 2016, Vol. 36
2016, Vol. 36文章信息
- 孔冬冬, 张强, 顾西辉, 王月, 李华贞
- KONG Dongdong, ZHANG Qiang, GU Xihui, WANG Yue, LI Huazhen.
- 植被对不同时间尺度干旱事件的响应特征及成因分析
- Vegetation responses to drought at different time scales in China
- 生态学报[J]. 2016, 36(24): 7908-7918
- Acta Ecologica Sinica[J]. 2016, 36(24): 7908-7918
- http://dx.doi.org/10.5846/stxb201506111182
-
文章历史
- 收稿日期: 2015-06-11
- 网络出版日期: 2016-04-13

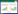




2. 中山大学华南地区水循环与水安全广东省普通高校重点实验室, 广州 510275;
3. 中山大学广东省城市化与地理环境空间模拟重点实验室, 广州 510275
2. Key Laboratory of Water Cycle and Water Security in Southern China of Guangdong High Education Institute, Sun Yat-sen University, Guangzhou 510275, China;
3. Guangdong Provincial Key Laboratory of Urbanization and Geo-simulation, School of Geography and Planning, Sun Yat-sen University, Guangzhou 510275, China
干旱通常是指可利用水分在一连续时段内显著低于正常值,且水分供应不能满足当前需求的自然现象[1]。在全球变暖影响下,干旱发生的频率和强度都有明显增加,严重干旱和极端干旱灾害自20世纪90年代后期变得愈益严重[2],干旱对生态系统的影响越来越突出。生态系统对干旱事件的响应是近年来的研究热点。尽管不同陆地生态系统对干旱的敏感性不同[3],但干旱在不同陆地生态系统种无疑是驱动地上净初级生产力(ANPP)减少的主要因素之一[4],ANPP减少可能会导致地球上有些地区从碳汇变成碳源。如果气候变化诱导的干旱持续增强,将会通过“碳-气候”正反馈调节而加速全球变暖[5-6],从而产生更加严重的干旱,并且可能会导致生态系统不可逆的退化[5]。干旱同时也影响生态系统中的植被生长,不同植被在应对干旱时的不同生理反应决定了对水亏损的抵抗力稳定性和恢复力稳定性的水平[1]。最终影响干旱损害的类型,如减缓生长、减少绿色、损失单位面积生物量、或者植被死亡[1]。干旱对生态系统具有较大的负面影响,彻底理解生态系统,尤其是植被对干旱的响应,变得尤为重要。研究植被对不同时间尺度干旱的响应,以及不同气象因素对植被干旱响应的影响对于提高全球气候变化情景下植被转移预测精度具有重要理论意义[1, 7],同时,对于理解植被对干旱的响应机制,对提高气候波动和气候变化对植被造成的脆弱性的认知具有重要科学价值[1]。
Vicente-Serrano等利用SPEI指标量化干旱、GIMMS-NDVI、ANPP、树木径向生长量化植被活动,在全球范围内研究植被对干旱的响应。但是Vicente-Serrano仅考虑水平衡对植被干旱响应的影响。通过降水和潜在蒸发量化的水平衡,被证明是影响植被响应的地理分布的主要因素[8]。同时很多研究指出NDVI与降水具有很强的相关性[9],虽然植被胁迫也可能受其他因素的影响,如洪水、病虫害、养分、放牧、失火和人类活动等,但它们的相关性主要受区域降水机制、土壤类型、植被类型的影响[9]。其次,有效积温、光照时数也会明显的影响植被活动。此外植被对干旱响应的研究多集中在热带森林[10]和一些国家的局部地区[6],中国区域植被对干旱的响应研究尚未开展。国内很多学者针对局部区域植被对干旱的响应进行研究,于泉洲等利用NDVI和MNDWI研究了南四湖湿地干旱响应特征[11];叶春等对鄱阳湖湿地植被的干旱响应及影响因素进行了研究分析[12];牟成香对青藏高原高寒草甸植物开花物候对极端干旱的响应进行了探讨[13];由于不同生态系统对干旱的敏感性不同,不同植被对干旱的抵抗力稳定和恢复力稳定性存在差异,因此更加细致的研究需要在分区、分植被类型进行研究。但是国内大多数研究集中在某一时间、或某一区域的干旱事件,我国不同区域、不同植被类型对干旱的响应机制依然不明朗。
