 2016, Vol. 36
2016, Vol. 36文章信息
- 武晓东, 袁帅, 付和平, 张晓东, 张福顺, 高全荣
- WU Xiaodong, YUAN Shuai, FU Heping, ZHANG Xiaodong, ZHANG Fushun, GAO Quanrong
- 不同干扰下阿拉善荒漠啮齿动物优势种对气候变化的响应
- Responses of dominant rodentspecies to climate change in different disturbed habitats in the Alashan desert
- 生态学报, 2016, 36(6): 1765-1773
- Acta Ecologica Sinica, 2016, 36(6): 1765-1773
- http://dx.doi.org/10.5846/stxb201408061564
-
文章历史
- 收稿日期: 2014-08-06
- 网络出版日期: 2015-07-29






2. 草业与草地资源教育部重点实验室, 呼和浩特 010019;
3. 内蒙古自治区林业监测规划院, 呼和浩特 010020;
4. 中国农业科学研究院草原研究所, 呼和浩特 010010;
5. 内蒙古医科大学, 呼和浩特 010110
2. Key Laboratory of Prataculture Grassland Resources, Ministry of Education, Hohhot 010019, China;
3. Inner Mongolia Forestry Monitoring and Planning Institute, Hohhot 010020, China;
4. Grassland Research Institute, Chinese Academy of Agricultural Sciences, Hohhot 010010, China;
5. Inner Mongolia Medical University, Hohhot 010110, China
过去几十年的气候变化已对物种分布范围和丰富度产生了极大的影响[1, 2],未来气候变化将使物种分布和丰富度发生更深刻的改变,这种改变将对生物多样性保护带来一定挑战[3]。为了在气候变化下有效保护生物多样性,科学认识气候变化对物种分布的影响将是关键[4, 5]。国际上关于动物对气候变化响应的研究已经广泛展开。这些研究表明,气候变化影响了动物的地理分布、物候、个体行为以及种群动态等[6]。但多数研究是在较大尺度和大陆尺度上展开的[7, 8],同时用模拟的方法预测未来几十年动物的响应[9, 10, 11],而在较小尺度上研究气候变化对动物的影响较少[12, 13, 14]。虽然我国气候变化对啮齿动物分布影响已经开展了一些研究[15, 16],但有关气候变化对荒漠啮齿动物影响的研究还较为少见。
温度和降水是研究气候变化对生物影响的两个常用指标。水热是制约荒漠生态系统的关键因子,环境的小幅波动都会造成这一系统的深刻变化。因此,荒漠生态系统对气候变化的敏感性将会更高。啮齿动物是荒漠生态系统中的重要组成部分和环境变化的重要指示者[17],研究啮齿动物对气候变化的响应,对于反映气候变化下的荒漠生态系统具有重要的意义。从试验地所处荒漠区有气象资料记录来看,2002—2010年平均温度为9.05℃,比1991—2001年平均温度升高了0.41℃。年均降雨量2002—2010为165.36mm,比上一个10a的年均降雨量增长了23.46mm。荒漠啮齿动物,特别是啮齿动物优势种在这样的气候条件下会做出如何响应?这些响应是否会因为人为干扰的存在而发生变化?是本研究关注的重点,现将结果报道如下。
1 研究区自然概况研究区(104°10′— 105°30′E,37°24′— 38°25′N)位于内蒙古阿拉善左旗南部的荒漠景观中,地处腾格里沙漠东缘。气候为典型的大陆性高原气候。年降水量75—215 mm,主要集中在7—9月。年蒸发量3000—4000mm。土壤为灰漠土和灰棕土。草场类型属于温性荒漠类型,植被稀疏,结构单调,以旱生、超旱生和盐生的灌木、小灌木、半灌木和小半灌木为主。建群种以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)植物为主。
在原生生境植被条件一致的情况下,依据该地区对草地利用方式的不同,选择4种不同干扰类型的生境作为取样样区,样区的植被特征如下[18]:
