 2016, Vol. 36
2016, Vol. 36文章信息
- 孙会婷, 江莎, 刘婧敏, 郭亚娇, 沈广爽, 古松
- SUN Huiting, JIANG Sha, LIU Jingmin, GUO Yajiao, SHEN Guangshuang, GU Song
- 青藏高原不同海拔3种菊科植物叶片结构变化及其生态适应性
- Structure and ecological adaptability of the leaves of three asteraceae species at different altitudes on the Qinghai-Tibet Plateau
- 生态学报, 2016, 36(6): 1559-1570
- Acta Ecologica Sinica, 2016, 36(6): 1559-1570
- http://dx.doi.org/10.5846/stxb201408251679
-
文章历史
- 收稿日期: 2014-08-25
- 网络出版日期: 2015-07-29






青藏高原平均海拔在4000 m 以上,是世界上平均海拔最高的高原。青藏高原独特的气候特点:高寒缺氧、太阳辐射强烈、CO2分压低、昼夜温差大等使在如此极端环境中生存的植物体必然会形成特殊的形态结构来适应环境[1]。近年来,以特定的生境、连续分布的高山植物为研究点,通过海拔梯度上植物叶片性状与环境因子相互关系来研究环境变化,已成为研究全球气候变化的热点之一[2, 3]。
海拔高度变化对植物叶片解剖结构的影响,近年来已取得了一些研究成果。李有忠等[4]对生长在不同海拔高度的6种植物的叶片内部结构进行了比较,结果表明不同植物叶片对环境中的某个或某些生态因子敏感程度有所不同。马建静等[5]通过对青藏高原高寒草地和内蒙古高原温带草地主要双子叶植物叶片解剖特征的比较研究,进一步探讨了两地植物叶片结构的差异及其对环境因子的响应。施海燕等[6]的研究结果表明:随着海拔高度的升高,植物叶片总厚度、栅栏组织厚度和海绵组织厚度均增加。目前,多数研究是用一些解剖学术语对不同海拔高度的叶片形态解剖结构特征进行了描述,没有解剖学数量指标的测定与分析,而全面系统地对青藏高原不同海拔高度的植物叶片形态解剖结构及其对环境响应的研究鲜为报道。
本文对分布于青藏高原冷龙岭不同海拔高度的3种菊科植物叶片进行了形态及解剖结构的观察,并对解剖学指标进行数量化测定和分析,阐明其叶片解剖结构对随海拔高度变化的响应,并探讨分析其生态适应性,以期为高山植物功能生态学的研究提供参考依据。
1 材料与方法 1.1 材料 1.1.1 研究地概况冷龙岭(101°12′—102°12′ E,37°15′—37°30′ N)位于青海省海北藏族自治州门源回族自治县东北,为祁连山脉东段的第一山,是我国青藏高原高寒区和西北荒漠区的过渡区。冷龙岭最高峰——岗什卡峰海拔5254.5 m,本区多年平均降水量410.0 mm,年均温度为0℃; 8月为最湿润月(多年平均降水量为93 mm);最热月(7月)的年均温度为11.4℃(1951—2006年)[7]。
1.1.2 实验材料在植物生长旺季的7—8月,在冷龙岭东段南麓坡地3600—4400 m不同海拔高度上采集3种菊科植物的新鲜成熟叶片:在每个海拔高度处随机选取10—15株发育程度相近的成熟植株,在植株中部相同的位置采集健康成熟的叶片3—5片,然后在叶片中部以主脉为中心剪取5 mm × 5 mm大小的样品,经FAA固定液(90 mL 70%乙醇+5mL甲醛+5mL冰乙酸)固定,带回实验室后放入4℃的冰箱中保存。每一样品的测量数量为3,共30个视野。植物及采集地点海拔高度信息见表 1。
| 植物种Species | 属Genus | 生活型Life forms | 样地海拔高度Altitude /m |
| 蒲公英Taraxacum mongolicumHand.-Mazz | 蒲公英属 | 多年生草本 | 3600,4000,4200,4400 |
| 火绒草Leontopodium leontopodioides (Willd.) Beauv | 火绒草属 | 多年生草本 | 3600,3800,4400 |
| 美丽风毛菊Saussurea superbaAnthony | 风毛菊属 | 多年生草本 | 3600,3800,4000 |
将FAA固定液中的样品放入4%戊二醛中固定1h,磷酸缓冲液漂洗3次后转移到1%锇酸内再次固定,磷酸缓冲液漂洗后经乙醇系列脱水,自然干燥后的样品固定于样品台上,喷金后在Hitachi S-3500N扫描电镜下观察并照相。
1.2.2 石蜡制片材料经FAA固定、乙醇系列脱水、二甲苯透明、浸蜡并包埋,利用旋转切片机切片,切片厚度8μm,番红-固绿对染,中性胶封片[8]。OLYMPUS BX53光学显微镜(LM)下观察并照相,同时利用CellSens软件测量各项解剖学指标。
1.2.3 表皮微形态观察取经FAA固定后的植物叶片,用50%的乙醇冲洗后,放入25%的次氯酸钠溶液中浸泡1—2h,待叶片发白后,蒸馏水冲洗,在解剖镜下用解剖针剥离上下表皮,水封后制成临时装片。Olympus BX53光学显微镜(LM)观察并照相。
1.3 指标测量 1.3.1 栅栏系数(CP)

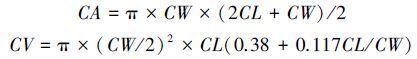
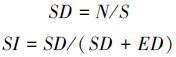
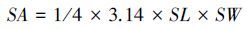
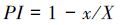
利用Excel(2010版)软件对叶片解剖学指标和气孔参数进行计算,利用SPSS 20.0软件,采用One-way ANOVA法分析不同海拔高度的叶片解剖学特征和气孔参数的差异。采用Pearson相关分析来检验叶片各解剖学特征与海拔的相关性。
2 实验结果 2.1 表皮细胞和气孔的形态特征3 种植物上下表皮角质层纹饰、气孔与表皮细胞的位置关系、气孔外拱盖内缘特征、表皮细胞特征和气孔器特征随海拔升高产生不同的变化(表 2)。
| 形态指标 Morphological indexes | 形态特征Morphological features | |||