本文利用GIMMS-NDVI数据与SPEI指标,研究中国不同区域、不同植被类型对干旱的响应机制。本文研究目的:通过归一化植被指数NDVI与SPEI干旱指标的最大相关性,研究干旱对植被活动的影响;通过NDVI指标与SPEI指标最大相关系数对应的干旱时间尺度,研究植被对干旱的敏感性和响应快慢程度等;本文选择4种气象要素:水平衡、降水、有效积温、光照时数,来进一步研究气象要素对植被干旱响应的影响。本文研究将为气候变化对中国生态系统的影响研究提供重要理论依据。
1 数据 1.1 气象数据气象数据来源于中国气象数据共享服务中心(http://cdc.nmic.cn/)提供的地面气候资料日值数据集824个气象站点,数据已由中国气象共享服务中心进行质量检测与控制。气象数据包含逐日降水、最高温度、最低温度、风速、光照时数、相对湿度、气压等要素。提取1980—2006年的气象数据。数据缺失大于1年的站点直接剔除,余下的693个气象站点分布如图 1所示。挑出的693个站点数据缺测极少,大多数站点缺测小于0.5%,最高不超过0.9%。对于缺失值只有1—2d的余下站点,采用该站点相邻日期的数据进行插值;其余缺失采用历史同时期的多年均值来替代[14-15]。

|
| 图 1 气象站点分布示意图 Fig. 1 Locations of the meteorological stations considered in this study |
为了便于NDVI与SPEI的相关性分析,利用站点气象资料计算的SPEI指标采用反距离权重插值为栅格数据,栅格数据精度与NDVI数据相同。
1.2 指标归一化指标(NDVI)本文所分析的归一化植被指数主要依据全球监测与模型组(Global Inventor Modeling and Mapping Studies,GIMMS)利用NOAA系列卫星(NOAA 7,9,11,14和16)合成分辨率为8km的15d最大值NDVI数据集(截取时间为1982年1月至2006年12月),数据经过几何粗较正、辐射较正和大气校正等预处理,再进一步对每日、每轨图像进行几何精校正、除云、除坏线等处理。
为避免不同来源的白噪点,GIMMS-NDVI半月数据融合到逐月时,选上半月与下半月中的最大值作为逐月值[1]。同时为减轻运算复杂程度,使用bilinear插值把NDVI指数栅格大小重采样至0.1°×0.1°。一般认为生长季节NDVI大于0.1的区域才有植被覆盖,NDVI增加表示绿色植被的增加;0.1以下表示地表无植被覆盖,如建设用地、裸土、沙漠、戈壁、水体、冰雪和云等[16](图 2)。

|
| 图 2 1982—2006年生长季多年平均GIMMS-NDVI分布 Fig. 2 The average GIMMS-NDVI during 1982—2006 白色表示生长季多年平均NDVI<0.1的区域为无植被覆盖区 |
土地利用数据来源于欧盟联合研究中心(JRC)空间应用研究所(SAI)开发的GLC2000数据集(http://bioval.jrc.ec.europa.eu/products/glc2000/products.php)[17-18],该数据利用2000年的1km空间分辨率的SPOT-4 VGT S10数据与DEM、积温和降水等通过AHP 法合成[17]。为保证NDVI与土地利用数据间的一致性,土地利用数据用生长季多年平均大于0.1的NDVI做掩膜处理。本研究将土地利用分为5种类型:森林、耕地、草地、灌木、其他(图 3)。

|
| 图 3 土地利用类型数据分布 Fig. 3 Spatial patterns of land use types across China |
Vicente-Serrano等[19]提出了标准化降水蒸散指数(Standardized Precipitation Evapotranspiration Index,简称SPEI)。SPEI的计算是基于降水与温度资料,并且具有多尺度的特点,其最核心的概念是构造一个气象水循环去描述累积的水分缺失或剩余,从而描述干旱。与标准化降水指数指标相比,SPEI指标加入了潜在蒸散发,综合考虑了温度对干旱的影响,能够更准确的刻画干旱,在气候显著升温的区域更能反应真实的干旱情况[15];与帕默尔指标相比,SPEI指标计算简单灵活,且具有多时间尺度特征,适用性广[20]。具体计算方法参见文献[21]。
2.2 模糊聚类分析由于我国东西南北跨度较大、地形复杂以及下垫面特征多样,气象环境以及植被情况复杂。本文先采用模糊聚类分析对中国进行分区,然后再在各均一性分区研究植被对干旱的响应机制。Ruspini[21]提出模糊聚类的概念,Dunn[22]进一步发展此方法。1981年,Bezdek[23]在Dunn[22]的基础上将FCM算法进行推广。聚类分析是根据数据相似性,将其归到不同集合中的方法。而模糊聚类分析则是建立在模糊理论之上,超越 “非此即彼”的传统硬聚类分析,更贴近实际数据内在结构。最常用模糊聚类为模糊C均值算法(fuzzy c-means算法,简称FCM)。
