作用,各因素的独立解释力(a、b、c)并不大。随机化检验的结果表明,在各方差组分中,仅d和g的作用是显著的。其中气候和林型的协同解释力(d+g)高达46.8% (k) —56.8% (t95%)。这说明气候对凋落叶分解的作用,在很大程度上是由于气候变化导致林型(物种组成)的变化、进而影响分解速率(见引言)。至少在本研究中,气候对分解速率的直接影响并不大(<9%)。
虽然在很多小尺度研究中已经证实林内光照、林冠空隙对凋落物分解有重要的影响[28, 48],但由于大尺度的凋落物分解研究没有测定过林分光照状况,宏观气候和林内光照对分解速率的相对作用大小还没有进行过研究。事实上,一些研究早就指出过,气候条件的变化会显著影响林内的光环境[49]:在高纬度地区的针叶林,树冠呈锥形、树木间间距较大、林冠垂直层次简单;而在南方的阔叶林中,树冠趋向扁平、林冠垂直层次复杂且林分密度较大。这种气候条件导致的林冠结构差异,会显著影响林内光照,并对森林更新、多样性产生重要影响[30, 49]。因此,在人为干扰不大的情况下(如本文的郁闭林分样地),林内光照条件应当是随气候条件而变化的。本研究的测定结果也证实了这一点:随着热量(WI)的增加,林冠透光率显著下降(图 6)。在东北地区,从北到南随着热量的增加,森林类型由垂直结构简单、透光率高的兴安落叶松林逐渐过渡到垂直层次复杂的针阔混交林[30, 50]。图 6的结果与这一事实是一致的。

|
| 图 6 林冠透光率和温暖指数的关系 Fig.6 The relationship between gap fraction and warmth index (WI) |
变异分离的结果表明(图 5),气候、林型和林冠透光率的协同作用(g)是最重要的变异组分之一(而且作用显著),分别解释了k和t95%变异的23.9%和22.3%。因此,气候通过林分光照等生物因素对凋落物分解的间接作用是不可忽视的,即热量的增加除了对分解速率的直接影响外,还可通过森林类型(物种组成和林冠结构)的改变导致的林冠透光率下降,从而间接地促进凋落叶的分解(图 4)[48, 51]。
3 结论与讨论本研究结果表明,各林型凋落物分解速率大小依次为:落叶阔叶林、针阔叶混交林、落叶针叶林、常绿针叶林、岳桦林。从总体上看,阔叶>针叶,落叶>常绿,这和大多数研究得出的结论相同[52]。一方面,相比于针叶凋落物,阔叶凋落物具有更高的养分含量和更低的木质素、多酚含量,因此可能分解得更快[53]。另一方面,阔叶林或混交林凋落物形成的枯枝落叶层相比于针叶林的更有利于凋落物分解。因为一般认为阔叶形成的细腐殖质枯枝落叶层(mull forest floor)所含营养元素更为丰富,大型土壤动物物种丰富度高,可加速凋落物的机械粉碎,刺激微生物活性,从而加速分解进程[54]。
凋落物分解是多种因素共同作用的结果,并且不同因素在凋落物分解的不同阶段所起的相对作用大小是不同的。因此,弄清楚这些因素是如何相互影响并且共同作用于凋落物分解过程就尤其重要。
气候、林型、林冠透光率对分解速率的影响结果表明,三者之间的协同作用对于凋落物分解起着控制性作用。气候中温度、水分在不同地区对于凋落物分解起着明显不同的作用。在干旱或半干旱地区,随海拔升高,分解速率反而加快,这是由于在这些地区海拔较高的地方,尽管温度降低,但湿度也会增大,总的作用还是加快分解速率[55]。而对于水分不构成限制性因素的东北地区,分解速率则随海拔的升高而减小。本研究结果表明,气候对分解速率的影响在很大程度上是通过如下机制实现的:1) 气候的变化导致林型的变化,即通过影响物种组成导致凋落叶质量的变化[24],进而影响分解速率;2) 气候和林型的变化还导致林冠透光率的变化,间接作用于分解速率。林型对分解速率的解释力为66.0%,可见不同林型间凋落物质量的差异对分解速率具有较强的解释力,这可以通过不同树种所形成的独特周围环境条件的影响来起作用,并且植物物种特性和植物-土壤的长期互相作用深刻地影响着凋落物的分解[56]。林冠透光率相比于气候、林型的解释力较小,但由于其受气候、林型双重影响,对于凋落物的分解作用依然不可忽视,不少研究发现林冠空隙大小对于凋落物分解具有重要作用[28, 51]。因此,需要深入研究气候和生物因素(如物种组成、林冠结构)如何交互作用影响凋落物分解,才能更好地理解气候变化对森林碳循环的影响。
| [1] | Aerts R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos, 1997, 79(3): 439-449. |
| [2] | Couteaux M M, Bottner P, Berg B. Litter decomposition, climate and liter quality. Trends in Ecology & Evolution, 1995, 10(2): 63-66. |
| [3] | Robinson C H. Controls on decomposition and soil nitrogen availability at high latitudes. Plant and Soil, 2002, 242(1): 65-81. |
| [4] | Shaver G R, Billings W D, Chapin F S, Giblin A E, Nadelhoffer K J, Oechel W C, Rastetter E B. Global change and the carbon balance of arctic ecosystems. BioScience, 1992, 42(6): 433-441. |
| [5] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B, 1992, 44(2): 81-99. |
| [6] | Fang J Y, Chen A P, Peng C H, Zhao S Q, Ci L J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 2001, 292(5525): 2320-2322. |