| 蒲公英 Taraxacum mongolicum | 火绒草 Leontopodium leontopodioides | 美丽风毛菊 Saussurea superba | ||
| 角质层纹饰Ornamentation of cuticle | 上表皮 | 光滑状 | 条纹状(3600)→光滑状(3800,4400) | 条纹状 |
| 下表皮 | 条纹状(3600,4000)→ 光滑状(4200,4400) | 光滑状(3600,3800)→条纹状(4400) | 条纹状 | |
| 气孔与表皮细胞位置关系 Relative positions between the stomata and epidermal | 上表皮 | 同平面 | 同平面(3600,3800)→气孔下陷(4400) | 同平面 |
| cells | 下表皮 | 同平面 | 同平面 | 同平面 |
| 气孔器外拱盖内缘 Inner margin of the outer stomatal ledge | 上表皮 | 光滑 | 光滑(3600,3800)→浅波状(4400) | 光滑 |
| 下表皮 | 光滑(3600,4000,4200)→浅波状(4400) | 光滑 | 光滑 | |
| 表皮细胞Epidermal cells | 上表皮 | 多边型(3600)→不规则型(4000,4200)→多边型(4400) | 多边型(3600)→不规则型(3800)→多边型(4400) | 不规则型(3600, 3800)→多边型(4000) |
| 下表皮 | 多边型(3600)→不规则型(4000,4200,4400) | 多边型(3600)→不规则型(3800,4400) | 不规则型 | |
| 气孔器Stomatal apparatus | 上表皮 | 无规则型 | 无规则型(3600)→平列型(3800)→无规则型(4400) | 无规则型 |
| 下表皮 | 无规则型 | 无规则型(3600)→平列型(3800)→无规则型(4400) | 无规则型 | |
| →表明指从低海拔到高海拔的变化趋势;3600,3800,4000,4200,4400分别指不同的海拔高度(m) | ||||
蒲公英和美丽风毛菊上下表皮气孔密度SD(图 1)随海拔高度的增加而呈现降低趋势。火绒草上表皮SD随海拔增加呈现明显的增加;其下表皮SD随海拔增加呈现先降低后增加的变化,各海拔高度间差异显著。

|
| 图1 3种植物上、下表皮气孔密度随海拔高度的变化 Fig.1 Changes of stomatal density in upper and lower epidermis of leaves of the three plants with altitude changing |
蒲公英上下表皮气孔指数SI(图 2)变化趋势相反。上表皮除海拔4000 m和4200 m两者之间差异不显著外,其余各海拔间差异显著;下表皮除海拔3600 m和4000 m之间差异不显著外,其余各海拔间差异显著。火绒草上表皮随海拔高度的增加而明显增加,海拔3800 m和4400 m的SI差异不显著;其下表皮SI随海拔的增加先增加后降低的变化,各海拔之间差异显著。美丽风毛菊上表皮SI随海拔的增加先降低后增加的趋势,各海拔高度间差异显著;其下表皮SI与上表皮具有同样的变化趋势,海拔3800 m明显低于其它海拔。

|
| 图2 3种植物上、下表皮气孔指数随海拔高度的变化 Fig.2 Changes of stomatal index in upper and lower epidermis of leaves of the three plants with altitude changing |
蒲公英上、下表皮气孔器面积SA(图 3)随海拔高度的增加而增加,4个海拔之间差异显著。火绒草上、下表皮SA随海拔高度的增加而增加,前者海拔3600 m明显低于其它两个海拔,后者3个海拔间差异显著。美丽风毛菊上、下表皮SA随海拔高度的增加先增加后降低的趋势,各个海拔高度间差异显著。

|
| 图3 3种植物上、下表皮气孔器面积随海拔高度的变化 Fig.3 Changes of stomatal apparatus area in upper and lower epidermis of leaves of the three plants with altitude changing a1、b1、c1/ a2、b2、c2/ a3、b3、c3,分别指蒲公英、火绒草和美丽风毛菊的各个指标随海拔变化有显著性差异(P<0.05) |
3种植物的解剖学数量化指标随海拔高度的变化(表 3—表 5)表明:蒲公英的各项指标均随海拔升高而增大;火绒草除栅栏系数、海绵组织厚度与叶厚之比外,其余各项指标变化趋势同蒲公英;美丽风毛菊除栅栏系数、栅栏细胞系数、上下角质层厚度和上下表皮厚度随海拔升高而增加,其余各项指标均随海拔升高而减小。
| 解剖指标Anatomical indexes | 海拔高度Altitude /m | |||
| 3600 | 4000 | 4200 | 4400 | |
| 叶片厚度Leaf thickness /μm | 238.74±4.42c | 313.59±1.08b | 322.52±2.36b | 413.11±3.80a |
| 上角质层厚度Upper cuticle thickness /μm | 3.19±0.02c | 4.44±0.04b | 5.41±0.05a | 5.34±0.05a |
| 下角质层厚度Lower cuticle thickness /μm | 2.58±0.02d | 3.50±0.04c | 3.83±0.03b | 4.04±0.04a |
| 上表皮厚度Upper epidermis thickness /μm | 27.29±0.18d | 30.85±0.21c | 31.72±0.34b | 50.05±0.39a |
| 下表皮厚度Lower epidermis thickness /μm | 18.50±0.14d | 24.67±0.22c | 28.23±0.15b | 40.74±0.49a |
| 栅栏组织厚度Palisade tissue thickness /μm | 74.45±1.07d | 99.37±0.78c | 105.58±0.75b | 156.98±1.42a |
