 2016, Vol. 36
2016, Vol. 36文章信息
- 王静娅, 王明亮, 张凤华
- WANG Jingya, WANG Mingliang, ZHANG Fenghua
- 干旱区典型盐生植物群落下土壤微生物群落特征
- Soil microbial properties under typical halophytic vegetation communities in arid regions
- 生态学报, 2016, 36(8): 2363-2372
- Acta Ecologica Sinica, 2016, 36(8): 2363-2372
- http://dx.doi.org/10.5846/stxb201410061958
-
文章历史
- 收稿日期: 2014-10-06
- 网络出版日期: 2015-08-21






2. 新疆兵团技术市场办公室, 乌鲁木齐 830002
2. Xinjiang corps technology market office, Urumqi 830002, China
土壤微生物是生态系统中重要的组成部分,而土壤微生物群落多样性作为描述其生态特征的关键指标,近来成为生态领域研究的热点问题。有研究指出,在荒漠生态系统中,植物群落类型对土壤微生物群落结构及功能多样性有着重要的影响[1]。同时,不同植物群落类型下土壤微生物群落结构和功能的差异也影响着植被凋落物的分解,物质能量的转换方式[2],包括碳、氮循环过程。由此可见,对干旱荒漠区不同植物群落对土壤微生物群落特征的影响进行研究能够很好了解当地的土壤环境状况,为荒漠生态系统植物群落的保育与恢复提供理论依据。在土壤微生物群落多样性的研究方法中,Biolog分析法作为研究土壤微生物群落结构和功能多样性的一种简单快速且灵敏度高,分辨力强的方法[3],可以一定程度上反映土壤碳源转化和土壤微生物多样性情况,被广泛运用于土壤微生物群落多样性对不同生境变化的响应方面。
目前,国内外对土壤微生物多样性的研究主要集中在环境变化和人为干扰方面,如不同农作模式[4],施肥处理[5],环境污染[6],土地退化[7]等。而在自然生态系统方面中,国外也有较多研究[8],但是在国内研究则较少,主要集中在森林生态系统方面[9, 10]。有研究指出[9],武夷山不同海拔植被带土壤微生物群落多样性随着海拔上升而下降,可能是生物量、林分凋落物、土壤养分、微小动物、植物根系等多种因素共同作用的结果。也有研究指出[10],江西退化红壤4 种人工森林土壤微生物对碳源利用能力显著差异,其中阔叶混交林碳源代谢能力最强,说明在退化土壤重建森林时,可以优先考虑阔叶混交林。而在干旱荒漠区,目前对自然生态系统中土壤微生物群落的研究则较匮乏[11]。不同荒漠盐生植物群落下土壤微生物多样性以及碳源利用差异性值得进一步揭示。
玛纳斯河是干旱区准噶尔盆地最大的内陆河,受气候、地形及灌溉等的影响,流域内不同地貌盐分种类和含量不同,其中玛纳斯河扇缘带属于硫酸盐土壤,盐分含量也最重[12],在特定的自然环境条件下发育着丰富的盐生植物[13],这些植被对干旱区绿洲荒漠复合生态系统的平衡起着积极的作用[14]。然而随着人工绿洲的不断建立与扩大,加之水土资源不合理利用,使得绿洲外围盐生植被带日趋衰落,其生态效益成为人们关注的焦点。本研究以玛河流域冲积扇扇缘带为研究区,综合考虑分别选取了生长在该区的白刺(Nitraria tangutorum Bobr.)、绢蒿(Seriphidium kaschgaricum (Krasch.) Poljak.)、花花柴(Karelinia caspia (Pall. ) Less.)、雾冰藜(Bassia dasyphylla (Fisch. et C. A. Mey.) Kuntze.)、梭梭(Haloxylon ammodendron (C. A. Mey.) Bunge.)、柽柳(Tamarix ramosissima Lour.)6种典型盐生植物群落,分析了不同盐生植物群落下土壤微生物群落功能多样性、碳源利用差异以及地上植被多样性与土壤微生物多样性相互作用关系,为了解该区域土壤环境状况,探讨地上植物分布与土壤微生物群落组成、结构和功能之间的关系提供科学依据,也为该区域盐生植被的保育与恢复提供依据。
1 材料与方法 1.1 研究区概况研究区位于玛河流域扇缘带,地理位置东经 86°03′—86°08′,北纬 44°64′—44°71′。该地区属温带大陆性干旱半干旱气候区,具有冬季严寒,夏季酷热,日照充足,干旱少雨等特点。年平均气温6—7 ℃之间,最高气温在7月,最热月温度在24—36 ℃,最冷月温度在-18—-19.5 ℃;农业主要以灌溉为主,年降水量平均值140—180 mm,多集中在4—5月。多年平均蒸发量为1521.43 mm,蒸发主要集中在4—9月,约占全年蒸发量的88%。该区地下水位高,流动不畅,土质多系盐碱土和荒漠灰钙土,土壤含盐量高,改良条件差。研究区植物种类贫乏,且生长稀疏,以旱生、超旱生、耐盐碱的荒漠植被占优势,植物群落的主要物种为白刺、绢蒿、花花柴、雾冰藜、梭梭、柽柳、骆驼刺(Alhagi camelorum Fisch.)、小叶碱蓬(Suaeda microphylla (C. A. Mey.) Pall.)、盐节木(Halocnemum strobilaceum (Pall.) Bieb.)、小獐毛(Aeluropus pungens (M. Bieb. ) C. Koch.)、叉毛蓬(Petrosimonia sibirica (Pall.) Bunge.)、盐爪爪(Kalidium foliatum (Pall.) Moq.)、补血草(Limonium sinense (Girard) Kuntze.)等。