| [7] | Tan K, Piao S L, Peng C H, Fang J Y. Satellite-based estimation of biomass carbon stocks for northeast China's forests between 1982 and 1999. Forest Ecology and Management, 2007, 240(1/3): 114-121. |
| [8] | Hobbie S E, Nadelhoffer K J, Högberg P. A synthesis: the role of nutrients as constraints on carbon balances in boreal and arctic regions. Plant and Soil, 2002, 242(1): 163-170. |
| [9] | 赵宗慈, 罗勇. 21世纪中国东北地区气候变化预估. 气象与环境学报, 2007, 23(3): 1-4. |
| [10] | Aber J D, Melillo J M, Nadelhoffer K J, Pastor J, Boone R D. Factors controlling nitrogen cycling and nitrogen saturation in northern temperate forest ecosystems. Ecological Applications, 1991, 1(3): 303-315. |
| [11] | Wardle D, Lavelle P. Linkages between soil biota, plant litter quality and decomposition//Cadisch G, Giller K E, eds. Driven by Nature: Plant Litter Quality and Decomposition. Wallingford: CAB International, 1997: 107-124. |
| [12] | Austin A T, Vivanco L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature, 2006, 442(7102): 555-558. |
| [13] | Zhang D Q, Hui D F, Luo Y Q, Zhou G Y. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. Journal of Plant Ecology, 2008, 1(2): 85-93. |
| [14] | 王凤友, 王业蘧. 红松和五种阔叶树枯叶的分解及养分释放//周晓峰. 森林生态系统定位研究.第一集. 哈尔滨: 东北林业大学出版社, 1991: 250-259. |
| [15] | 沈海龙, 丁宝永, 沈国舫, 陈爱民. 樟子松人工林下针阔叶凋落物分解动态. 林业科学, 1996, 32(5): 393-402. |
| [16] | 张东来, 毛子军, 朱胜英, 周彪. 黑龙江省帽儿山林区6种主要林分类型凋落物研究. 植物研究, 2008, 28(1): 104-108. |
| [17] | 郭忠玲, 郑金萍, 马元丹, 李庆康, 于贵瑞, 韩士杰, 范春楠, 刘万德. 长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究. 生态学报, 2006, 26(4): 1037-1046. |
| [18] | 王健健, 王永吉, 来利明, 赵学春, 王飞, 申国珍, 赖江山, 鲁洪斌, 赵春强, 郑元润. 我国中东部不同气候带成熟林凋落物生产和分解及其与环境因子的关系. 生态学报, 2013, 33(15): 4818-4825. |
| [19] | 唐仕姗, 杨万勤, 殷睿, 熊莉, 王海鹏, 王滨, 张艳, 彭艳君, 陈青松, 徐振锋. 中国森林生态系统凋落叶分解速率的分布特征及其控制因子. 植物生态学报, 2014, 38(6): 529-539 |
| [20] | Liski J, Nissinen A, Erhard M, Taskinen O. Climatic effects on litter decomposition from arctic tundra to tropical rainforest. Global Change Biology, 2003, 9(4): 575-584. |
| [21] | Liu C, Wang X P, Wu X, Dai S, He J S, Yin W L. Relative effects of phylogeny, biological characters and environments on leaf traits in shrub biomes across central Inner Mongolia, China. Journal of Plant Ecology, 2013, 6(3): 220-231. |
| [22] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H, Diemer M. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. |
| [23] | He J S, Wang X P, Schmid B, Flynn D F B, Li X F, Reich P B, Fang J Y. Taxonomic identity, phylogeny, climate and soil fertility as drivers of leaf traits across Chinese grassland biomes. Journal of Plant Research, 2010, 123(4): 551-561. |
| [24] | Santiago L S. Extending the leaf economics spectrum to decomposition: evidence from a tropical forest. Ecology, 2007, 88(5): 1126-1131. |