Ⅰ 禁牧区,面积206.6 hm2,在原生植被基础上,自1997年开始围封禁牧,植被以刺叶柄棘豆(Oxytropis aciphylla)和圆头蒿(Artemisia sphaerocephala)为主,其次为短脚锦鸡儿(Caragana brachypoda)、红砂(Reaumuria songarica)和黑沙蒿(Artemisia ordosica)等小灌木,草本以蒙古虫实(Corispermum mongolicum)、沙蓬(Agriophyllum pungens)和糙隐子草(Cleistogenes squarrosa)为主,伴生有雾冰藜(Bassia dasyphylla)等一年生植物,植被盖度较高为23.0%。
Ⅱ 轮牧区,面积173.3 hm2,1995年开始采取围栏轮牧的利用方式,划分为3个区,轮牧50—60只成年羊,每区放牧的时间为1.5个月,3个区轮替放牧。植被以红砂建群,其次为短脚锦鸡儿(Caragana brachypoda)、白刺(Nitraria sphaerocarpa)和霸王(Sarcozygium xanthoxylon)等多年生小灌木,草本植物以糙隐子草、雾冰藜、白草(Pennisetum centrasiaticum)和条叶车前(Plantago lessingii)为主,植被盖度19.7%。
Ⅲ 过牧区,面积146.6 hm2,连续放牧,放牧550—620只羊,荒漠草原理论载畜量每公顷0.625—1.07只羊单位,该区的放牧强度为每公顷3.75—4.23只羊单位。植被以红砂建群,伴生有霸王、白刺、短脚锦鸡儿和驼绒藜(Ceratoides latens)等多年生小灌木,草本以牛心朴子(Cynanchum komarovii)和骆驼蓬(Peganum harmala)等多年生植物为主,伴生有白草和蒙古虫实等一年生植物,植被盖度较低为16.4%。
Ⅳ 开垦区,面积180 hm2,1994年开垦。植被主要以人工种植的梭梭(Haloxylon ammodendron)为主,伴生有短脚锦鸡儿等小灌木,草本以牛尾蒿(Artemisia dubia)、雾冰藜和条叶车前等一年生杂类草为主,植被盖度可达23.5%。
2 研究方法 2.1 数据采集2 002—2010年在上述4种不同干扰类型的生境中分别选取2个固定的标志流放地,面积分别为1 hm2,采用标志重捕法。每年4—10月的月初进行野外调查取样,每个样地布放56个鼠笼,笼距和行距为15 m×15 m,每月连捕4d。为了减少标志对鼠类正常活动的影响,跳鼠采用带编号的金属环进行标记,其余鼠种用剪趾法标记。记录所捕个体的种名、性别、体重(g)、繁殖状况及捕获位置,标志后原地释放[19]。
气象数据均来自“阿拉善盟孪井滩气象站”,该站距离研究样地平均距离5.5km。气象资料包括1991—2010年的年平均温度(℃)及年降水量(mm)。
2.2 数据分析啮齿动物种群相对数量使用百笼捕获率,计算公式如下:

使用Spearman相关分析啮齿动物优势种群相对数量与年平均温度和降水量的相关性。
Spearman相关系数:
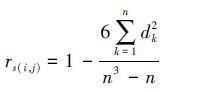
使用典范对应分析(Canonical Correspondence Analysis,CCA)进一步分析环境因子变量组(年平均温度、降水、灌木生物量和草本生物量)与啮齿动物优势种群数量之间是否存在非线性关系。Spearman相关系数分析采用SAS9.0分析软件。CCA采用CANOCO软件分析。显著性水平为P=0.05。
3 研究结果2002— 2010年在研究区共捕获啮齿动物3科,8属,9种。其中跳鼠科(Dipodidae) 3种:三趾跳鼠(Dipus sagitta) 、蒙古羽尾跳鼠(Stylodipus andrewsi)、五趾跳鼠(Allactaga sibirica); 仓鼠科(Cricetidae)5种: 小毛足鼠(Phodopus roborovskii) 、黑线仓鼠(Cricetulus barabensis) 、短耳仓鼠(Allocricetulus eversmanni) 、子午沙鼠(Meriones meridianus) 、长爪沙鼠(Meriones unguiculatus); 松鼠科(Sciuridae)1种: 阿拉善黄鼠(Spermophilus alaschanicus) 。禁牧区优势鼠种为子午沙鼠、次优势种为三趾跳鼠;轮牧区中的优势种均为三趾跳鼠,次优势种为子午沙鼠;过牧区中优势种为五趾跳鼠,次优势种为三趾跳鼠和子午沙鼠;开垦区优势鼠种为子午沙鼠,次优势种为黑线仓鼠(表 1)。