| 栅栏系数Coefficient of palisade | 0.68±0.02b | 0.66±0.01b | 0.72±0.01b | 1.04±0.04a |
| 栅栏细胞系数Coefficient of palisade cell | 2.33±0.05c | 2.51±0.02b | 2.57±0.03b | 3.29±0.03a |
| 海绵组织厚度Spongy tissue thickness /μm | 112.72±3.72b | 150.76±1.42a | 147.75±2.35a | 155.96±4.57a |
| 栅栏组织厚度与叶厚度比 Palisade tissue thickness/Leaf thickness | 0.31±0.00c | 0.32±0.00bc | 0.33±0.00b | 0.38±0.01a |
| 海绵组织厚度与叶厚度比 Spongy tissue thickness/Leaf thickness | 0.47±0.01ab | 0.48±0.00a | 0.46±0.00b | 0.38±0.01c |
| 叶肉细胞表面积Mesophyll cell area/μm2 | 2806.66±113.54c | 4147.10±48.21b | 3912.44±83.72b | 12969.60±184.75a |
| 叶肉细胞体积Mesophyll cell volume/μm3 | 6785.68±392.85c | 12215.39±202.62b | 11263.60±332.53b | 71189.00±1442.32a |
| 解剖指标Anatomical indexes | 海拔高度Altitude /m | ||
| 3600 | 3800 | 4400 | |
| 叶片厚度Leaf thickness /μm | 205.51±0.55c | 217.64±0.69b | 429.94±2.25a |
| 上角质层厚度Upper cuticle thickness /μm | 2.75±0.03c | 2.97±0.22b | 5.42±0.13a |
| 下角质层厚度Lower cuticle thickness /μm | 1.95±0.04c | 2.30±0.04b | 4.53±0.05a |
| 上表皮厚度Upper epidermis thickness /μm | 29.81±0.24c | 33.47±0.22b | 38.78±0.29a |
| 下表皮厚度Lower epidermis thickness /μm | 20.39±0.24c | 24.18±0.12b | 28.41±0.33a |
| 栅栏组织厚度Palisade tissue thickness /μm | 62.12±0.90c | 73.61±1.93b | 143.63±1.06a |
| 栅栏系数Coefficient of palisade | 0.71±0.02c | 0.94±0.05a | 0.69±0.01b |
| 栅栏细胞系数Coefficient of palisade cell | 2.59±0.04c | 3.05±0.04b | 3.91±0.05a |
| 海绵组织厚度Spongy tissue thickness /μm | 88.49±0.99b | 81.10±1.97c | 209.18±2.68a |
| 栅栏组织厚度与叶厚度比Palisade tissue thickness/Leaf thickness | 0.30±0.00b | 0.34±0.01a | 0.33±0.00a |
| 海绵组织厚度与叶厚度比Spongy tissue thickness/Leaf thickness | 0.43±0.00b | 0.37±0.01c | 0.49±0.00a |
| 叶肉细胞表面积Mesophyll cell area/μm2 | 3196.22±69.29b | 3270.94±57.18b | 6951.77±116.00a |
| 叶肉细胞体积Mesophyll cell volume/μm3 | 8330.56±260.08b | 8878.68±211.84b | 28955.92±686.08a |
| 解剖指标Anatomical indexes | 海拔高度Altitude /m | ||
| 3600 | 3800 | 4000 | |
| 叶片厚度Leaf thickness /μm | 525.90±0.43a | 418.05±1.58b | 358.40±1.08c |
| 上角质层厚度Upper cuticle thickness /μm | 2.20±0.02c | 3.24±0.03b | 3.81±0.04a |
| 下角质层厚度Lower cuticle thickness /μm | 1.77±0.03c | 2.34±0.03b | 2.95±0.02a |
| 上表皮厚度Upper epidermis thickness /μm | 37.71±0.21c | 42.93±0.24b | 51.61±0.24a |
| 下表皮厚度Lower epidermis thickness /μm | 26.28±0.15c | 31.36±0.17b | 32.71±0.13a |
| 栅栏组织厚度Palisade tissue thickness /μm | 190.77±0.74a | 147.30±1.12b | 132.24±0.69c |
| 栅栏系数Coefficient of palisade | 0.71±0.00c | 0.78±0.02b | 0.98±0.01a |
| 栅栏细胞系数Coefficient of palisade cell | 3.34±0.07c | 3.56±0.06b | 4.18±0.07a |
| 海绵组织厚度Spongy tissue thickness /μm | 267.18±0.76a | 190.88±2.25b | 135.08±1.10c |
| 栅栏组织厚度与叶厚度比Palisade tissue thickness/Leaf thickness | 0.36±0.00a | 0.35±0.00b | 0.37±0.00a |