根据FCM算法得到的隶属矩阵,可以根据一定的标准获得分区成果。阈值Ti的选取带有一定的主观因素,本文试图选择一个经实践检验的合理值[24-25]。因而,Srinivas[24]提出的划分阈值被应用于文中。
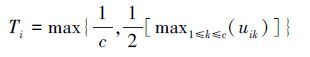
|
本文采用皮尔逊相关系数研究干旱与NDVI植被指数的相关性[1]。研究区域的每个格点有12个NDVI(1—12月)序列,分别计算与1—24月时间尺度SPEI指标的相关系数,这样对于每个格点将有288个相关系数值。自然界植物现象同环境周期性变化,为了剔除物候学对相关性结果的干扰[1],每年1—12月份,1—24月时间尺度这288个相关系数中最大值定义为年最大相关系数Rmax:
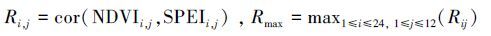
|
式中,cor代表皮尔逊相关函数,i代表时间尺度,1,2,...,24; j代表月份,1,2,…,12。
年最大相关系数对应的时间尺度i定义为Rscale。本研究主要对年最大相关系数和其对应时间尺度的SPEI进行分析。
3 结果与讨论 3.1 模糊聚类分区结果本文聚类分析把693个气象站点1—12月多年平均温度、降雨,以及对应站点位置的多年平均NDVI作为输入气象变量和植被变量。
为了消除不同变量间的方差、数量级、重要程度对聚类分析带来的影响,输入变量首先进行标准化处理,否则那些具有较大方差和数量级的变量将对分区的形成占据控制作用[25]。关于输入聚类分析的变量为什么要进行标准化处理,Dunn 和Hosking[26-27]有更加详细的描述。
| 主成分 | 主成分1 | 主成分2 | 主成分3 | 主成分4 | 主成分5 | 主成分6 | 主成分7 |
| Principle components | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 |
| 累积解释方差/% Cumulative explained variance | 74.50 | 90.60 | 94.70 | 96.50 | 97.90 | 98.70 | 99.20 |
为了避免变量自相关带来的数据冗余,以及维数过高带来的计算灾难,使用主成分分析进行降维[25]。由表 1看出,4个主成分的解释方差已超过95%,因此本文选择前4个主因子外加站点地理信息要素(经度、纬度、高程)作为模糊聚类分析的输入变量。关于分区个数c的确定,本文考虑了6种常用指标[25, 28]优选分区个数:MPC、FPI、NCE、S、SC和XBm[25, 29]。由图 4看出,MPC、FPI、NCE指标在c=7处达到最优,S、SC和XBm指标随着分区个数c的增大,接近单调递减,但是基本在c=7处达到稳定,之后变化幅度较小,因此选择c=7作为最佳分区个数。

|
| 图 4 模糊聚类分析分区个数率定 Fig. 4 The validity of cluster number (horizontal axis represent cluster number c) |
图 5a为初始分区结果,由图 5a可知,初始分区G区重叠模糊的站点较多,根据气候、高程等把C区划分为两个区域。调整后的分区结果如图 5b所示。最终分区结果与肖名忠等人划分的气候分区大致相同[14],也进一步验证了模糊聚类分区的可靠性。

|
| 图 5 模糊聚类分区结果:初始分区结果(a);调整后的最终分区结果(b) Fig. 5 Regionalization by the Fuzzy C-means technique: Initial regionalization results (a); Adjusted regionalization (b) A: 西北区,B: 青藏高原区,C: 内蒙古及黄土高原区,D: 东北区,E: 华北区,F: 长江中下游地区,G: 华南地区,H: 四川云南区 |
SPEI与GIMMS-NDVI(下文简称NDVI)年最大相关系数反映了干旱控制的植被活动的多少,而年最大相关系数对应的干旱时间尺度反映了植被变化对相应时间尺度干旱影响的敏感性[1]。短时间尺度SPEI相关性越大说明植被对干旱影响响应较快;相应的,长时间尺度SPEI相关性越大则说明植被对干旱影响的响应越慢,生态系统对干旱的抵抗力或恢复力稳定性也越强。