| 鼠种 Rodent species | 捕获量Catch/% | |||
| 禁牧区 Grazing exclusion | 轮牧区 Rotational grazing | 过牧区 Over grazing | 开垦区 Cultivation | |
| 五趾跳鼠Allactaga sibirica | 6.33 | 13.35 | 35.53 | 11.11 |
| 三趾跳鼠Dipus sagitta | 32.18 | 35.96 | 22.76 | 0.00 |
| 蒙古羽尾跳鼠Stylodipus andrewsi | 0.00 | 0.00 | 0.12 | 0.00 |
| 小毛足鼠Phodopus roborovskii | 15.17 | 12.11 | 6.72 | 1.97 |
| 黑线仓鼠Cricetulus barabensis | 5.03 | 0.88 | 1.27 | 19.64 |
| 短耳仓鼠Allocricetulus eversmanni | 0.00 | 0.19 | 0.12 | 0.00 |
| 子午沙鼠Meriones meridianus | 35.56 | 33.98 | 22.66 | 56.81 |
| 长爪沙鼠Meriones unguiculatus | 0.04 | 0.84 | 2.61 | 4.07 |
| 阿拉善黄鼠Spermophilus alaschanicus | 5.69 | 2.69 | 8.22 | 6.40 |
2 002—2010年各区啮齿动物捕获率与温度和降水变化趋势如图 1和图 2。由图 1和图 2可知,2003年和2008年是研究区啮齿动物数量的两个高峰年,4个样区的平均捕获率分别为15.90%和15.18%,分别是2002—2010年平均捕获率(9.70%)的1.64倍和1.56倍,与之相对应的年平均温度是两个较低的年份,分别为8.8℃和8.3℃,分别比2002—2010年平均温度(9.06℃)低0.26 ℃和0.76℃,降水量则为两个较高的年份,分别为202.6mm和175.2mm,比2002—2010年平均降水量(165.37mm)分别高出37.23mm和9.83mm。

|
| 图1 不同生境中啮齿动物丰富度与年均温度变化 Fig.1 Annual mean temperature and rodent abundance in different habitats from 2002 to 2010 |

|
| 图2 不同生境中啮齿动物丰富度与年降水量变化 Fig.2 Annual mean precipitation and rodent abundance in different disturbance habitats |
为了分析各样区啮齿动物种群数量与年平均温度和降水量的相关性,选择捕获率相对较高的三趾跳鼠、五趾跳鼠、子午沙鼠和黑线仓鼠,采用Spearman相关进行分析(表 2)。从温度与啮齿动物种群数量间关系的分析结果可以看出,禁牧区中的优势种子午沙鼠和常见种黑线仓鼠的种群数量与年平均温度具有显著的负相关关系(P<0.05);轮牧区中的次优势种子午沙鼠和常见种黑线仓鼠种群数量与年均温度显著负相关(P<0.05,P<0.01);过牧区中的次优势种子午沙鼠和开垦区中的优势种子午沙鼠种群数量均与年平均温度存在显著的负相关关系(P<0.01,P<0.05)。从降雨与啮齿动物种群数量间关系的分析结果可以看出,除开垦区外,其他干扰区内的子午沙鼠种群数量均与上年的年均降雨量存在显著正相关关系(P<0.05)。轮牧区和开垦区的子午沙鼠种群数量与当年降雨量显著正相关(P<0.05)。开垦区内的次优势种黑线仓鼠种群数量与年平均降雨量呈显著正相关关系(P<0.05)。除过牧区内的三趾跳鼠与上年平均降雨量有显著正相关关系外(P<0.05),其余干扰区内的跳鼠均为未与年平均温度和降雨量产生显著的线性关系。
| 处理 Treatments | 鼠种 Rodent species | 平均温度 Annual mean temperature | 上年年降雨量 Annual precipitation in last year | 当年年降雨量 Annual precipitation in this year |
| 禁牧 | 三趾跳鼠Dipus sagitta | 0.00664 | 0.01768 | 0.27205 |
| Prohibited-grazing | 五趾跳鼠Allactaga sibirica | -0.30838 | 0.16630 | 0.03491 |