| 海绵组织厚度与叶厚度比Spongy tissue thickness/Leaf thickness | 0.51±0.00a | 0.46±0.00b | 0.38±0.00c |
| 叶肉细胞表面积Mesophyll cell area/μm2 | 9477.20±244.08a | 8425.20±1900.26b | 5774.99±104.69c |
| 叶肉细胞体积Mesophyll cell volume/μm3 | 44787.87±1727.71a | 38146.39±1900.26b | 22275.47±593.71c |
对3种植物叶片的各解剖学指标(叶片总厚度、上下表皮厚度、上下角质层厚度、栅栏厚度、栅栏细胞系数、海绵组织厚度、栅栏系数、叶肉表面积和体积)的Pearson相关分析结果表明:蒲公英(表 6)除了海绵组织厚度与栅栏系数相关性不显著之外,其他解剖指标之间均呈显著相关性(P < 0.01);火绒草(表 7)除了栅栏细胞系数与叶肉表面积和体积相关性不显著之外,其他解剖指标之间均呈显著相关性(P< 0.01);美丽风毛菊(表 8)各个解剖指标之间均呈显著相关性(P< 0.01)。
| 解剖指标 Anatomical indexes | 叶片厚度 Leaf thickness | 上表 皮厚度 Upper epidermis thickness | 下表 皮厚度 Lower epidermis thickness | 上角 质层 厚度 Upper cuticle thickness | 下角 质层 厚度 Lower cuticle thickness | 栅栏组织 厚度 Palisade tissue thickness | 栅栏 细胞 系数 Coefficient of palisade cell | 海绵 组织 厚度 Spongy tissue thickness | 栅栏 系数 Coefficient of palisade | 叶肉细胞 表面积 Mesophyll cell area | 叶肉细胞 体积 Mesophyll cell volume |
| 叶片厚度 Leaf thickness | 1 | 0.891* * | 0.939* * | 0.786* * | 0.850* * | 0.937* * | 0.772* * | 0.761* * | 0.534* * | 0.870* * | 0.847* * |
| 上表皮厚度 Upper epidermis thickness | 1 | 0.933* * | 0.607* * | 0.681* * | 0.949* * | 0.885* * | 0.434* * | 0.770* * | 0.967* * | 0.963* * | |
| 下表皮厚度 Lower epidermis thickness | 1 | 0.766* * | 0.814* * | 0.956* * | 0.861* * | 0.542* * | 0.706* * | 0.911* * | 0.898* * | ||
| 上角质层厚度 Upper cuticle thickness | 1 | 0.917* * | 0.736* * | 0.578* * | 0.604* * | 0.426* * | 0.542* * | 0.511* * | |||
| 下角质层厚度 Lower cuticle thickness | 1 | 0.791* * | 0.664* * | 0.662* * | 0.449* * | 0.618* * | 0.587* * | ||||
| 栅栏组织厚度 Palisade tissue thickness | 1 | 0.848* * | 0.494* * | 0.790* * | 0.938* * | 0.924* * | |||||
| 栅栏细胞系数 Coefficient of palisade cell | 1 | 0.315* * | 0.741* * | 0.834* * | 0.839* * | ||||||
| 海绵组织厚度 Spongy tissue thickness | 1 | -0.132 | 0.415* * | 0.381* * | |||||||
| 栅栏系数 Coefficient of palisade | 1 | 0.762* * | 0.769* * | ||||||||
| 叶肉细胞 表面积 Mesophyll cell area | 1 | 0.998* * | |||||||||
| 叶肉细胞 体积 Mesophyll cell volume | 1 | ||||||||||
| * *指有极显著相关性(P<0.01);*指有显著相关性(P<0.05) | |||||||||||
| 解剖指标 Anatomical indexes | 叶片厚度 Leaf thickness | 上表 皮厚度 Upper epidermis thickness | 下表 皮厚度 Lower epidermis thickness | 上角 质层 厚度 Upper cuticle thickness | 下角 质层 厚度 Lower cuticle thickness | 栅栏组织 厚度 Palisade tissue thickness | 栅栏 细胞 系数 Coefficient of palisade cell | 海绵 组织 厚度 Spongy tissue thickness | 栅栏 系数 Coefficient of palisade | 叶肉细胞 表面积 Mesophyll cell area | 叶肉细胞 体积 Mesophyll cell volume |
| 叶片厚度 Leaf thickness | 0.846* * | 0.517* * | 0.803* * | 0.869* * | 0.902* * | 0.254* * | 0.962* * | -0.448* * | 0.927* * | 0.960* * | |
| 上表皮厚度 Upper epidermis thickness | 1 | 0.677* * | 0.765* * | 0.833* * | 0.857* * | 0.430* * | 0.738* * | -0.185* | 0.731* * | 0.786* * | |