1.2 样地设置与采样方法选取玛河流域扇缘带弃耕20a以上的天然盐碱荒地,根据样地植物群落优势种的不同,综合物种组成和群落结构等因素,选择了坡向、坡度和海拔高度基本一致的白刺、绢蒿、花花柴、雾冰藜、梭梭和柽柳6种典型盐生植物群落。采用样方法(每个群落设置3个20 m×20 m的样方)对6个群落进行植被调查,调查包括植物种类、株高、冠幅、植被多样性等。样地植物种类描述见表 1。
| 样地类型 Sample types | 物种组成(按优势度排序) Species composition(Order by the degree of dominance) | 优势物种平均株高 Average height of dominant species/cm | 优势物种平均冠幅 Average canopy of dominant species/(cm×cm) |
| N:白刺Nitraria tangutorum Bobr.;S:绢蒿 Seriphidium kaschgaricum (Krasch.) Poljak;K:花花柴 Karelinia caspia (Pall. ) Less;B:雾滨藜 Bassia dasyphylla (Fisch. et C. A. Mey.);H:梭梭 Bassia dasyphylla (Fisch. et C. A. Mey.);T:柽柳 Tamarix ramosissima Lour | |||
| N | 白刺、疏叶骆驼刺、小叶碱蓬、盐节木 | 62±10.32 | 134×152 |
| S | 绢蒿、小叶碱蓬、柽柳、白刺、疏叶骆驼刺、花花柴、小獐茅 | 25±8.44 | 13×12 |
| K | 花花柴、雾滨藜、小叶碱蓬、叉毛蓬、白刺 | 32±5.54 | 38×47 |
| B | 雾滨藜、盐爪爪、花花柴 | 3±1.51 | — |
| H | 梭梭、盐爪爪、补血草、小獐茅、盐节木 | 76±13.52 | 150×110 |
| T | 柽柳、花花柴、小獐茅 | 57±10.81 | 110×80 |
于2013年5月中旬,分别在各个群落样地内设置 3个 20 m×20 m 的样方,在每个样方用直径为 7.5 cm 的土钻以 S 型分别采集 5 份土壤( 0—20 cm) 进行混合,装入封口袋并放入装有冰块的保温箱带回实验室。过2 mm筛后,一份保存在4 ℃用于微生物群落功能多样性测定,另一份风干后用于土壤理化性质测定。土壤基本理化性质见表 2。
| 样地类型 Plot type | 含水率 Water content/% | 容重 Bulk density/(g/cm) | pH | 电导率 Electrical conductivity (s/m) | 有机质 Organic matter/(g/kg) | 全氮 Totalnitrogen/(g/kg) | 碱解氮 Alkali-hydrolyzable nitrogen/(mg/kg) | 速效磷 Available phosphorus/(mg/kg) | 速效钾 Available potassium/(mg/kg ) |
| 同一列数据中不同字母表示差异显著(P<0.05) | |||||||||
| N | 6.43bc | 1.38a | 7.94d | 1.67c | 7.22c | 0.36d | 30.89a | 8.19c | 387.87c |
| S | 5.99bc | 1.42a | 7.97d | 1.84c | 9.05bc | 0.38d | 23.11b | 9.40bc | 381.02d |
| K | 6.28bc | 1.01c | 8.86b | 6.96a | 10.22b | 0.50bc | 18.08c | 13.50a | 387.87c |
| B | 7.33b | 1.42a | 8.53c | 3.36d | 9.90b | 0.48c | 18.67c | 14.04a | 445.10b |
| H | 12.07a | 1.15b | 9.08a | 4.65b | 13.04a | 0.52b | 18.13c | 10.27b | 467.31a |
| T | 4.80c | 1.37a | 8.15b | 1.68c | 13.22a | 0.57a | 27.75a | 12.93a | 337.55e |
土壤pH值采用电极电位法测定,有机碳(SOC)采用重铬酸钾氧化-外加热法测定,全氮(TN)采用半微量开氏法测定,碱解氮(AN)采用扩散吸收法测定,速效磷(AP)采用 NaHCO3提取-钼锑抗显色-紫外分光光度法测定,速效钾(AK)采用NH4Ac浸提-原子吸收法测[15]。
1.3.2 土壤微生物功能多样性测定土壤微生物碳源多样性采用Biolog生态测试板(ECO)测定。称取相当于10 g烘干土重的新鲜土样加入内有100 mL无菌缓冲液水(NaCl 0.67%,KH2PO2 0.024%,Na2HPO4· 7H2O 0.086%,调pH值)的三角瓶中,加无菌棉花塞,在200 r/min下振荡30 min,按1∶100比例稀释液接种生态测试板,接种量为150 μL,将接种好的测试板黑暗下28 ℃连续培养,每隔12 h用BIOLOG自动读数装置在595 nm下读数至到144 h[16, 17]。