| [25] | Chapin III F S, Zavaleta E S, Eviner V T, Naylor R L, Vitousek P M, Reynolds H L, Hooper D U, Lavorel S, Sala O E, Hobbie S E. Consequences of changing biodiversity. Nature, 2000, 405(6783): 234-242. |
| [26] | Reitalu T, Seppä H, Sugita S, Kangur M, Koff T, Avel E, Kihno K, Vassiljev J, Renssen H, Hammarlund D. Long-term drivers of forest composition in a boreonemoral region: the relative importance of climate and human impact. Journal of Biogeography, 2013, 40(8): 1524-1534. |
| [27] | Prescott C E. The influence of the forest canopy on nutrient cycling. Tree Physiology, 2002, 22(15/16): 1193-1200. |
| [28] | Prescott C E, Hope G D, Blevins L L. Effect of gap size on litter decomposition and soil nitrate concentrations in a high-elevation spruce fir forest. Canadian Journal of Forest Research, 2003, 33(11): 2210-2220. |
| [29] | McCarthy J. Gap dynamics of forest trees: a review with particular attention to boreal forests. Environmental Reviews, 2001, 9(1): 1-59. |
| [30] | Wang X P, Fang J Y, Sanders N J, White P S, Tang Z Y. Relative importance of climate vs local factors in shaping the regional patterns of forest plant richness across northeast China. Ecography, 2009, 32(1): 133-142. |
| [31] | 张新平, 王襄平, 朱彪, 宗占江, 彭长辉, 方精云. 我国东北主要森林类型的凋落物产量及其影响因素. 植物生态学报, 2008, 32(5): 1031-1040. |
| [32] | Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter. Oikos, 2004, 104(2): 230-246. |
| [33] | Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 1963, 44(2): 322-331. |
| [34] | 张德强, 叶万辉, 余清发, 孔国辉, 张佑倡. 鼎湖山演替系列中代表性森林凋落物研究. 生态学报, 2000, 20 (6): 938-944. |
| [35] | 黄建辉, 陈灵芝, 韩兴国. 森林生态系统中凋落物分解的研究进展//李承森. 植物科学进展Vol. 1. 北京: 高等教育出版社, 1998: 218-316. |
| [36] | Ndagurwa H G T, Dube J S, Mlambo D. Decomposition and nutrient release patterns of mistletoe litters in a semi-arid savanna, southwest Zimbabwe. Austral Ecology, 2015, 40(2): 178-185. |
| [37] | 方精云. 地理要素对我国温度分布影响的数量评价. 生态学报, 1992, 12(2): 97-104. |
| [38] | Wang X P, Tang Z Y, Fang J Y. Climatic control on forests and tree species distribution in the forest region of Northeast China. Journal of Integrative Plant Biology, 2006, 48(7): 778-789. |
| [39] | Heikkinen R K, Luoto M, Kuussaari M, Pöyry J. New insights into butterfly-environment relationships using partitioning methods. Proceedings of the Royal Society B: Biological Sciences, 2005, 272(1577): 2203-2210. |
| [40] | Wang X P, Tang Z Y, Shen Z H, Zheng C Y, Luo J C, Fang J Y. Relative influence of regional species richness vs local climate on local species richness in China's forests. Ecography, 2012, 35(12): 1176-1184. |
| [41] | 刘颖, 武耀祥, 韩士杰, 林鹿. 长白山四种森林类型凋落物分解动态. 生态学杂志, 2009, 28(3): 400-404. |