| 下表皮厚度 Lower epidermis thickness | 1 | 0.785* * | 0.784* * | 0.753* * | 0.796* * | 0.298* * | 0.356* * | 0.286* * | 0.407* * | ||
| 上角质层厚度 Upper cuticle thickness | 1 | 0.921* * | 0.913* * | 0.635* * | 0.649* * | 0.055 | 0.594* * | 0.692* * | |||
| 下角质层厚度 Lower cuticle thickness | 1 | 0.956* * | 0.623* * | 0.723* * | -0.012 | 0.675* * | 0.769* * | ||||
| 栅栏组织 厚度Palisade tissue thickness | 1 | 0.566* * | 0.752* * | -0.056 | 0.725* * | 0.808* * | |||||
| 栅栏细胞 系数Coefficient of palisade cell | 1 | 0.023 | 0.632* * | -0.072 | 0.067 | ||||||
| 海绵组织 厚度Spongy tissue thickness | 1 | -0.652* * | 0.963* * | 0.960* * | |||||||
| 栅栏系数 Coefficient of palisade | 1 | -0.667* * | -0.563* * | ||||||||
| 叶肉细胞 表面积Mesophyll cell area | 1 | 0.988* * | |||||||||
| 叶肉细胞 体积Mesophyll cell volume | 1 | ||||||||||
| * *指有极显著相关性(P<0.01);*指有显著相关性(P<0.05). | |||||||||||
| 解剖指标 Anatomical indexes | 叶片厚度 Leaf thickness | 上表 皮厚度 Upper epidermis thickness | 下表 皮厚度 Lower epidermis thickness | 上角 质层 厚度 Upper cuticle thickness | 下角 质层 厚度 Lower cuticle thickness | 栅栏组织 厚度 Palisade tissue thickness | 栅栏 细胞 系数 Coefficient of palisade cell | 海绵 组织 厚度 Spongy tissue thickness | 栅栏 系数 Coefficient of palisade | 叶肉细胞 表面积 Mesophyll cell area | 叶肉细胞 体积 Mesophyll cell volume |
| 叶片厚度 Leaf thickness | 1 | -0.930* * | -0.949* * | -0.964* * | -0.945* * | 0.973* * | -0.613* * | 0.993* * | -0.812* * | 0.881* * | 0.685* * |
| 上表皮厚度 Upper epidermis thickness | 1 | 0.842* * | 0.891* * | 0.941* * | -0.884* * | 0.639* * | -0.946* * | 0.867* * | -0.852* * | -0.445* * | |
| 下表皮厚度 Lower epidermis thickness | 1 | 0.922* * | 0.861* * | -0.929* * | 0.556* * | -0.940* * | 0.742* * | -0.829* * | -0.725* * | ||
| 上角质层厚度 Upper cuticle thickness | 1 | 0.912* * | -0.955* * | 0.587* * | -0.950* * | 0.743* * | -0.854* * | -0.664* * | |||
| 下角质层厚度 Lower cuticle thickness | 1 | -0.902* * | 0.644* * | -0.951* * | 0.833* * | -0.856* * | -0.536* * | ||||
| 栅栏组织 厚度Palisade tissue thickness | 1 | -0.577* * | 0.939* * | -0.664* * | 0.883* * | 0.744* * | |||||
| 栅栏细胞 系数Coefficient of palisade cell | 1 | -0.624* * | 0.587* * | -0.722* * | -0.283* * | ||||||
| 海绵组织 厚度Spongy tissue thickness | 1 | -0.875* * | 0.872* * | 0.626* * | |||||||
| 栅栏系数 Coefficient of palisade | 1 | -0.815* * | -0.282* * | ||||||||
| 叶肉细胞 表面积Mesophyll cell area | 1 | 0.996* * | |||||||||
| 叶肉细胞 体积Mesophyll cell volume | 1 | ||||||||||
| * *指有极显著相关性(P<0.01);*指有显著相关性(P<0.05) | |||||||||||
叶片解剖指标(上表皮厚度、下表皮厚度、上表皮气孔密度及下表皮气孔密度、栅栏厚度、海绵厚度、栅栏细胞系数及叶片厚度)的可塑性分析结果表明3种植物的叶片结构表现出较大的可塑性(表 9)。其中,蒲公英和火绒草的上、下表皮气孔密度可塑性显著高于表皮厚度可塑性;美丽风毛菊的海绵组织厚度可塑性最大。不同植物同一指标相比较,蒲公英的上、下表皮厚度和上表皮气孔密度最大;而3种植物中,火绒草的下表皮气孔密度、栅栏组织厚度、海绵组织厚度、栅栏细胞系数和叶片厚度最大。