由图 6(a)看出,全国大部分区域NDVI与SPEI呈显著正相关,特别是在内蒙古、宁夏、北疆地区、西藏西南部等地,两者相关性较强。而沿北纬30度附近的长江流域区域,特别是长江流域东南部区域,NDVI与SPEI相关性较弱,另外,珠江流域下游、黑龙江东北部、长白山地区、四川西部,NDVI与SPEI也表现出弱的相关性。研究表明,降水量变化为地表植被状况或地表生物量的最佳预测变量[30],但植被变化对干旱事件的响应与干旱时间尺度有显著相关性。由图 6b看出,C区、H区、E区大部分地区、D区西南北、A区北部等区域(区域位置参见图 5b),NDVI对3—6个月SPEI响应敏感。3—6月SPEI干旱变化反映季节尺度土壤水分的变化[9],表明上述区域植被受季节性降水和土壤水分变化影响较大。同时,上述区域也是参照蒸发[31]与蒸发皿蒸发[28]呈增加趋势的区域,以SPEI反映的干旱变化受蒸发影响大,植被变化极易受降水及蒸发变化导致的干旱状况的影响。另外,由图 3可以看出,对3—6个月SPEI响应敏感的区域也是以草地与耕地为主的区域,草地及农作物生长周期的季节性显著,尤其是3个月或半年为时间尺度的时间变化明显,这也是上述区域内植被变化对3—6个月SPEI响应敏感的主要原因。

|
| 图 6 1982—2006年SPEI与NDVI 相关系数空间分布特征 Fig. 6 Spatial patterns of correlation coefficient between SPEI and NDVI during 1982—2006 a: 1—24月时间尺度SPEI与NDVI的年最大相关系数空间分布图(r > 0.40为95%显著性水平上的正相关;r > 0.51为99%显著性水平的正相关);b: 年最大相关系数对应的干旱时间尺度 |
而山东半岛、中国东南沿海、青藏高原以及天山南北坡、东北地区的东北部与北部区域,NDVI对SPEI响应的时间较长,达12—24个月,表明这些区域植被变化对干旱反映的时间尺度较大,在此种现象背后的原因较为复杂。在长江中下游、珠三角,京津冀等地区,由于经济快速发展、城市化程度高,植被覆盖变化在很大程度上受人类活动影响[32-33]。山东半岛及淮河流域,农作物一年两熟,加上农业灌溉的影响,使该区域植被变化与长时间尺度的SPEI相关性较好。而中国东南部大部分地区,主要是由该区域气候湿润,雨量大,降水与蒸发变化不是该区域影响植被变化的主要影响因子,导致NDVI与SPEI相关程度不高。在黑龙江的东北部、长白山地区,其植被主要以针叶林为主,此类植被耐旱、耐寒,一年四季常绿,导致植被对干旱的响应不敏感。四川西部、天山南北坡以及青藏高原地区,植被以林地、高原草甸、草地等为主,该区域以冻土占优势,水壤水分的变化除受降水影响以外,还有降雪以及随气温而变化的冰雪融水及冻土冻融变化的影响,这是导致上述区域NDVI与SPEI相关性较差的主要原因。
3.3 影响植被对干旱响应的因素从图 7看出,影响植被干旱响应的主要因素有多年平均水平衡、降雨、光照时数,上述因子与年最大相关系数的线性相关均通过了99%的显著性检验。如图 7,水平衡综合考虑降雨与潜在蒸散发,对于年最大相关系数,多年平均水平衡解释的变化(R2=0.64)、多年平均降水(R2=0.61)以及多年平均日光照时数(R2=0.58)对植被变化影响的区域差性最大,而多年平均有效积温对植被影响的区域差异性影响较小。其中多年平均水平衡相对单纯的降水变化来讲,是影响NDVI与SPEI相关关系的关键因素。所以,由降水与蒸发之差的水量平衡所反映的土壤水分变化以及土壤持水力,是影响植被变化的关键气象因子,而水量平衡对植被活动的影响在干旱以及半干旱区尤为显著[9]。正是基于这一关系,有研究直接利用植被覆盖程度来研究干旱时空特征演化的检测[34-35]。由图 7看出,从干旱区、半湿润半干旱区、湿润区,多年平均水平衡对植被影响的强度逐减,从定量角度进一步解释了图 6所示现象。光照时数也对年最大相关系数有较强的影响,由7可看出,光照时数对植被变化的影响在青藏高原、新疆、华北以及内蒙等高寒地区最为显著,而在中国东南部地区,光照时数并非是影响植被变化的主要影响因子。有效积温对植被的影响较为复杂,当一个区域有效积温小于3000℃时,有效积温则成为植被变化的关键因素,由图 7可知,青藏高原以及内蒙与华北部分地区,有效积温是决定植被覆盖变化的重要因子。对于有效积温大于3000℃的区域,有效积温则不是影响植被变化的重要因子。
年最大相关系数对应的干旱事件的时间尺度反映了植被对不同时间尺度干旱响应的敏感性。不同植被对特定时间尺度干旱响应的敏感性存在明显差异。相对于多年平均有效积温以及多年平均日光照时数来讲,一个区域与土壤水分及持水力有关的多年平均水平衡以及降水量变化对于该区域植被与干旱事件的时间尺度的相关性较大(图 7)。同时,由图 7可以看出,不同植被类型,植被对干旱事件的响应的快慢也有不同。草地较林地对干旱影响更为敏感,降水量较少区域的植被对干旱的影响更为敏感。多年平均水平衡大于零的区域,植被变化所对应的干旱时间尺度更大,进一步说明一个区域水量平衡变化是决定该区域植被变化的关键因子。一个区域日光照时数与有效积温状况,是影响植被对干旱响应的重要因子,且这种影响关系较为复杂。对于有效积温来讲,当多年有效平均积温低于3000℃时,不同植被类型对于不同时间尺度干旱影响的敏感性差异显著,其中以华北、内蒙、青藏高原地区最为明显,随着多年平均有效积温的增加,林地、灌木、草地对应的干旱时间尺度渐次缩短,表明草地受干旱影响最为敏感,其次为灌木与林地。在多年平均积温大于3000℃的区域,不同植被类型对干旱敏感性的影响差异不大,而多年平均积温越高,植被对干旱的影响越不敏感,尤其是华南地区,各植被类型对干旱的影响最不敏感。由图 7h可以看出,在多年平均日照时数低于6h的区域,如华南及中国东南部地区,各植被类型对干旱时间尺度的变化受多年平均日照时数的影响较小,而多年平均日照时数大于6h的区域,如华北的西北部、内蒙以及西北地区,不同植被类型对不同时间尺度干旱的影响较为明显,其中草地受干旱的影响最为敏感,其次为灌木、林地。