1.4 数据处理方法土壤微生物群落功能多样性通过其对碳源利用能力来衡量,其中平均颜色变化率(Average well color development,AWCD)反映的微生物总体活性,计算方法如下[18]:
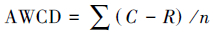
式中,C为每孔底物的OD值,R为对照孔OD值,当C-R为负值时记为0,表示对应孔的底物碳源没有被利用[19]。
土壤微生物功能多样性测定与植物生态学中的计算公式相同。丰富度指数(S)指被利用的碳源总数目,即颜色变化孔数,光密度值0.25是其颜色变化的最低阈值[20]。相关公式如下:
丰富度指数采用Margalef指数(M)
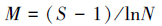
多样性指数采用Shannon-Weinner指数(H)
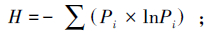
优势度指数采用Simpson 指数(D)
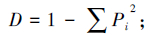
均匀度指数采用Pielou指数(E)
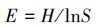
式中,Pi为第i孔的光密度值与对照孔的光密度值之差与微平板上所有孔的光密度总差的比值,即Pi=(C-R)/∑(C-R);在植物多样性上,式中Pi=Ni/N,N为样地植物重要值总和;Ni为样方中第i中植物的重要值,Pi为重要值比例,S为样地群落物种数。
本研究采用AWCD值在96 h的数据计算Biolog板中微生物群落代谢类型,如培养基指数的计算、主成分分析(PCA)、单种培养基与主成分得分系数的相关分析等。此时AWCD值接近0.6,这样可以比较颜色变化率基本相同情况下Biolog板的碳源利用情况,而不考虑接种密度带来的差异[21]。应用SPSS 11.5软件对数据进行方差分析( one-way ANOVA),并用LSD法进行差异显著性检验,利用Pearson相关系数评价不同因子间的相关关系。
2 结果与分析 2.1 土壤微生物群落总碳源利用的动力特征将平均颜色变化率(AWCD)作为微生物整体活性的有效指标。AWCD值的变化速度(斜率)和最终能达到的AWCD值反映了土壤微生物利用某一碳源物质的能力。如图 1所示,各群落土壤AWCD值均随着时间的增加而升高,最终进入缓慢平稳期。从整体来看变化趋势为:梭梭>花花柴>白刺>绢蒿>柽柳>雾冰藜。通过对96h AWCD值方差分析(表 3),可见梭梭群落显著高于其他群落,为0.59,其次是花花柴群落,为0.34,而白刺,绢蒿,柽柳群落差异不显著,雾冰藜群落显著最低(P<0.05)。由此可见,不同群落之间的土壤碳源利用能力差异明显,其中梭梭群落土壤碳源利用能力最强,其次是花花柴群落,而雾冰藜群落最弱。

|
| 图 1 培养过程中AWCD变化 Fig.1 AWCD dynamics with incubation time N:白刺Nitraria tangutorum Bobr.;S:绢蒿 Seriphidium kaschgaricum (Krasch.) Poljak;K:花花柴 Karelinia caspia (Pall. ) Less;B:雾滨藜 Bassia dasyphylla (Fisch. et C. A. Mey.);H:梭梭 Bassia dasyphylla (Fisch. et C. A. Mey.);T:柽柳 Tamarix ramosissima Lour。 |
| 群落类型 Community type | 平均颜色变化率 Average well color development(AWCD) | 多样性指数 Shannon index(H) | 优势度指数 Simpson index(D) | 均匀度指数 Pielou index(E) |
| N | 0.182±0.054c | 2.330±0.494a | 0.856±0.073a | 1.275±0.170b |
| S | 0.121±0.003cd | 2.625±0.358a | 0.897±0.038a | 1.794±0.299a |
| K | 0.340±0.066b | 2.012±0.124a | 0.860±0.026a | 1.114±0.164b |
| B | 0.025±0.025d | 1.277±0.733b | 0.581±0.268b | 0.833±0.314b |
| H | 0.568±0.112a | 2.646±0.144a | 0.917±0.010a | 0.959±0.017b |
| T | 0.074±0.005cd | 2.771±0.121a | 0.929±0.009a | 0.822±0.421b |
对土壤微生物培养96 h的碳源利用程度数据进行多样性指数分析,Shannon指数(H)可以表征土壤中微生物群落多样性,Simpson指数(D)可以表征土壤中微生物群落优势度,Shannon指数(E)可以表征土壤中微生物群落均匀度。由表 3可见,在多样性指数和优势度指数上,除雾冰藜群落显著低于其他群落以外(P<0.05),其他群落差异不显著(P>0.05),说明了除雾冰藜群落,其他群落对常见菌种影响不大。在均匀度指数上,绢蒿显著高于其他群落,为1.794,其他群落之间差异不显著(P>0.05),说明了除绢蒿群落,其他群落菌群分布较均匀。