| 指标Indexes | 可塑性指数plasticity index | ||
| 蒲公英 Taraxacum mongolicum | 火绒草 Leontopodium leontopodioides | 美丽风毛菊 Saussurea superba | |
| 上表皮厚度Upper epidermis thickness | 0.45 | 0.23 | 0.27 |
| 下表皮厚度Lower epidermis thickness | 0.54 | 0.28 | 0.2 |
| 上表皮气孔密度Upper epidermis stomatal density | 0.73 | 0.54 | 0.18 |
| 下表皮气孔密度Lower epidermis stomatal density | 0.49 | 0.84 | 0.27 |
| 栅栏组织厚度Palisade tissue thickness | 0.53 | 0.57 | 0.31 |
| 海绵组织厚度Spongy tissue thickness | 0.23 | 0.61 | 0.49 |
| 栅栏细胞系数Coefficient of palisade cell | 0.29 | 0.34 | 0.2 |
| 叶片厚度Leaf thickness | 0.42 | 0.52 | 0.32 |
植物及其生长环境是统一的整体,环境的长期作用能够直接影响植物的生态特征及形态解剖结构[13]。叶片作为植物体暴露在环境中面积最大的器官,对环境的变化表现得更为敏感[14]。
3.1 气孔性状的生态适应性在长期的进化过程中,不同和同种植物不同种群的气孔性状差异显著,尤其是对温度、光照、CO2浓度、水分等环境因子的变化很敏感[15]。
本研究中,火绒草上、下表皮气孔密度随海拔的升高而增加,这与安黎哲等[16]的研究结果一致。通常,随着海拔高度的增加,温度、O2和CO2分压会降低,植物可能通过增大气孔密度来加大对O2和CO2的摄入,从而提高光合作用效率,同时改善在较高光强下由于具有较大的叶肉表面积而造成对CO2扩散的阻力[17],这是植物对高山环境中低O2和CO2分压的一种适应。但是,蒲公英和美丽风毛菊的上、下表皮气孔密度却随海拔的增加而降低,Reddy等[18]认为,温度增高可以增加叶片气孔密度,随海拔高度的增加,气温下降,这2种植物很可能通过降低气孔密度来适应高山环境中的低温。这说明不同的植物对CO2浓度、光强、温度等环境因子的敏感性不同,导致上述植物叶片气孔密度随海拔高度增加出现相反的变化趋势。
本文中的3种植物叶片上、下表皮气孔指数随海拔高度的增加均未出现规律性变化。Kürschner[19]的研究表明气孔指数随光照强度的增加而保持相对稳定,但Poole等[20]的研究表明,气孔指数随光照增强而增加。也有研究指出,气孔指数对温度和CO2变化的反应具有稳定性[19, 21]。对有些植物来说,水分的增多会使气孔指数出现稍微的增加或者减少的现象[22],但总体来说,水分对气孔指数的影响不明显[23]。本研究结果说明气孔指数对环境因子的响应和适应表现的更加复杂和多样,因此,要揭示气孔指数随海拔梯度的变化规律,还需进行更深入的研究。
贺金生等[24]对不同海拔高度的高山栎(Quercus semecarpifolia Smith)叶片的形态结构进行了研究,结果表明:随着海拔高度的升高,气孔数目减少、气孔器面积增大。Carpenter和smith[25]认为气孔器面积主要受温度、光照和降水的影响。本研究中,蒲公英和火绒草的上下表皮气孔器面积随海拔高度的升高呈现出增大的趋势,这是植物与高原独特的气候长期适应的结果。
3.2 叶解剖学特征的生态适应性段喜华等[26]对分布于青藏高原东部边缘干旱河谷地区的不同海拔高度阳坡或半阳坡泡沙参(Adenophora potaninii Korsh)叶片进行进行了研究,结果表明:随着海拔的升高,角质层增厚,这与本研究结果相一致。角质层增厚可以有效地防止强光照射,降低叶片的蒸腾作用,使植物具有较强的抗逆性。本研究结果进一步证实了植物对高山环境中的寒冷气候和强辐射的适应。
本研究的3种植物上、下表皮厚度及叶片总厚度均随海拔的升高而增厚。温度通常随海拔的升高而降低,同时空气越来越稀薄,大气的保温作用减弱,不利于热量的积累和保持,较厚的叶片增强了植物的保温作用。蔡永立和宋永昌[27]的研究也表明叶片总厚度和上、下表皮厚度随着海拔的升高而增加是植物对低温环境的一种适应。叶片厚度的增加,一方面增加了叶肉细胞面积和叶面积之比,有利于减少体内水分的过度蒸腾,提高了水分利用效率[6],另一方面叶片厚度的增加也可以防止强辐射对叶肉细胞造成损伤[28]。本研究结果说明随海拔的升高植物叶片增厚是植物对低温和强辐射的一种适应。
本文研究结果显示,蒲公英和火绒草的栅栏组织和海绵组织随海拔升高而增大,而美丽风毛菊则相反;而3种植物栅栏系数和栅栏细胞系数均随海拔升高呈增加趋势。Mendes等[29]的研究指出,发达的栅栏组织是植物对强辐射的一种适应。强辐射和低温同时存在高山地区,发达的栅栏组织也可能是环境中强辐射和低温共同作用的结果。直接而强烈的太阳辐射,使高原植物处于减少叶灼伤和提高光合效率的矛盾之中,较厚的栅栏组织,既可以增加光量子的吸收,提高光合作用效率,又可以减少强光对叶肉的损伤[6, 29]。蔡永立和宋永昌[1]的研究指出,栅栏组织和海绵组织厚度增加都是植物对低温环境的反应,而李全发等[30]指出栅栏组织厚度与植物的保水功能关系密切,海绵组织增加是植物对低温环境的适应。Rengifo[31]等在CO2浓度控制实验中发现栅栏组织、海绵组织厚度及其与叶片厚度的比值与CO2浓度有密切关系。在一定程度上,栅栏系数相对较大的植物,净光合速率较大[32]。
3.3 解剖学特征相关性和可塑性与生态适应性结构特征最能体现环境因子与植物的协同进化[33]。本研究结果显示,虽然3种植物叶片的解剖特征随海拔高度的变化有所不同,但除了蒲公英的栅栏系数与海绵组织厚、火绒草的栅栏细胞系数与叶肉细胞表面积和体积相关性不显著外,其它各指标相互之间均显著相关。由此可知,叶片的解剖学特征之间存在明显的协同进化:较厚的上、下角质层厚度对应较厚的上、下表皮厚度,但不同的植物在不同海拔高度上表现出的协同进化又具有种间差异,说明随着海拔高度的变化,不同的植物采取不同的响应机制来适应高山恶劣环境[5]。同时,解剖学特征的协同进化也是植物对环境变化的一个很重要的适应策略。