|
| 图 7 多年平均水平衡、多年平均降雨量、多年平均有效积温、多年平均日光照时数对不同土地利用类型、不同区域NDVI与SPEI相关程度的影响;多年平均水平衡、多年平均降雨、多年平均有效积温、多年平均日光照时数对不同土地利用类型、不同区域植被相关系数对应的不同时间尺度干旱的响应 Fig. 7 Illustrate influences of long-term average water balance,long-term mean precipitation,long-term average effective accumulative temperature,land long-term average sunshine hours on vegetation responses to SPEI in different regions with different land use types; demonstrate influences of long-term average water balance,long-term mean precipitation,long-term average effective accumulative temperature,land long-term average sunshine hours on vegetation responses to SPEI with different time scales in different regions with different land use types 标记物的大小代表植被面积[log(面积)],形状代表不同的植被类型,颜色代表不同的分区; 误差线代表±0.5倍标准差,黑色实线表示一元线性拟合,阴影部分表示95%置信区间 |
本文利用模糊聚类分析把中国划分了8个气候分区,对各分区及代表性区域的植被NDVI指标对SPEI干旱指标的响应进行了全面分析,同时探讨了水平衡、降雨、有效积温、日光照时数对不同土地利用类型、不同区域NDVI与SPEI相关程度的影响。上述分析得出如下几点重要的结论:
(1) 我国大部分区域NDVI与SPEI呈显著正相关关系,表明降水及蒸发变化是中国植被覆被变化的关键性因素。NDVI与SPEI相关性显著的区域主要在干旱、半干旱地区,且上述区域NDVI对3—6个月SPEI响应敏感。沿北纬30度附近的长江流域区域(尤其是长江流域东南部地区)、珠江流域下游、黑龙江东北部、长白山地区、四川西部等区域,NDVI与SPEI相关性不显著。
(2) 定量研究影响NDVI与SPEI相关程度的影响因子后发现,影响植被干旱响应的主要因素有水平衡、降雨、光照时数。研究发现水平衡相对单纯的降水变化来讲,是影响NDVI与SPEI相关关系的关键因素,在干旱以及半干旱区尤为显著。光照时数也对NDVI与SPEI相关性有较强影响,光照时数对植被变化的影响在青藏高原、新疆、华北以及内蒙等高寒地区及干旱与半干旱地区最为显著。
(3) 不同区域植被对干旱时间尺度响应的敏感性差异较大,降水与水平衡是影响植被对干旱响应敏感性的主要因素。另外,不同植被类型对不同时间尺度干旱事件的响应也有不同,草地较林地对干旱影响更为敏感,而降水量较少区域的植被对干旱的影响更为敏感。对有效积温的影响来讲,当多年有效平均积温低于3000℃时,不同植被类型对于不同时间尺度干旱影响的敏感性差异显著,草地受干旱影响最为敏感,其次为灌木与林地。对于日照时数来讲,多年平均日照时数低于6h的区域,各植被类型对干旱时间尺度的变化受多年平均日照时数的影响较小,而多年平均日照时数大于6h的区域,草地受干旱的影响最为敏感,其次为灌木、林地。
此项研究系统研究了不同气候区、不同水热条件等对NDVI与SPEI相关性影响,探讨了不同植被类型对不同时间尺度干旱的响应敏感性及可能原因。研究结果将对中国区域植被保育、农业与林业及畜牧业等的规划、管理与发展提供重要理论依据。同时,对理解气候波动和气候变化下植被的脆弱性具有重要意义,可以提高在全球气候变化情景下植被转移预测的精度[1, 7]。