2.3 土壤微生物群落利用碳源的类型按化学基团的性质将生态板板上的31种碳源分成6类,分别是氨基酸类、碳水化合物类、羧酸类、聚合物类和胺类。由图 2可知,不同植物群落土壤微生物对6类碳源的利用差异显著(P<0.05)。除胺类,梭梭群落土壤微生物对其他5种碳源利用率均显著高于其他群落(P<0.05);花花柴群落土壤微生物对酚酸类、氨基酸类、胺类的利用率较高;白刺和绢蒿群落对聚合物类和碳水化合物类利用率稍高;雾冰藜和柽柳群落土壤微生物对6类碳源的利用率显著最低(P<0.05)。其中,梭梭群落土壤微生物对羧酸类、聚合物类、碳水化合物类、酚酸类、氨基酸类、胺类的利用率分别是雾冰藜群落的3.3、2.5、5.1、3.9、3.8、2.1倍。由此可见,在6种植物群落中,梭梭群落土壤微生物对主要碳源的利用率较高,其次是花花柴群落,而柽柳和雾冰藜群落的利用率最低,与培养过程中AWCD值变化规律基本一致。总体而言,碳水化合物类和氨基酸类是各植物群落土壤微生物的主要碳源,其次为酚酸类,聚合物类和羧酸类,胺类碳源的利用率最低。

|
| 图 2 不同植物群落对土壤微生物群落6类碳源利用率的影响 Fig.2 Effects of different vegetation communities on substrate utilization by soil microbial community Ⅰ: 羧酸类;Ⅱ: 聚合物类;Ⅲ: 碳水化合物类;Ⅳ: 酚酸类;Ⅴ: 氨基酸类;Ⅵ: 胺类; 不同字母间表示差异显著 |
土壤微生物多样性反映了群落总体的变化,但未能反映微生物群落代谢的详细信息,研究土壤微生物对不同碳源利用能力的差异,有助于更全面地了解微生物群落代谢功能特性[22]。应用主成分分析( Principal Component Analysis,PCA)在31种因子中提取的2个主成分因子,由图 3可见,主成分1(PC1)可以解释所有变量方差的41.51%,主成分2(PC2)可以解释所有变量方差的25.35%,累计贡献率达到66.86%。图 3中各群落在PC1和PC2上分布较分散,说明了各群落碳源利用差异较大,PC1和PC2基本上能够区分不同植物群落土壤微生物的群落特征。由表 4可见,通过对不同植物群落土壤微生物在PC1和PC2上的得分进行方差分析,在PC1上,柽柳群落得分显著高于其他群落(P<0.05),白刺,绢蒿和雾冰藜群落得分较低且差异不显著(P>0.05),而花花柴和梭梭群落显著低于其他群落(P<0.05)。在PC2上,梭梭群落得分显著高于其他群落(P<0.05),雾冰藜和柽柳群落显著最低(P<0.05),而其他群落之间差异不显著(P>0.05)。由表 5可见,进一步将主成分得分系数与单一碳源AWCD值做相关分析,选择相关性较高的碳源,在31种碳源中,与PC1相关的有17个,其中碳水化合物类6个,羧酸类5个,氨基酸类和酚酸类各2个,聚合物类和胺类各1个,对PC1 起分异作用的主要碳源是碳水化合物类。与PC2相关的有16个,氨基酸类5个,碳水化合物类、聚合物类和羧酸类各3个,胺类、酚酸类各1个,对PC2起分异作用的主要碳源是氨基酸类。说明了在主成分分离中起主要贡献作用的是碳水化合物类和氨基酸类碳源。

|
| 图 3 不同盐生植物群落土壤微生物碳源利用主成分分析 Fig.3 Principal component analysis of carbon utilization of soil microbial community in different vegetation communities |
| 主成分 Principal component | 群落类型 Community type | 得分 Score | P<0.05 |
| 主成分1(PC1) | N | 0.044±0.086 | b |
| 主成分2(PC2) | S | 0.107±0.012 | ab |
| K | -0.073±0.064 | c | |
| B | 0.134±0.052 | ab | |
| H | -0.061±0.006 | c | |
| T | 0.153±0.039 | a | |
| N | 0.125±0.061 | b | |
| S | 0.093±0.013 | b | |
| K | 0.102±0.061 | b | |
| B | -0.060±0.009 | c | |
| H | 0.212±0.020 | a | |
| T | -0.020±0.031 | c |
| 主成分1 PC1 | r | 主成分2 PC2 | r |
| 碳水化合物Carbohydrates | 碳水化合物Carbohydrates | ||
| α-甲基-D-葡糖糖苷α-Methyl-D-Glucoside L | -0.579 | α-甲基-D-葡糖糖苷α-Methyl-D-Glucoside L | -0.719 |
| D-木糖D-Xylose | 0.746 | D-甘露醇D-Mannitol | -0.673 |
| i-赤藻糖苷i-Erythritol | 0.629 | α-D-乳糖α-D-Lactose | 0.520 |
| N-乙酰基-D-葡糖糖胺A-Acetyl-D-Glucosamine | -0.938 | 氨基酸类Amino acids | |
| D-纤维二糖D-Cellobiose | -0.884 | L-天冬酰氨酸L-Asparagine | 0.526 |