植物的表型可塑性与其生态适应能力密切相关[34]。较大的表型可塑性可以使植物在资源受到限制、存在各种胁迫的条件下仍能维持必要的生理功能。本文中3种植物,尤其是火绒草叶片的栅栏组织厚度、海绵组织厚度、栅栏细胞系数和叶片厚度等指标较大的可塑性说明这些指标能够通过改变和调节光能的吸收和利用效率,忍受高山强烈的辐射,是对青藏高原强紫外线环境的一种适应,加之蒲公英和火绒草具有较大的表皮气孔密度可塑性,使这两种植物能够适应较宽范围的生态环境。植物叶片结构上的这种表型可塑性,是它们广泛分布于不同生境的解剖学基础,可以使物种具有更宽的生态幅,从而占据更加广阔的地理空间,成为生态位理论中的广幅种。
| [1] | 卢存福, 简令成, 匡廷云. 高山植物的抗寒抗冻特性. 植物学通报, 1998, 15(3): 17-22. |
| [2] | Willis C G, Ruhfel B, Primack R B, Miller-Rushing A J, Davis C C. Phylogenetic patterns of species loss in Thoreau's woods are driven by climate change. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(44): 17029-17033. |
| [3] | Gedan K B, Bertness M D. Experimental warming causes rapid loss of plant diversity in New England salt marshes. Ecology Letters, 2009, 12(8):842-848. |
| [4] | 李有忠, 贲桂英, 韩发, 师生波. 海拔高度的变化对植物叶片内部结构的影响. 青海师范大学学报: 自然科学版, 1995(4): 34-40. |
| [5] | 马建静, 吉成均, 韩梅, 张婷芳, 闫雪东, 胡东, 曾辉, 贺金生. 青藏高原高寒草地和内蒙古高原温带草地主要双子叶植物叶片解剖特征的比较研究. 中国科学: 生命科学, 2012, 42(2): 158-172. |
| [6] | 施海燕, 王一峰, 王剑虹. 青藏高原不同海拔地7种风毛菊属植物叶片解剖结构的研究. 植物研究, 2009, 29(3): 289-294. |
| [7] | 侯迎, 王乃昂, 张学敏, 程弘毅, 路俊伟. 基于树轮资料重建祁连山东段冷龙岭1848年以来的干湿变化. 山地学报, 2011, 29(1): 12-18. |
| [8] | Pandey S, Kumar N, Kushwaha R. Morpho-anatomical and physiological leaf traits of two alpine herbs,Podophyllum hexandrum and Rheumemodi in the Western Himalaya under different irradiances. Photosynthetica, 2006, 44(1):11-16. |
| [9] | Pyankov V I, Kondratchuk A V, Shipley B. Leaf structure and specific leaf mass: the alpine desert plants of the Eastern Pamirs, Tadjikistan. New Phytologist, 1999, 143(1): 131-142. |
| [10] | Kouwenberg L L R, McElwain J C, Kürschner W M, Wagner F, Beerling D J, Mayle F E, VisscherH. Stomatal frequency adjustment of four conifer species to historical changes in atmospheric CO2. American Journal of Botany, 2003, 90(4): 610-619. |
| [11] | Wise R R, Sassenrath-Cole G F, Percy R G. A comparison of leaf anatomy in field-grownGossypium hirsutum and G. barbadense. Annals of Botany, 2000, 86(4): 731-738. |
| [12] | Gratani L, Covone F, Larcher W. Leaf plasticity in response to light of three evergreen species of the Mediterranean maquis. Trees, 2006, 20(5): 549-558. |
| [13] | 高建平, 王彦涵, 陈道峰. 不同产地华中五味子叶表皮结构和导管分子的解剖学特征及其与环境因子的关系. 西北植物学报, 2003, 23(5): 715-723. |
| [14] | 祁建, 马克明, 张育新. 辽东栎(Quercus liaotungensis) 叶特性沿海拔梯度的变化及其环境解释. 生态学报, 2007, 27(3): 930-937. |
| [15] | 张立荣, 牛海山, 汪诗平, 李英年, 赵新全. 增温与放牧对矮嵩草草甸4种植物气孔密度和气孔长度的影响. 生态学报, 2010, 30(24): 6961-6968. |
| [16] | 安黎哲, 戴怡龄, 陈拓, 徐世健, 张满效, 刘艳红, 程国栋. 乌鲁木齐河源区不同海拔的火绒草叶片结构特征的比较研究. 冰川冻土, 2004, 26(4): 474-480. |
| [17] | Mott K A, Gibson A C, O'leary J W. The adaptive significance of amphistomatic leaves. Plant, Cell and Environment, 1982, 5(6): 455-460. |