| 子午沙鼠Meriones meridianus | -0.66152* | 0.70940* | 0.28766 | |
| 黑线仓鼠Cricetulus barabensis | -0.53362* | 0.34187 | 0.21142 | |
| 轮牧 | 三趾跳鼠Dipus sagitta | -0.11074 | 0.28222 | 0.43307 |
| Rotational-grazing | 五趾跳鼠Allactaga sibirica | 0.01780 | 0.03761 | 0.28476 |
| 子午沙鼠Meriones meridianus | -0.73977* * | 0.63054* | 0.54246* | |
| 黑线仓鼠Cricetulus barabensis | -0.52190* | 0.37574 | 0.30638 | |
| 过牧 | 三趾跳鼠Dipus sagitta | -0.49945 | 0.58518* | 0.15738 |
| Over-grazing | 五趾跳鼠Allactaga sibirica | 0.04430 | 0.10177 | 0.08929 |
| 子午沙鼠Meriones meridianus | -0.72566* * | 0.64878* | 0.22799 | |
| 黑线仓鼠Cricetulus barabensis | -0.37338 | 0.37574 | 0.26720 | |
| 开垦 | 三趾跳鼠Dipus sagitta | — | — | — |
| Cultivation | 五趾跳鼠Allactaga sibirica | 0.02836 | -0.08757 | -0.40541 |
| 子午沙鼠Meriones meridianus | -0.65488* | 0.30719 | 0.58200* | |
| 黑线仓鼠Cricetulus barabensis | -0.29399 | -0.20912 | 0.64424* | |
| —未捕获啮齿动物,*P<0.05,** P<0.01 | ||||
为了进一步分析环境因子变量组与群落物种组成变量组之间的相关性,本文对年平均温度、降水、和植物生物量与啮齿动物优势种种群数量之间的关系进行CCA分析(表 3,图 3)。表 3的结果表明,所有生境排序轴1的特征值均大于其它排序轴,禁牧区前两个排序轴对物种数据的解释量最高,达到了61.8%;轮牧区前两个排序轴对物种数据的解释量最低,仅为32.7%。4种生境中前两个排序轴均解释了89%以上啮齿动物与环境变量的关系。禁牧、过牧、轮牧和开垦生境中前两个物种排序轴近似垂直,相关系数分别为:0.0232,0.0726,0.0463和-0.2712;前两个环境排序轴的相关系数为0,表明排序结果可信,能够较好的反应啮齿动物种群数量与环境因子之间的关系。
| 生境 Habitats | 禁牧 Grazing exclusion | 过牧 Over grazing | 轮牧区 Rotational grazing | 开垦区 Cultivation | ||||
| 排序轴 Axis | 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 |
| 特征值Eigenvalues | 0.147 | 0.010 | 0.101 | 0.005 | 0.057 | 0.004 | 0.183 | 0.023 |
| 物种与环境间的相关系数 Species-environment correlations | 0.843 | 0.566 | 0.778 | 0.509 | 0.658 | 0.556 | 0.792 | 0.551 |
| 物种数据变化的累积比例 Cumulative percentage variance of species data | 57.900 | 61.800 | 50.700 | 53.000 | 32.700 | 34.800 | 50.100 | 56.200 |
| 物种与环境关系变化累积比例 Cumulative percentage variance of species-environment relation | 92.800 | 99.000 | 95.400 | 99.600 | 90.400 | 96.100 | 89.000 | 100.000 |