致谢: 文本使用的气象数据来源于中国气象数据共享服务中心,GIMMS NDVI数据来源于国家自然科学基金委员会中国西部环境与生态科学数据中心,土地利用数据来源于欧盟联合研究中心(JRC)空间应用研究所(SAI)开发的GLC2000数据集。| [1] | Vicente-Serrano S M, Gouveia C, Camarero J J, Begueria S, Trigo R, Lopez-Moreno J I, Azorin-Molina C, Pasho E, Lorenzo-Lacruz J, Revuelto J, Moran-Tejeda E, Sanchez-Lorenzo A. Response of vegetation to drought time-scales across global land biomes. Proceedings of the National Academy of Sciences of the United States of America , 2013, 110 (1) : 52–57. DOI:10.1073/pnas.1207068110 |
| [2] | Yu M X, Li Q F, Hayes M J, Svoboda M D, Heim R R. Are droughts becoming more frequent or severe in China based on the Standardized Precipitation Evapotranspiration Index: 1951-2010?. International Journal of Climatology , 2014, 34 (3) : 545–558. DOI:10.1002/joc.3701 |
| [3] | Knapp A K, Smith M D. Variation among biomes in temporal dynamics of aboveground primary production. Science , 2001, 291 (5503) : 481–484. DOI:10.1126/science.291.5503.481 |
| [4] | Zhao M, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science , 2010, 329 (5994) : 940–943. DOI:10.1126/science.1192666 |
| [5] | Ivits E, Horion S, Fensholt R, Cherlet M. Drought footprint on European ecosystems between 1999 and 2010 assessed by remotely sensed vegetation phenology and productivity. Global Change Biology , 2014, 20 (2) : 581–593. DOI:10.1111/gcb.2014.20.issue-2 |
| [6] | Ma Z, Peng C, Zhu Q, Chen H, Yu G, Li W, Zhou X, Wang W, Zhang W. Regional drought-induced reduction in the biomass carbon sink of Canada's boreal forests. Proceedings of the National Academy of Sciences of the United States of America , 2012, 109 (7) : 2423–2427. DOI:10.1073/pnas.1111576109 |
| [7] | Fang J Y, Piao S L, Tang Z Y, Peng C H, Wei J. Interannual variability in net primary production and precipitation. Science , 2001, 293 (5536) : 1723. DOI:10.1126/science.293.5536.1723a |
| [8] | Bevan S L, Los S O, North P R J. Response of vegetation to the 2003 European drought was mitigated by height. Biogeosciences , 2014, 11 (11) : 2897–2908. DOI:10.5194/bg-11-2897-2014 |
| [9] | Ji L, Peters A J. Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sensing of Environment , 2003, 87 (1) : 85–98. DOI:10.1016/S0034-4257(03)00174-3 |