| [42] | Trofymow J A, Moore T R, Titus B, Prescott C, Morrison I, Siltanen M, Smith S, Fyles J, Wein R, Camiré C, Duschene L, Kozak L, Kranabetter M, Visser S. Rates of litter decomposition over 6 years in Canadian forests: influence of litter quality and climate. Canadian Journal of Forest Research, 2002, 32(5): 789-804. |
| [43] | Gholz H L, Wedin D A, Smitherman S M, Harmon M E, Parton W J. Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition. Global Change Biology, 2000, 6(7): 751-765. |
| [44] | Vitousek P M, Turner D R, Parton W J, Sanford R L. Litter decomposition on the Mauna Loa environmental matrix, Hawai'i: patterns, mechanisms, and models. Ecology, 1994, 75(2): 418-429. |
| [45] | Latter P M, Howson G, Howard D M, Scott W A. Long-term study of litter decomposition on a Pennine peat bog: which regression?. Oecologia, 1997, 113(1): 94-103. |
| [46] | Austin A T, Vitousek P M. Nutrient dynamics on a precipitation gradient in Hawai'i. Oecologia, 1998, 113(4): 519-529. |
| [47] | Zhang Q S, Zak J C. Effects of gap size on litter decomposition and microbial activity in a subtropical forest. Ecology, 1995, 76(7): 2196-2204. |
| [48] | Ritter E. Litter decomposition and nitrogen mineralization in newly formed gaps in a Danish beech (Fagus sylvatica) forest. Soil Biology and Biochemistry, 2005, 37(7): 1237-1247. |
| [49] | Terborgh J. The vertical component of plant species diversity in temperate and tropical forests. The American Naturalist, 1985, 126(6): 760-776. |
| [50] | 周以良. 中国东北植被地理. 北京: 科学出版社, 1997: 289-299. |
| [51] | Sariyildiz T. Effects of gap-size classes on long-term litter decomposition rates of beech, oak and chestnut species at high elevations in Northeast Turkey. Ecosystems, 2008, 11(6): 841-853. |
| [52] | Prescott C E, Zabek L M, Staley C L, Kabzems R. Decomposition of broadleaf and needle litter in forests of British Columbia: influences of litter type, forest type, and litter mixtures. Canadian Journal of Forest Research, 2000, 30(11): 1742-1750. |
| [53] | Perry D A, Choquette C, Schroeder P. Nitrogen dynamics in conifer-dominated forests with and without hardwoods. Canadian Journal of Forest Research, 1987, 17(11): 1434-1441. |
| [54] | Anderson J M. Spatiotemporal effects of invertebrates on soil processes. Biology and Fertility of Soils, 1988, 6(3): 216-227. |
| [55] | Murphy K L, Klopatek J M, Klopatek C C. The effects of litter quality and climate on decomposition along an elevational gradient. Ecological Applications, 1998, 8(4): 1061-1071. |
| [56] | Vivanco L, Austin A T. Tree species identity alters forest litter decomposition through long-term plant and soil interactions in Patagonia, Argentina. Journal of Ecology, 2008, 96(4): 727-736. |