| 特征值总和Sum of all eigenvalues | 0.254 | 0.199 | 0.176 | 0.366 | ||||
| 典范特征值总和 Sum of all canonical eigenvalues | 0.159 | 0.106 | 0.064 | 0.206 | ||||
从图 3可以看出,年均温度和上一年的降雨量对禁牧区啮齿动物种群的数量分布有较大影响。降雨对灌木和草本的生物量存在正相关关系。子午沙鼠距离中心点较近,表明禁牧区子午沙鼠受各环境变量影响均较大。三趾跳鼠与草本生物量的反向延长线距离较近,表明三趾跳鼠与草本生物量关系密切。根据啮齿动物物种在各环境变量线上的投影点的距离,可以看出黑线仓鼠和子午沙鼠对较高温度的最适值要低于跳鼠,而对较多降雨的最适值要高于跳鼠;啮齿动物对灌木生物量较高时的最适值与降雨一致,均为黑线仓鼠>子午沙鼠>三趾跳鼠>五趾跳鼠。

|
| 图3 降水量、温度与不同生境中啮齿动物种群数量的CCA排序图 Fig.3 Two-dimensional CCA ordination diagram for weather variables and rodent population abundance P:上年年降水量,T:年平均温度,S:灌木生物量,G:草本生物量,Dipus sagitta:三趾跳鼠、Allactaga sibirica:五趾跳鼠、Meriones meridianus:子午沙鼠、Cricetulus barabensis:黑线仓鼠的捕获率 |
禁牧区中草本生物量与轴1显著正相关(0.61),上年降雨量与轴2显著负相关(-0.52);过牧区年均温度与排序轴1显著负相关(-0.55),草本生物量与排序轴1极显著正相关(0.75);开垦区中的灌木生物量与排序轴1显著负相关(-0.61,表 4)。
| 生境 Habitats 排序轴 Axis | 禁牧 Grazing exclusion | 过牧 Over grazing | 轮牧区 Rotational grazing | 开垦区 Cultivation | ||||
| 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 | 排序轴1 Axis 1 | 排序轴2 Axis 2 | |
| 年均温度 Annual mean temperature | -0.3763 | 0.4132 | -0.5479* | 0.3548 | 0.4414 | 0.1805 | 0.1576 | -0.033 |
| 上年降雨量 Annual precipitation in last year | 0.2245 | -0.5164* | 0.488 | -0.2109 | -0.3093 | 0.0961 | -0.1914 | 0.276 |
| 灌木年生物量 Annual shrub biomass | 0.1506 | -0.2971 | 0.3782 | -0.0723 | -0.0949 | 0.2336 | -0.6055* | 0.1286 |
| 草本年生物量 Annual grass biomass | 0.6051* | -0.1749 | 0.7524* * * | -0.091 | -0.4278 | 0.2706 | -0.2358 | -0.1478 |
| *P <0.05,* * P<0.01,* * * P<0.001 | ||||||||
从图 3可以看出,轮牧区的啮齿动物种群的数量分布受草本生物量和年均温度的影响较大。轮牧区中的灌木和草本与降雨量存在显著的正相关关系,与温度呈负相关。五趾跳鼠种群数量与年平均温度关系密切;子午沙鼠与上年降雨量关系密切;三趾跳鼠与上年降雨量和草本生物量关系密切。根据啮齿动物物种在各环境变量线上的投影点距离,可以看出轮牧区啮齿动物在较高的上年降雨量、灌木生物量和草本生物量时均表现出相同的最适值顺序,即黑线仓鼠>子午沙鼠>三趾跳鼠>五趾跳鼠。但在年均温较高时,跳鼠的最适值要高于子午沙鼠和黑线仓鼠。
从图 3可以看出过牧区的啮齿动物种群数量分布受年均温度、草本生物量和上年降雨量的影响较大。草本和灌木生物量与降雨量呈显著正相关。过牧区的子午沙鼠数量与植物的生物量关系密切;五趾跳鼠和三趾跳鼠距离中心点较近,表明两种跳鼠的数量分布受环境变量的影响较大。过牧区啮齿动物在较高的上年降雨量、灌木生物量和草本生物量时同样均表现出相同的最适值顺序,即黑线仓鼠>子午沙鼠>三趾跳鼠>五趾跳鼠。但在年均温较高时,跳鼠的最适值要高于其他两个鼠种。