| α-D-乳糖α-D-Lactose | 0.752 | L-苯基丙氨酸L-Phenylalamine | -0.825 |
| 氨基酸类Amino acids | L-丝氨酸L-serine | 0.876 | |
| L-精氨酸L-Arginine | 0.765 | L-苏氨酸L-Threomine | -0.703 |
| L-苯基丙氨酸L-Phenylalamine | 0.565 | 甘氨酰-L-谷氨酸Glycose-L-Glutamic Acid | 0.778 |
| 聚合物类Polymers | 聚合物类Polymers | ||
| α-环式糊精α-Cyclodextrin | -0.991 | 吐温40 Tween-40 | -0.758 |
| 羧酸类Carboxylic acids | 吐温80 Tween-80 | -0.866 | |
| D-半乳糖γ-内脂D-Galactonic Acid-γ-Lactone | 0.954 | 肝糖Glycogen | 0.660 |
| D-半乳糖醛酸D-Galacturonic Acid | 0.749 | 羧酸类Carboxylic acids | |
| γ-羟基丁酸γ-Hydroxybutyric Acid | 0.662 | D-半乳糖醛酸D-Galacturonic Acid | -0.528 |
| 衣康酸Itaconic Acid | 0.540 | 2-羟基苯甲酸2-Hydroxy-Benzoic Acid | -0.559 |
| D-苹果酸D-Malic Acid | -0.876 | 4-羟基苯甲酸4-Hydroxy-Benzoic Acid | 0.636 |
| 胺类Amines | 胺类Amines | ||
| 苯乙基胺Phenylethylamine | -0.754 | 腐胺Putrescine | 0.884 |
| 酚酸类Phenolic acids | 酚酸类Phenolic acids | ||
| 丙酮酸甲酯Pyruvic Acid Methyl Eester | 0.523 | 丙酮酸甲酯Pyruvic Acid Methyl Eester | 0.700 |
| D,L-α-磷酸甘油D,L-α-Glycrol Phosphate | -0.916 |
通过表 6可以看出,绢蒿,梭梭和柽柳群落的丰富度指数显著高于其他群落(P<0.05)。在多样性指数和优势度指数上,白刺,绢蒿,梭梭和柽柳群落的差异不显著,但显著高于其他群落(P<0.05)。在均匀度指数上,白刺显著高于其他群落(P<0.05),其他群落之间差异不显著(P>0.05)。从总体上看,丰富度指数,多样性指数和优势度指数变化趋势相同,均为绢蒿,梭梭和柽柳群落较为优势,可见该群落植被多样性较丰富。通过表 7相关性分析表明,植物丰富度指数,多样性指数和优势度指数与土壤微生物多样性指数均呈极显著正相关(P<0.01),为别为0.851**,0.877**,0.738**,与土壤微生物优势度指数也呈极显著正相关,为0.786**。说明了土壤微生物多样性与植物多样性呈正相关关系,说明了在一般情况下,植物多样性越丰富,土壤微生物多样性越丰富。
| 群落类型 Community type | 丰富度指数 Margalef index (M) | 多样性指数 Shannon index (H) | 优势度指数 Simpson index (D) | 均匀度指数 Pielou index (E) |
| N | 0.691±0.224b | 1.002±0.314a | 0.563±0.325a | 0.723±0.224a |
| S | 1.171±0.324a | 1.052±0.325a | 0.518±0.223a | 0.479±0.315b |
| K | 0.693±0.224b | 0.679±0.225b | 0.306±0.344b | 0.379±0.224c |
| B | 0.420±0.311c | 0.569±0.221b | 0.286±0.3224b | 0.411±0.223b |
| H | 1.090±0.335a | 0.949±0.322a | 0.474±0.223a | 0.488±0.322b |
| T | 0.857±0.322a | 0.917±0.223a | 0.469±0.224a | 0.512±0.221b |
| 项目 Items | 丰富度指数 Margalef index | 多样性指数 Shannon index | 优势度指数 Simpson index | 均匀度指数 Pielou index |
| *P<0.05; ** P<0.01 | ||||
| AWCD | 0.428 | 0.170 | 0.079 | -0.077 |
| 多样性指数 Shannon index | 0.851** | 0.877** | 0.783** | 0.375 |
| 优势度指数 Simpson index | 0.786** | 0.772* | 0.659* | 0.294 |
| 均匀度指数 Pielou index | 0.541 | 0.579* | 0.492 | 0.204 |