| [18] | Reddy K R, Robana R R, Hodges H F, Liu X J, McKinion J M. Interactions ofCO2 enrichment and temperature on cotton growth and leaf characteristics. Environmental and Experimental Botany, 1998, 39(2):117-129. |
| [19] | Kürschner W M. The anatomical diversity of recent and fossil leaves of the durmast oak(Quercus petraea Lieblein/Q. pseudocastanea Goeppert)‐implications for their use as biosensors of palaeoatmospheric CO2 levels. Review of Palaeobotany and Palynology, 1997, 96(1/2): 1-30. |
| [20] | Poole I, Weyers J D B, Lawson T, Raven J A. Variations in stomatal density and index:implications for palaeoclimatic reconstructions. Plant, Cell and Environment, 1996, 19(6):705-712. |
| [21] | Beerling D J,Royer D L. Reading a CO2 signal from fossil stomata. New Phytologist, 2002, 153(3):387-397. |
| [22] | Tichá I. Photosynthetic characteristics during ontogenesis of leaves. 7. Stomata density and sizes. Photosynthetica, 1982, 16(3): 375-471. |
| [23] | Dixon M, Thiec D L E, Garrec J P. The growth and gas exchange response of soil-planted Norway spruce [Picea abies(L.)Karst.] and red oak(Quercus rubra L.) exposed to elevated CO2 and to naturally occurring drought. New Phytologist, 1995, 129(2): 265-273. |
| [24] | 贺金生, 陈伟烈, 王勋陵. 高山栎叶的形态结构及其与生态环境的关系. 植物生态学报, 1994, 18(3): 219-227. |
| [25] | Carpenter S B, Smith N D. Stomatal distribution and size in southern Appalachian hardwoods. Canadian Journal of Botany, 1975, 53(11): 1153-1156. |
| [26] | 段喜华, 孙立夫, 马书荣, 祖元刚. 不同海拔高度泡沙参叶片形态研究. 植物研究, 2003, 23(3): 334-336. |
| [27] | 蔡永立, 宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学 Ⅰ. 叶片解剖特征的比较. 植物生态学报, 2001, 25(1): 90-98. |
| [28] | 李芳兰, 包维楷, 刘俊华, 吴宁. 岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究. 应用生态学报, 2006, 17(1): 5-10. |
| [29] | Mendes M M, Gazarini L C, Rodrigues M L. Acclimation ofMyrtus communis to contrasting Mediterranean light environments-effects on structure and chemical composition of foliage and plant water relations. Environmental and Experimental Botany, 2001, 45(2):165-178. |
| [30] | 李全发, 王宝娟, 安丽华, 吉成均. 青藏高原草地植物叶解剖特征. 生态学报, 2013, 33(7): 2062-2070. |
| [31] | Rengifo E, Urich R, Herrera A. Water relations and leaf anatomy of the tropical species,Jatropha gossypifolia and Alternanthera crucis, grown under an elevated CO2 concentration. Photosynthetica, 2002, 40(3): 397-403. |
| [32] | Martin P J, Stephens W. Willow growth in response to nutrients and moisture on a clay landfill cap soil.Ⅰ. Growth and biomass production. Bioresource Technology, 2006, 97(3): 437-448. |
| [33] | 刘全宏, 王孝安, 田先华, 肖娅萍. 太白红杉(Larix chinensis)叶的形态解剖学特征与环境因子的关系. 西北植物学报, 2001, 21(5): 885-893. |
| [34] | 周世权, 马恩伟. 植物分类学. 北京: 中国林业出版社, 1995. |