从图 3可以看出,开垦区的啮齿动物种群数量分布受灌木生物量和上年降雨量影响较大。降雨对开垦区灌木生物量有正相关作用。子午沙鼠数量与上年降雨量关系密切;黑线仓鼠与草本生物量关系密切;而五趾跳鼠与年均温度和灌木生物量关系密切。黑线仓鼠和子午沙鼠对较高草本生物量、灌木生物量以及上年降雨量的最适值要高于五趾跳鼠;而对较高年均温度的最适值则低于五趾跳鼠。
通过以上分析表明单因素相关分析和环境变量组(温度和降水)与各区啮齿动物群落物种变量组分析均表明啮齿动物与温度和降雨量关系密切,各区优势种总体表现出与温度的负相关关系,和与降雨量的正相关关系。只是在不同的干扰区内不同物种与温度和降水的相关性密切程度不一致。从CCA的结果来看,虽然各区内优势种与环境变量的密切性不同,但并没有改变啮齿动物物种对气候因子的适应性,即跳鼠对温度的最适值要高于其他鼠种,而仓鼠科的啮齿动物对降雨的适应性高于跳鼠。
4 结论与讨论(1)气候变化与啮齿动物
有关气候变化对动物的影响国内外已经积累了很多研究[1, 15, 16, 20]。这些研究多数考察了温度和降雨对啮齿动物的影响。Kausrud等发现温度和湿度强烈影响高山林地鼠类(旅鼠)的动态[13]。Myers等研究了美洲北大湖区域气候变化对9种啮齿动物分布的影响发现气温升高和降雨增加,使得两种鼠类分布区扩大,其啮齿动物集群数量上的主导种由北方种类向南方种类变化[1]。Deitleff等研究20a白足鼠(Peromyocus Leucopus)和草原田鼠(Microtus pennsylvanicus)的丰富度与气候变化的关系发现草原田鼠与降雨正相关[20]。温度变化主要通过对物种个体生理活动和性别发育的影响而对物种产生影响,降水则主要通过对物种繁殖过程和生理活动的影响而对物种产生直接影响。对许多物种来说,气候还主要通过对食物和栖息地的影响而对其产生间接影响[21]。本文的研究表明啮齿动物优势种对温度和降雨的响应不同,尤其以子午沙鼠表现最为显著。虽然子午沙鼠是荒漠区优势鼠种之一,但是温度升高反而会抑制其种群数量,相同研究区的野外观察也发现子午沙鼠对高温的耐受性要低于跳鼠,这与其自身的生物学特性有关。CCA的结果表明降雨促进了植物生物量的增加,而子午沙鼠与植物生物量关系密切,表明降雨对子午沙鼠种群数量存在脉冲效应[22],Spearman相关分析的结果中子午沙鼠对上年降雨量的响应要高于对当年降雨量的响应,也为这种脉冲效应的存在提供了间接证据。
(2)气候干扰与人为干扰
一般认为气候变化是发生在大尺度上的干扰事件,这一干扰在较大尺度上影响着动物分布和丰盛度[7, 8]。而在较小尺度上发生的人为干扰事件可能会加剧或缓冲气候变化对动物的影响。本研究表明虽然不同人为干扰生境中啮齿动物群落的优势种不尽相同,但这些优势种对降雨和温度的最适值在各生境之间均表现出相似的顺序。即跳鼠对温度的适宜性要高于仓鼠科的子午沙鼠和黑线仓鼠,这主要是由于跳鼠本身的生物学和生态学特性决定的,如具有较大的体型和冬眠行为等。而仓鼠科啮齿动物对降雨的适宜性高于跳鼠,从食性来看黑线仓鼠属于食谷类啮齿动物,子午沙鼠虽属于杂食类啮齿动物,但是食种子的比例要高于跳鼠[23, 24],所以荒漠区更高的降雨意味着更多种子产量,从而有利于仓鼠科啮齿动物数量的增长;从生境角度来看,仓鼠科啮齿动物更喜欢栖息于植物盖度大的生境,而跳鼠则喜栖于开阔生境[24, 25],所以较多的降雨也有利于子午沙鼠和黑线仓鼠。因此本研究认为气候变化对荒漠啮齿动物优势种的影响,从温度上,是通过影响啮齿动物的生物学和生态学特性的基础上进行的;而降雨短期内是从影响啮齿动物食物和生境的途径上实现的,这与许多学者认为降雨的作用是一种底-顶效应的观点是一致的[16, 26]。目前在较小尺度上研究气候变化对动物的影响较少[12, 13, 14, 27]。本研究认为较小尺度上的人为干扰更可能从改变食性和生境的途径上加剧或缓冲降雨对荒漠啮齿动物优势种的影响,而不是改变温度对啮齿动物的作用。这种人为干扰将使降雨对荒漠啮齿动物的底-顶效应在小尺度上的表现变得更为复杂,进而可能对荒漠啮齿动物群落构成产生影响。
致谢: 内蒙古农业大学研究生鄂晋、吉晟男、查木哈、韩艳静等在野外数据收集过程中付出了辛勤的劳动;野外工作同时也得到了内蒙古阿拉善盟草原站的大力帮助,在此一并致以衷心的感谢。
| [1] | Myers P, Lundrigan B, Hoffman S M G, Haraminac A P, Seto S H. Climate-induced changes in the small mammal communities of the Northern Great Lakes Region. Global Change Biology, 2009, 15(6): 1434-1454. |