植被类型对土壤微生物活性、功能类群具有重要影响[23]。研究结果表明:土壤平均颜色变化率(AWCD)随培养时间延长而逐渐增加,大小顺序依次为:梭梭>花花柴>白刺>绢蒿>柽柳>雾冰藜。且差异显著。不同植物群落土壤微生物对主要碳源利用率的趋势也与此规律相一致。说明了梭梭群落土壤微生物群落活性,以及碳源利用能力均较强。6类碳源中,碳水化合物类和氨基酸类是6种植被土壤微生物的主要碳源,其次是酚酸类,聚合物类和羧酸类,胺类的利用率最低。主成分分析表明,对PC1和PC2起分异作用的主要碳源分别为碳水化合物类和氨基酸类。说明了碳水化合物类和氨基酸类是6种植物群落的微生物群落变化的敏感碳源。土壤微生物群落在Shannon指数,Simpson指数上,除雾冰藜群落,其他植物群落的土壤微生物多样性差异不明显。在Pielou指数上,除绢蒿群落,其他群落的微生物群落分布较均匀。植物群落丰富度指数,多样性指数和优势度指数变化趋势相同,均为绢蒿,梭梭和柽柳群落较为优势。相关性分析表明,植物丰富度、优势度、多样性指数与土壤微生物优势度、多样性指数均呈显著正相关关系(P<0.05),说明了植物多样性越丰富,土壤微生物多样性越丰富。总体来说,干旱区不同盐生植物群落对土壤微生物群落多样性具有重要影响。其中,梭梭群落的土壤微生物群落具有较强的微生物总体活性和功能多样性。碳水化合物类和氨基酸类是6种植物群落的微生物群落变化的敏感碳源。
本研究中,不同群落的平均颜色变化率(AWCD值)大小顺序依次为:梭梭>花花柴>白刺>绢蒿>柽柳>雾冰藜。说明了梭梭群落土壤微生物群落活性,以及碳源利用能力均较强。相关研究表明,梭梭和柽柳群落具有较强的“肥岛”效应,且土壤有机质含量均高于研究区周围土壤[24],从样地基本理化性质(表 2)也可以看出二者的有机质含量均最高,但是土壤有机碳含量高,不一定被微生物所利用的碳源较高。有研究指出影响微生物量碳含量的因素一是有机质输入量的多少,二是有机质活性与易矿化程度[25]。可能柽柳群落土壤中有机碳活性较低,造成供给土壤微生物可利用的碳源较少,所以该群落整体微生物碳源利用能力较弱。同时在样地描述中可见(表 1),柽柳群落物种少,冠幅低,而梭梭群落物种较丰富,冠幅大,有研究表明[26],植物群落的生物量、群落高度、群落盖度和群落物种多样性与土壤养分分布均表现一致的正相关关系。由此可见。梭梭群落碳源利用能力要强于柽柳群落。而雾冰藜群落的AWCD值最低,这是因为该群落植株矮小,物种单一,且为一年生草本植物,因此对于土壤输入的碳源较低,所以微生物群落活性较低。
不同植物群落土壤微生物对6类碳源的利用差异显著(P<0.05)。说明了不同植物群落通过根系等影响着土壤微生物群落对不同碳源的利用。研究表明[27],不同植物群落下土壤微生物群落对各碳源具有不同的利用能力,是因为这些碳源在不同根系土壤中的有效性不一样,后者能影响微生物群落中利用此类碳源的微生物的生长。本研究中,碳水化合物类和氨基酸类是6种植物群落的主要碳源,也是微生物群落变化的敏感碳源,说明了两种化合物是土壤中广泛存在且能被各种微生物很好利用的碳源种类,如果向土壤中增加此碳源会迅速改变微生物群落的结构和功能。梭梭群落土壤微生物对胺类利用率较低,其他碳源均较高,说明了土壤中胺类化合物可能含量很低,而其他5种碳源分布较多,可能是因为培养过程中微生物种群的竞争机制,或者是微生物对各类底物的适应性不同引起的;花花柴群落土壤微生物对酚酸类、氨基酸类、胺类的利用率较高,说明了这些化合物可能在这些土壤中含量较高,可能是土壤上的植物释放出更多的这些化合物进入土壤;雾冰藜和柽柳群落土壤微生物对6类碳源的利用率显著最低,说明了这两个群落土壤微生物总体活性较低,对于各类碳源利用率较差。
研究表明[28],植物群落的异质性可显著改变地下生物群落结构。用Shannon指数和Simpson指数是研究群落物种数及其个体数和分布均匀程度的综合指标,是目前应用最为广泛的群落多样性指数之一。在本研究中,也用来度量植物多样性和土壤微生物多样性。通过相关性分析表明了植被群落越丰富,土壤微生物多样性越高。这是因为植被物种越丰富,其根系分泌物种类越多,会使根系附近土壤微生物群落增加,同时植被凋落物也多,导致土层腐殖质增加,养分条件好,促进整体碳氮的循环,为微生物生长提供较高的动力。与前人研究结果一致[29],植物群落是土壤微生物赖以生存的营养物质和能量来源,植物群落种类多样性与土壤微生物种类多样性呈显著正相关关系。但是这也并非绝对的规律。研究中虽然梭梭群落的多样性显著高于花花柴群落,但是微生物多样性差异并未达到一个显著水平。在研究中,土壤AWCD值与植物多样性相关性不显著,说明了土壤微生物群落总体活性和植物多样性无关。土壤AWCD值反映了土壤微生物群落的总碳源利用和活性,而不是微生物数量的多少。而植物多样性越丰富,土壤微生物碳源种类越多,但是不一定土壤微生物群落活性越高。土壤AWCD值与植物群落向土壤输入质量较好的有机质、大量的根系分泌物、较高的可矿化有机质含量,以及土壤物理结构均相关,由此可见对土壤AWCD值的讨论还需从多角度去考虑,而不单单是植物多样性。有较多研究表明[30],微生物群落多样性指数与含水率、有机质、全氮、全磷、全钾等呈正相关关系,可见土壤养分是影响土壤微生物群落多样性的重要因素。在研究中土壤养分特征所示(表 2),柽柳群落含水率最低、容重较大,虽然有机质含量较高,但缺乏保水透气的土壤物理结构,微生物对底物的利用程度相应较低,则AWCD值较低。也有研究表明[31],土壤微生物多样性指数与土壤转化酶,碱性磷酸酶,过氧化氢酶活性之间达到极显著正相关关系,说明土壤酶活与微生物代谢共同影响微生物群落多样性。由此可见,相关研究表明,土壤微生物群落多样性是土壤养分、水热状况、凋落物和根系分泌物等因素综合作用的结果[30],也与土壤类型、气候及人类活动等密切相关[32]。因此对土壤微生物多样性的探讨还需从多角度去考虑。