| [10] | Engelbrecht B M J, Comita L S, Condit R, Kursar T A, Tyree M T, Turner B L, Hubbell S P. Drought sensitivity shapes species distribution patterns in tropical forests. Nature , 2007, 447 (7140) : 80–82. DOI:10.1038/nature05747 |
| [11] | 于泉洲, 刘煜杰, 梁春玲, 周蕾, 张祖陆, 王永生. 基于遥感的南四湖湿地干旱响应特征研究. 资源科学 , 2014, 36 (7) : 1519–1526. |
| [12] | 叶春, 吴桂平, 赵晓松, 王晓龙, 刘元波. 鄱阳湖国家级自然保护区湿地植被的干旱响应及影响因素. 湖泊科学 , 2014, 26 (2) : 253–259. |
| [13] | 牟成香, 孙庚, 罗鹏, 王志远, 罗光荣. 青藏高原高寒草甸植物开花物候对极端干旱的响应. 应用与环境生物学报 , 2013, 19 (2) : 272–279. |
| [14] | Xiao M Z, Zhang Q, Singh V P, Chen X H. Regionalization-based spatiotemporal variations of precipitation regimes across China. Theoretical and Applied Climatology , 2013, 114 (1/2) : 203–212. |
| [15] | Zhang Q, Qi T Y, Singh V P, Chen Y D, Xiao M Z. Regional frequency analysis of droughts in China: a multivariate perspective. Water Resources Management , 2015, 29 (6) : 1767–1787. DOI:10.1007/s11269-014-0910-x |
| [16] | 宋怡, 马国明. 基于GIMMS AVHRR NDVI数据的中国寒旱区植被动态及其与气候因子的关系. 遥感学报 , 2008, 12 (3) : 499–505. |
| [17] | 徐文婷, 吴炳方, 颜长珍, 黄慧萍. 用SPOT-VGT数据制作中国2000年度土地覆盖数据. 遥感学报 , 2005, 9 (2) : 204–214. |
| [18] | Bartholomé E, Belward A S. GLC2000: a new approach to global land cover mapping from Earth observation data. International Journal of Remote Sensing , 2005, 26 (9) : 1959–1977. DOI:10.1080/01431160412331291297 |
| [19] | Vicente-Serrano S M, Beguería S, López-Moreno J I. A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. Journal of Climate , 2010, 23 (7) : 1696–1718. DOI:10.1175/2009JCLI2909.1 |
| [20] | 王林, 陈文. 标准化降水蒸散指数在中国干旱监测的适用性分析. 高原气象 , 2014, 33 (2) : 423–431. |
| [21] | Ruspini E H. A new approach to clustering. Information and Control , 1969, 15 (1) : 22–32. DOI:10.1016/S0019-9958(69)90591-9 |
| [22] | Dunn J C. A fuzzy relative of the ISODATA process and its use in detecting compact, well-separated clusters. Journal of Cybernetics , 1974, 3 (3) : 32–57. |
| [23] | Bezdek J C. A convergence theorem for the fuzzy ISODATA clustering algorithm. IEEE Transactions on Pattern Analysis and Machine Intelligence , 1980, 1 (2) : 1–8. |