| [2] | Jarema S I, Samson J, Mcgill B J, Humphries M M. Variation in abundance across a species' range predicts climate change responses in the range interior will exceed those at the edge: a case study with North American beaver. Global Change Biology, 2009, 15(2): 508-522. |
| [3] | Miller-Rushing A J, Lloyd-Evans T L, Primack R B, Satzinger P. Bird migration times, climate change, and changing population sizes. Global Change Biology, 2008, 14(9): 1959-1972. |
| [4] | Ahola M P, Laaksonen T, Eeva T, Lehikoinen E. Climate change can alter competitive relationships between resident and migratory birds. Journal of Animal Ecology, 2007, 76(6): 1045-1052. |
| [5] | Hill J K, Fox R. Climate change and British butterfly distributions. Biologist, 2003, 50(3): 106-110. |
| [6] | 马瑞俊, 蒋志刚. 全球气候变化对野生动物的影响. 生态学报, 2005, 25(11): 3061-3066. |
| [7] | 李湘涛. 气候变暖对野生动物的影响. 大自然, 2010, (4): 8-11. |
| [8] | 彭少麟, 李勤奋, 任海. 全球气候变化对野生动物的影响. 生态学报, 2002, 22(7): 1153-1159. |
| [9] | Midgley G F, Hughes G O, Thuiller W, Rebelo A G. Migration rate limitations on climate change-induced range shifts in Cape Proteaceae. Diversity and Distributions, 2006, 12(5): 555-562. |
| [10] | Rebelo H, Tarroso P, Jones G. Predicted impact of climate change on European bats in relation to their biogeographic patterns. Global Change Biology, 2010, 16(2): 561-576. |
| [11] | Loarie S R, Duffy P B, Hamilton H, Asner G P, Field C B, Ackerly D D. The velocity of climate change. Nature, 2009, 462(7276): 1052-1055. |
| [12] | 石建斌, Dunba R, 李迪强, 肖文发. 气候与白昼长度变化对苏格兰拉姆岛上野化山羊种群日活动节律的影响. 动物学研究, 2006, 27(6): 561-568. |
| [13] | Kausrud K L, Mysterud A, Steen H, Vik J O, Østbye E, Cazelles B, Framstad E, Eikeset A M, Mysterud I, Solhøy T, Stenseth N C. Linking climate change to lemming cycles. Nature, 2008, 456(7218): 93-97. |