本研究仅利用Biolog法分析干旱区不同盐生植物群落土壤微生物多样性和碳源利用差异,取得了一定结果。不同植物群落土壤对碳源的优势利用种类和利用程度也许指示土壤中碳源的有效性和可获得性的差异。但是,Biolog法只能表征土壤中快速生长或富营养微生物活性,不能反映土壤中生长缓慢的微生物活性,且无法直接获取微生物群落结构的详细信息[30]。而在荒漠区,盐生植物群落下的盐碱土必然生长着各种耐盐或嗜盐的微生物,因此,研究要结合现代微生物筛选技术,以及群落结构分析方法来进一步开展土壤微生物群落的研究。同时,此次研究主要针对土壤中整体微生物群落多样性,而土壤团聚体是构成土壤的结构单元,对土壤结构和土壤肥力的形成具有重要作用[33]。在不同土壤团聚体中微生物群落的分布特征目前仍然未知,因此研究将进一步结合土壤团粒结构中微生物群落分布的差异性,来探讨不同盐生植物群落下土壤团聚体中微生物多样性,将进一步有助于在土壤结构单元下来揭示干旱区地上盐生植物群落与地下土壤微生物相互作用关系。
| [1] | Han X M, Wang R Q, Liu J, Wang M C, Zhou J, Guo W H. Effects of vegetation type on soil microbial community structure and catabolic diversity assessed by polyphasic methods in North China. Journal of Environmental Sciences, 2007, 19(10): 1228-1234. |
| [2] | 严海元, 辜夕容, 申鸿. 森林凋落物的微生物分解. 生态学杂志, 2010, 29(9): 1827-1835. |
| [3] | 鲁顺保, 张艳杰, 陈成榕, 徐志红, 郭晓敏. 基于Biolog指纹解析三种不同森林类型土壤细菌群落功能差异. 土壤学报, 2013, 50(3): 618-623. |
| [4] | 顾美英, 徐万里, 茆军, 张志东, 唐光木, 葛春辉. 新疆绿洲农田不同连作年限棉花根际土壤微生物群落多样性. 生态学报, 2012, 32(10): 3031-3140. |
| [5] | 罗希茜, 郝晓晖, 陈涛, 邓婵娟, 吴金水, 胡荣桂. 长期不同施肥对稻田土壤微生物群落功能多样性的影响. 生态学报, 2009, 29(2): 740-748. |
| [6] | 华建峰, 林先贵, 蒋倩, 张华勇, 陈茜, 殷云龙. 砷矿区农田土壤微生物群落碳源代谢多样性. 应用生态学报, 2013, 24(2): 473-480. |
| [7] | 江玉梅, 谢晶, 曹广泮, 陈成龙, 徐志红, 刘苑秋. 江西退化红壤人工重建森林土壤微生物碳源代谢功能研究. 土壤学报, 2014, 51(1): 158-165. |
| [8] | Yoshitake S, Fujiyoshi M, Watanabe K, Masuzawa T, Nakatsubo T, Koizumi H. Successional changes in the soil microbial community along a vegetation development sequence in a subalpine volcanic desert on Mount Fuji, Japan. Plant and Soil, 2013, 364(1/2): 261-272. |
| [9] | 吴则焰, 林文雄, 陈志芳, 方长旬, 张志兴, 吴林坤, 周明明, 沈荔花. 武夷山国家自然保护区不同植被类型土壤微生物群落特征. 应用生态学报, 2013, 24(8): 2301-2309. |
| [10] | Zhang H F, Li G, Song X L, Yang D L, Li Y J, Qiao J, Zhang J N, Zhao S L. Changes in soil microbial functional diversity under different vegetation restoration patterns for Hulunbeier Sandy Land. Acta Ecologica Sinica, 2013, 33(1): 38-44. |
| [11] | 罗倩, 黄宝灵, 唐治喜, 来利明, 魏伟, 郑元润. 新疆盐渍土3种植被类型土壤微生物碳源利用. 应用与环境生物学报, 2013, 19(1): 96-104. |
| [12] | 李玉义, 张凤华, 潘旭东, 陈阜, 赖先齐. 新疆玛纳斯河流域不同地貌类型土壤盐分累积变化. 农业工程学报, 2007, 23(2): 60-64. |
| [13] | 弋良朋, 马健, 李彦. 荒漠盐生植物根际土壤盐分和养分特征. 生态学报, 2007, 27(9): 3565-3571. |
| [14] | 王雪梅, 柴仲平, 塔西甫拉提·特依拜. 西北干旱区典型绿洲盐生植被群落特征及多样性研究. 西南农业学报, 2011, 24(1): 258-262. |
| [15] | 鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000: 30-30. |
| [16] | 孔维栋, 刘可星, 廖宗文. 有机物料种类及腐熟水平对土壤微生物群落的影响. 应用生态学报, 2004, 15(3): 487-492. |