| [24] | Srinivas V V, Tripathi S, Ramachandra Rao A, Govindaraju R S. Regional flood frequency analysis by combined self-organizing feature map and fuzzy clustering. Journal of Hydrology , 2008, 348 (1/2) : 148–166. |
| [25] | Satyanarayana P, Srinivas V V. Regionalization of precipitation in data sparse areas using large scale atmospheric variables-A fuzzy clustering approach. Journal of Hydrology , 2011, 405 (3/4) : 462–473. |
| [26] | Dunn D M, Landwehr J M. Analyzing clustering effects across time. Journal of the American Statistical Association , 1980, 75 (369) : 8–15. DOI:10.1080/01621459.1980.10477414 |
| [27] | Hosking J R M, Wallis J R. Regional Frequency Analysis: An Approach Based on L-moments. Cambridge: Cambridge University Press, 2005. |
| [28] | Zhang Q, Qi T Y, Li J F, Singh V P, Wang Z Z. Spatiotemporal variations of pan evaporation in China during 1960-2005: changing patterns and causes. International Journal of Climatology , 2014, 35 (6) : 903–912. |
| [29] | Dave R N. Validating fuzzy partitions obtained through c-shells clustering. Pattern Recognition Letters , 1996, 17 (6) : 613–623. DOI:10.1016/0167-8655(96)00026-8 |
| [30] | Wang J, Price K P, Rich P M. Spatial patterns of NDVI in response to precipitation and temperature in the central Great Plains. International Journal of Remote Sensing , 2001, 22 (18) : 3827–3844. DOI:10.1080/01431160010007033 |
| [31] | Zhang Q, Xu C Y, Chen X H. Reference evapotranspiration changes in China: natural processes or human influences?. Theoretical and Applied Climatology , 2011, 103 (3/4) : 479–488. |
| [32] | 朴世龙, 方精云. 1982-1999年我国陆地植被活动对气候变化响应的季节差异. 地理学报 , 2003, 58 (1) : 119–125. |
| [33] | 张学珍, 戴君虎, 葛全胜. 1982-2006年中国东部春季植被变化的区域差异. 地理学报 , 2012, 67 (1) : 53–61. |
| [34] | Garik Gutman G. Towards monitoring droughts from space. Journal of Climate , 1990, 3 (2) : 282–295. DOI:10.1175/1520-0442(1990)003<0282:TMDFS>2.0.CO;2 |
| [35] | Tucker C J. Comparing SMMR and AVHRR data for drought monitoring. International Journal of Remote Sensing , 1989, 10 (10) : 1663–1672. DOI:10.1080/01431168908903997 |