| [14] | Garroway C J, Bowman J, Cascaden T J, Holloway G L, Mahan C G, Malcolm J R, Steele M A, Turner G, Wilson P J. Climate change induced hybridization in flying squirrels. Global Change Biology, 2010, 16(1): 113-121. |
| [15] | Jiang G S, Liu J, Xu L, Yan C, He H L, Zhang Z B. Intra- and interspecific interactions and environmental factors determine spatial-temporal species assemblages of rodents in arid grasslands. Landscape Ecology, 2014, doi: 10.1007/s10980-014-0039-6. |
| [16] | Jiang G, Zhao T, Liu J, Xu L, Yu G, He H, Krebs C J, Zhang Z. Effects of ENSO-linked climate and vegetation on population dynamics of sympatric rodent species in semiarid grasslands of Inner Mongolia, China. Canadian Journal of Zoology, 2011, 89(8): 678-691. |
| [17] | Dale V H, Beyeler S C. Challenges in the development and use of ecological indicators. Ecological Indicators, 2001, 1(1): 3-10. |
| [18] | 张晓东, 武晓东, 付和平, 袁帅, 查木哈. 荒漠破碎化生境中长爪沙鼠集合种群野外验证研究. 动物学杂志, 2013, 48(6): 834-843. |
| [19] | 武晓东, 付和平. 人为干扰下荒漠啮齿动物群落格局——变动趋势与敏感性反应. 生态学报, 2006, 26(3): 849-861. |
| [20] | Deitloff J, Falcy M R, Krenz J D, McMillan B R. Correlating small mammal abundance to climatic variation over twenty years. Journal of Mammalogy, 2010, 91(1): 193-199. |
| [21] | 吴建国. 气候变化对陆地生物多样性影响研究的若干进展. 中国工程科学, 2008, 10(7): 60-67. |
| [22] | Orland M C, Kelt D A. Responses of a heteromyid rodent community to large- and small-scale resource pulses: diversity, abundance, and home-range dynamics. Journal of Mammalogy, 2007, 88(5): 1280-1287. |
| [23] | 周延林, 王利民, 鲍伟东. 几种啮齿动物食性比较研究. 内蒙古大学学报: 自然科学版, 2000, 31(6): 613-617. |
| [24] | 娜日苏, 苏和, 武晓东. 五趾跳鼠的植物性食物选择与其栖息地植被的关系. 草地学报, 2009, 17(3): 383-388. |
| [25] | 赵肯堂. 三趾跳鼠(Dipus sagitta Pallas)的生态研究. 动物学杂志, 1964, (2): 59-62. |
| [26] | Brown J H, Ernest S K M. Rain and rodents: complex dynamics of desert consumers. Bioscience, 2002, 52(11): 979-987. |
| [27] | Wilson R J, Gutiérrez D, Gutiérrez J, Monserrat V J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Global Change Biology, 2007, 13(9): 1873-1887. |