| [17] | Schutter M, Dick R. Shift in substrate utilization potential and structure of soil microbial communities in response to carbon substrates. Soil Biology and Biochemistry, 2001, 33(11): 1481-1491. |
| [18] | Gomez E, Ferreras L, Toresani S. Soil bacterial functional diversity as influenced by organic amendment application. Bioresource Technology, 2006, 97(13): 1484-1489. |
| [19] | Miguel C S, Dulinski M, Tate R L. Direct comparison of individual substrate utilization from a CLPP study: a new analysis for metabolic diversity data. Soil Biology and Biochemistry, 2007, 39(8): 1870-1877. |
| [20] | Garland J. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiology Ecology, 1997, 24(4): 289-300. |
| [21] | Garland J L, Mills A L, Young J S. Relative effectiveness of kinetic analysis vs single point readings for classifying environmental samples based on community-level physiological profiles (CLPP). Soil Biology and Biochemistry, 2001, 33(7/8): 1059-1066. |
| [22] | 孔维栋, 刘可星, 廖宗文, 朱永官, 王碧玲. 不同腐熟程度有机物料对土壤微生物群落功能多样性的影响. 生态学报, 2005, 25(9): 2291-2296. |
| [23] | Fisk M C, Ruether K F, Yavitt J B. Microbial activity and functional composition among northern peatland ecosystems. Soil Biology and Biochemistry, 2003, 35(4): 591-602. |
| [24] | 刘耘华, 杨玉玲, 盛建东, 张瑾. 北疆荒漠植被梭梭立地土壤养分"肥岛"特征研究. 土壤学报, 2010, 47(3): 545-554. |
| [25] | 文倩, 赵小蓉, 陈焕伟, 妥德宝, 林启美. 半干旱地区不同土壤团聚体中微生物量碳的分布特征. 中国农业科学, 2004, 37(10): 1504-1509. |
| [26] | 刘忠宽, 汪诗平, 陈佐忠, 王艳芬, 韩建国. 不同放牧强度草原休牧后土壤养分和植物群落变化特征. 生态学报, 2006, 26(6): 2048-2056. |
| [27] | Grayston S J, Wang S Q, Campbell C D, Edwards A C. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biology and Biochemistry, 1998, 30(3): 369-378. |
| [28] | Zou X M, Ruan H H, Fu Y, Yang X D, Sha L Q. Estimating soil labile organic carbon and potential turnover rates using a sequential fumigation-incubation procedure. Soil Biology and Biochemistry, 2005, 37(10): 1923-1928. |
| [29] | 周桔, 雷霆. 土壤微生物多样性影响因素及研究方法的现状与展望. 生物多样性, 2007, 15(3): 306-311. |
| [30] | 吴则焰, 林文雄, 陈志芳, 方长旬, 张志兴, 吴林坤, 周明明, 陈婷. 中亚热带森林土壤微生物群落多样性随海拔梯度的变化. 植物生态学报, 2013, 37(5): 397-406. |
| [31] | 李凤霞, 王学琴, 郭永忠, 许兴. 宁夏引黄灌区不同盐化程度土壤酶活性及微生物多样性研究. 水土保持研究, 2013, 20(1): 61-65. |
| [32] | 靳正忠, 雷加强, 徐新文, 李生宇, 范敬龙, 赵思峰, 周宏伟, 谷峰. 塔里木沙漠公路防护林土壤微生物生物量与土壤环境因子的关系. 应用生态学报, 2009, 20(1): 51-57. |
| [33] | 刘中良, 宇万太. 土壤团聚体中有机碳研究进展. 中国生态农业学报, 2011, 19(2): 447-455. |